Ana Maria Araújo Soares
Implementações metodológicas para o estudo
eletrofisiológico e comportamental em um modelo
animal de autismo
Ana Maria Araújo Soares
Implementações metodológicas para o estudo
eletrofisiológico e comportamental em um modelo
animal de autismo
Dissertação apresentada ao Instituto do Cérebro da Universidade Federal do Rio Grande do Norte, para a obtenção de Tí-tulo de Mestre em Neurociências.
Orientador: Rodrigo Neves Romcy Pereira
ii
Soares, Ana M. A.
Implementações metodológicas para o estudo eletrofisi-ológico e comportamental em um modelo animal de autismo
57 páginas
Dissertação (Mestrado) - Instituto do Cérebro da Uni-versidade Federal do Rio Grande do Norte.
1. eletrofisiologia in vivo
2. autismo
3. modelo VPA
I. Universidade Federal do Rio Grande do Norte. Instituto do Cérebro.
Comissão Julgadora:
Prof. Dr. Prof. Dr.
Claudio Queiroz Marcelo Cairrão Araújo Rodrigues
Prof. Dr.
iii
iv
Agradecimentos
Ao sair de engenharia elétrica para encarar um mestrado em neurociências, nunca imaginei
que chegar a esse momento seria fácil. Mas confesso também que não sabia quão grande seria
esse desafio. Durante essa jornada tive a enorme sorte de cruzar pessoas maravilhosas em meu
caminho e, sem elas, com certeza eu não estaria escrevendo essas palavras hoje.
Depois de três meses conhecendo o mínimo da neurociência, me habituando a manipular os
ratos, aplicar injeções e outras coisinhas mais. Chegou o dia que eu realizaria a minha primeira
cirurgia em um animal. Naquele dia eu estava tão nervosa, a vontade que eu tinha era de largar
tudo naquele momento e voltar para os meus circuitos eletrônicos. Foram 9h de cirurgia. Durante
todo esse tempo, não fiquei sozinha um minuto. Houve um revesamento de colegas naquela sala
para ajudar, me aconselhar, me acalmar ou, simplesmente, trazer algo para eu me alimentar.
No final do dia, eu estava morta, o rato vivo. E comecei a pensar em todas as pessoas que me
ajudaram durante aquele dia, em como eu nunca teria conseguido sem cada uma delas. Fiz uma
listinha e fui comprar chocolates para cada uma delas que tinha me ajudado. Resultado: 10% da
bolsa daquele mês foi gasto em chocolates. Essas 9h são um retrato do período em que trabalhei
no Instituto, muitos desafios, mas sempre vencidos com ajuda de inúmeras mãos. Então, hoje,
essa listinha cresceu muito e não seria justo que eu citasse cada pessoa que faz parte dela, pois
a chance de esquecer alguém seria muito grande. Então meu agradecimento é amplo e irrestrito
a todos do ICe que, com certeza, tem uma pequena responsabilidade por essa dissertação. Seja
por me ajudar nas cirurgias, seja por ensinar o que é um neurônio ou simplesmente por ter tido
tempo para um cafézinho naqueles dias que eu estava precisando.
Em particular, gostaria de agradecer ao meu orientador Prof. Rodrigo Neves Romcy Pereira e
aos professores Richardson Leão, Katharina Leão e Adriano Tort que participaram do meu comitê
de acompanhamento e me deram sugestões e contribuições muito relevantes para realização desse
trabalho. Além disso, agradeço a Capes que apoiou financeiramente a realização desse mestrado.
