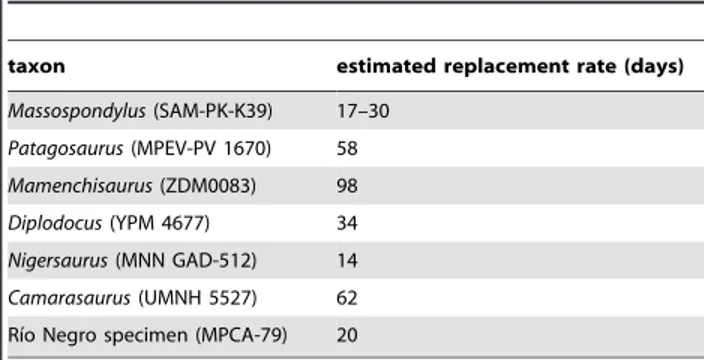
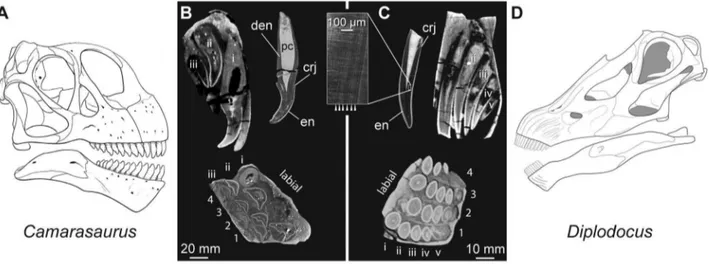
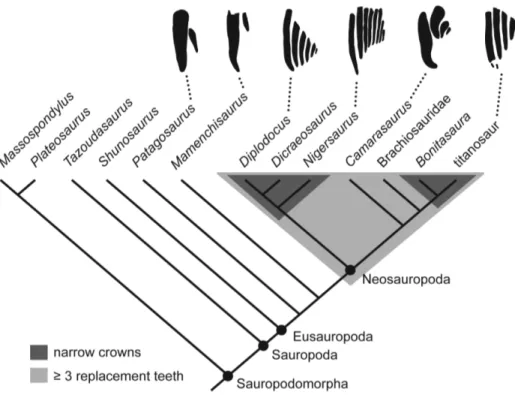
Evolution of high tooth replacement rates in sauropod dinosaurs.
Texto
Imagem
Documentos relacionados
could be related to different mutations located in several genes involved in tooth enamel formation, and that the amelogenin gene forms the skeleton for the mineralization and
Rare/ Painless slow-growing lesion which could present bone expansion, and, in some cases, be associated with an unerupted tooth (involvement with the tooth crown)/
Given the aforementioned issues, the present study aimed to estimate the prevalence of self-reported edentulism, severe tooth loss and lack of functional dentition in
A: Sauropod tooth from the Matsuo Group (the Toba sauropod tooth); B and C: Sauropod teeth from the Futaba Group (the Kohisa specimens), IMCF no.. 959 and 1122, respectively; D and
Methodology/Principal Findings: We investigated the effects of a major environmental stressor (a decrease in water volume) on diversity-abundance and diversity-stability relations
2,4,5 Clinical findings (increased volume of the face, presence of an intra-oral mass and tooth mobility) and the radiology (poorly defined borders of the lesion) of this case
An unusual replacement pattern occurs within acrodontan lizards (agamids and chameleons), in which tooth replacement occurs during embryonic developmental stages, but ceases
Enamel findings in AI are highly variable, ranging from deficient enamel formation to defects in the mineral and protein content.. Enamel formation requires the expression of