Por fim, agradeço a toda minha família. Meus pais que sempre foram meus exemplos e
tornaram viável todo caminho que percorri para chegar aqui. Meus irmãos que me aturaram
sempre, me ligavam toda semana para me dar força e não me deixaram desistir. E ao meu
com-panheiro, melhor amigo, namorido, Vincent, que sempre me apoiou e me incentivou a continuar
nos momentos mais difíceis.
v
Resumo
O autismo é um transtorno do desenvolvimento que se manifesta nos primeiros anos de
vida e apresenta semiologia heterogênea. Esta patologia afeta a maturação do encéfalo e produz
alterações sensoriais, de linguagem e de interação social no início na infância. O modelo
expe-rimental de autismo utilizando ácido valproico (VPA) durante o período gestacional tem sido
demonstrado ter alta validade de face e permitir estudos tanto das bases neuropatológicas quanto
neuro-funcionais durante o desenvolvimento. A despeito do recente interesse por este modelo
como instrumento de compreensão dos aspectos básicos da fisiopatologia do autismo, a maioria
dos estudos experimentais têm se concentrado nos aspectos comportamentais, histológicos e
ce-lulares. Neste trabalho, foram propostas estratégias experimentais de avaliação comportamental
associadas a eletrofisiologiain vivo, uma técnica que nunca fora utilizada para avaliação desse
modelo. Animais controles e experimentais, submetidos previamente a um procedimento cirúrgico
para implante de eletrodos crônicos, participaram de experimentos de livre exploração, interação
social e condicionamento ao medo.
vi
Abstract
Autism is a neurodevelopmental disorder that presents a very heterogeneous semiology. This
pathology affects brain maturation causing sensory, social and language impairments in early
childhood. The animal model of autism induced by prenatal exposure to Valproate (VPA)
exhibits both construct and face validity. It allows to study autism neuropathological and
neurofunctional basis during development. Several works based on VPA models that aim to
understand behavioral, histologic and cellular aspects of autism have been reported. In this work,
we present new experiments to evaluate electrophysiological activity in vivo during behavioral
tasks of model VPA. Control and experimental animals were submitted to a surgical implantation
of chronical electrode arrays on medial Prefrontal Cortex (mPFC) and hippocampus. After one
week, these animals were used in open-field, social interaction and fear conditioning experiments.
Lista de Figuras
1.1 Etiologia do autismo . . . 2
1.2 Diagnóstico segundo DSM-V . . . 2
1.3 Curva de crescimento cerebral . . . 4
1.4 Animais VPA x Animais controles . . . 8
4.1 Linha temporal do desenho experimental. . . 16
4.2 Matrizes de eletrodos. . . 18
4.3 posição dos eletrodos . . . 19
4.4 Experimento de interação social. . . 21
4.5 Gaiola compartilhada com animais familiares . . . 22
4.6 Setup do experimento de interação social . . . 22
4.7 Representação temporal dos estímulos sonoros. . . 24
4.8 Divisão do aparato em zonas . . . 26
5.1 Trecho ilustrativo de LFP dos animais em exploração. . . 28
5.2 PSD do LFP dos animais em atividade exploratória . . . 28
5.3 Trecho ilustrativo de LFP dos animais em repouso . . . 29
5.4 PSD do LFP dos animais em repouso . . . 29
5.5 Trecho ilustrativo de LFP dos animais em repouso . . . 30
5.6 PSD do LFP dos animais em interação social . . . 31
5.7 PSD média do ritmo teta . . . 31
5.8 PSD média do ritmo beta . . . 32
5.9 PSD média do ritmo low gamma . . . 32
5.10 PSD média do ritmo high gamma. . . 33
Lista de Figuras viii
5.12 Taxa de congelamento durante extinção do medo . . . 35
5.13 Espectrograma cortical do animal controle na extinção do medo. . . 36
5.14 Espectrograma hipocampal do animal controle na extinção do medo . . . 37
5.15 Espectrograma cortical do animal VPA na extinção do medo . . . 38
Lista de abreviaturas
CEUA Comissão de Ética no Uso de Animais em Pesquisa
CPFm Cortex pré-frontal medial
CS+ estímulo sonoro pareado ao choque
CS- estímulo sonoro não pareado ao choque
DMN Default Mode Network
DSM-V Manual de Diagnóstico e Estatística dos Transtornos Mentais V
E12 Dia embrionário 12
FFT Transformada Rápida de Fourier
i.p. intraperitoneal
ICe Instituto do Cérebro
IRM Imagem por Ressonância Magnética
PSD Densidade Espectral de Potência
TDI Transtorno Desintegrativo Infantil
TEA Transtornos do Espectro Autista
UFRN Universidade Federal do Rio Grande do Norte
VPA Ácido valpróico
Sumário
1 Introdução 1
1.1 Autismo . . . 1
1.2 Neuropatologia do autismo . . . 4
1.3 Alterações funcionais e de circuitaria no autismo . . . 5
1.4 Modelo de autismo por exposição pré-natal a ácido valpróico . . . 6
1.5 Circuitos neurais em modelos experimentais de autismo . . . 9
1.6 Comunicação hipocampo-córtex pré-frontal medial . . . 10
1.7 Medo e extinção do medo . . . 11
1.8 Vocalizações ultrassônicas de ratos . . . 11
2 Justificativa 14 3 Objetivos 15 3.1 Objetivos específicos . . . 15
4 Materiais de Métodos 16 4.1 Modelo animal de autismo . . . 16
4.2 Confecção de matrizes de eletrodos . . . 17
4.3 Cirurgia para implante de eletrodos. . . 18
4.4 Acoplamento hipocampo-CPFm durante atividade exploratória e interação social 20 4.5 Condicionamento pelo medo . . . 23
4.6 Extinção do medo . . . 24
4.7 Registro eletrofisiológico . . . 24
4.8 Análise de dados . . . 25
4.8.1 Pré-tratamento do sinal . . . 25
Sumário xi
5 Resultados 27
5.1 Exploração de um novo ambiente . . . 27
5.2 Animais em interação social . . . 30
5.3 Condicionamento ao medo . . . 34
6 Discussão 40
Capítulo 1
Introdução
1.1
Autismo
O autismo é um transtorno do desenvolvimento de etiologia multifatorial que se manifesta nos
primeiros anos de vida apresentando diversas formas de expressão. A descrição inicial dessa
patologia foi realizada pelo psicólogo americano Leo Kanner, em 1943, que descreveu um grupo
de 11 crianças com dificuldade de interação social, deficiência na comunicação verbal e
não-verbal, ansiedade, dificuldades em processar informações emocionais e comportamentos repetitivos
(Kanner et al., 1943). Segundo Kanner, o autismo afetava crianças biologicamente normais
possivelmente em decorrência de cuidados neonatais inadequados. Esta hipótese prevaleceu até
a década de 60, quando diversos pesquisadores passaram a defender causas biológicas para o
transtorno. (figura1.1).
Atualmente, a denominação transtorno do espectro autista (TEA) engloba várias formas
de expressão do autismo: o autismo propriamente dito, a síndrome de Asperger, o transtorno
desintegrativo infantil (TDI) e TDI não-especificado. Esta classificação foi apresentada na última
versão do Manual de Diagnóstico e Estatística dos Transtornos Mentais (DSM-V) (Association
et al.,2003). Vale ressaltar que cada uma destas patologias difere quanto às suas características
clínicas, etiopatogênicas e de prognóstico. A síndrome de Rett, anteriormente presente como
um TEA por suas características clínicas, foi removida deste grupo por apresentar etiologia
genética conhecida. Segundo as novas definições, para o diagnóstico de TEA são necessárias
disfunções em pelo menos dois domínios de comportamentos: interação/comunicação social
e padrões comportamentais repetitivos e restritivos (figura 1.2). Também é muito comum a
1.1. Autismo 2
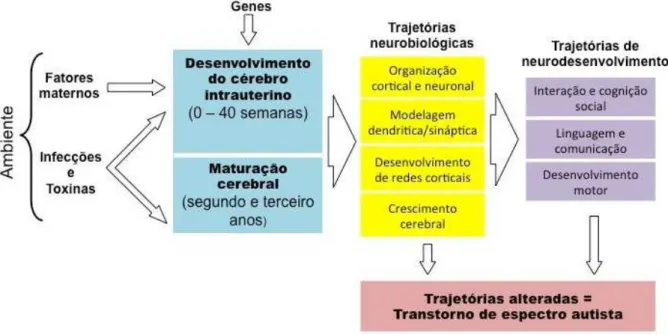
Figura 1.1: Modificação na expressão gênica ou exposição a fatores ambientais durante o período de neurogênese e maturação das redes neurais promovem alterações na organização sináptica e cortical que refletem no desenvolvimento motor e comportamental característico do indivíduo com autismo (Adaptado dePardo e Eberhart(2007)).
Declínio cognitivo pode ser observado em uma parcela dos autistas, nos quais atividade paroxística
ou crises epilépticas são mais frequentes (García-Peijas,2009).
Figura 1.2: Para diagnóstico de TEA, segundo DSM-V, é necessária disfunção em dois domínios comportamentais.
A implementação de novos critérios de diagnóstico e serviços especializados mostram que
a prevalência atual dos TEA na população atinge 1 em 68 crianças norte-americanas, sendo a
prevalência do autismo propriamente dito de 1 em 500 (for Disease Control et al.,2014). Meninos
são mais afetados pelo transtorno, na proporção de 4:1 com relação a meninas (Wing e Potter,
2002;Geschwind,2009).
Os principais fatores etiológicos associados ao autismo podem ser agrupados em fatores
1.1. Autismo 3
embrionário e/ou pós-natal levando ao fenótipo autista. Com relação aos fatores genéticos, vários
estudos mostram um aumento no risco para TEA em famílias com um ou mais parentes com
o transtorno. Além disso, a concordância na expressão do autismo entre gêmeos monozigóticos
varia de 60-90 %, enquanto que entre irmãos dizigóticos é de 3-5 % (Gilbert, 2000). Contudo, este
componente genético parece depender da interação entre genes de risco e a composição genética
global do indivíduo (background genético). De fato, a maioria dos estudos são correlacionais e
ainda não foi encontrada nenhuma alteração gênica que por si só fosse responsável por
desenca-dear TEA em 100% dos portadores Ű o que indica que a herança de traços autistas deve ser a
resultante da interação entre múltiplos genes.
Outro fator etiológico associado ao autismo é a resposta imune da mãe quando exposta a
agentes patogênicos durante a gravidez. Evidências indicam que estas reações podem afetar o
desenvolvimento cerebral do feto através da placenta, levando a anomalias no desenvolvimento
neuronal ou glial (Mead e Ashwood,2015;Knuesel et al.,2014). Um exemplo disto são os estudos
epidemiológicos mostrando que infecção materna por rubéola durante a gravidez aumenta o
risco de desenvolvimento de autismo nos filhos (Singh,2009). De maneira similar, a exposição
do bebê a agentes infecciosos no período pós-natal também pode contribuir para alterações
comportamentais semelhantes às do autismo, como infecções do sistema nervoso central como
meningoencefalites bacterianas e virais (García-Peijas,2009).
Recentemente, o aumento do uso de pesticidas e drogas para controle de algumas patologias
(p.ex. câncer e epilepsia) têm aumentado o risco de mulheres grávidas darem a luz a crianças
autistas. Drogas administradas durante a gravidez como a talidomida, ácido valpróico (VPA) e
etanol, principalmente no primeiro trimestre de gestação, são agentes teratogênicos associados ao
risco de desenvolvimento de alterações comportamentais e autismo (Arndt et al.,2005;Strömland
et al.,1994; Christianson et al., 1994; Nanson,1992). Em um recente estudo epidemiológico
avaliando uma população de mais de 600 mil crianças nascidas com autismo na Dinamarca
entre 1996 e 2006 verificou-se que o uso do VPA, por mulheres com ou sem epilepsia, durante
a gravidez aumenta o risco de nascimento de crianças com TEA ou autismo propriamente dito
em até 5 vezes (Christensen et al., 2013). Interessante, os autores não observaram diferenças
na frequência de crianças com TEA ou autismo propriamente dito com relação ao período
da gravidez (1o ou 2o-3o trimestres) em que as mães receberam VPA ( 750 mg/dia). O uso
de outras drogas anti-epilépticas, como carbamazepina (>500 mg/dia), oxcarbazepina (>500
1.2. Neuropatologia do autismo 4
TEA ou autismo propriamente dito.
1.2
Neuropatologia do autismo
Estudos de imagem por ressonância magnética (IRM) e de tecidopost-mortem revelam a
exis-tência de padrões morfológicos alterados bem característicos de pacientes autistas quando
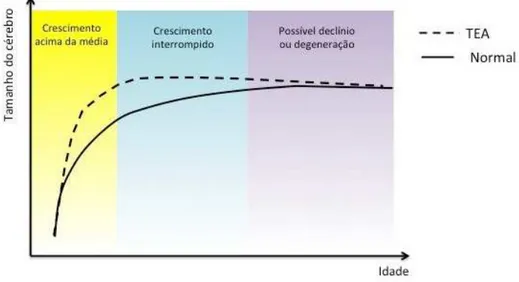
com-parados a indivíduos controle. Em termos gerais,Courchesne et al.(2007) observaram que os
encéfalos de pacientes autistas sofrem um rápido crescimento durante o período pós-natal inicial
(0-2 anos) e depois passam por uma desaceleração no crescimento com a idade (4 anos em diante)
(figura 1.3). Em alguns casos, uma terceira fase de mudanças estruturais pode ser observada em
regiões cerebrais de pré-adolescentes, caracterizada por um processo de degeneração neuronal.
Figura 1.3: Curva de crescimento do cérebro ao longo dos anos em indivíduos autistas e normais. (Courchesne et al.,2001)
Entre as principais alterações neuropatológicas estão o aumento do volume cerebral durante
os primeiros anos de vida (figura1.3), anormalidades nos núcleos dos nervos cranianos, hipoplasia
de estruturas do tronco cerebral, perda de células de Purkinje no cerebelo, além de alterações
nas estruturas do córtex frontal e lobo temporal (Stevenson et al.,1997;Courchesne et al.,2001;
Bauman e Kemper, 2005). A consistência destas alterações estruturais foi avaliada em uma
série de 17 estudos com 359 pacientes com TEA mostrando aumento do volume encefálico e
do volume da amígdala em crianças, redução do volume do corpo caloso em jovens e adultos,
redução de matéria branca no lobo temporal, aumento da espessura cortical no lobo parietal,
1.3. Alterações funcionais e de circuitaria no autismo 5
córtex pré-frontal e giro cingulado (Chen et al.,2011). Regionalmente, os lobos frontais e os lobos
occipitais apresentam o maior e o menor grau de crescimento observados (Carper et al.,2002),
respectivamente , sendo que dentro do lobo frontal, a convexidade dorsolateral se mostra mais
hiperatrofiada (Carper e Courchesne,2005). Além disso, pacientes autistas apresentam aumento
da densidade celular e diminuição do tamanho dos neurônios no córtex pré-frontal (CPFm) e na
área lobo temporal. Essas estruturas estão fortemente envolvidas no processamento cognitivo,
social e emocional (Bauman e Kemper, 2005). Schumann et al. (2004) e Groen et al. (2010),
também mostraram um aumento do volume do hipocampo esquerdo em crianças e adolescentes
autistas. A análise ultraestrutural de tecidos post-mortem de pacientes com TEA, utilizando
a coloração de Golgi, revelou um aumento na densidade de espinhas dendríticas apicais em
neurônios piramidais da camada II dos lobos frontal, temporal e parietal, assim como na camada
V restrita ao lobo temporal (Hutsler e Zhang,2010). Anormalidades sinápticas e redução do
tamanho das minicolunas corticais também foram observadas porCasanova et al.(2002). Estas
observações fortemente sugerem um funcionamento aberrante em diversas circuitarias corticais
em indivíduos com TEA.
1.3
Alterações funcionais e de circuitaria no autismo
Do ponto de vista funcional, estudos recentes indicam uma sincronização inter-hemisférica
re-duzida em crianças autistas que se correlaciona com a severidade do autismo (Anderson et al.,
2011; Dinstein et al., 2011). Associado aos estudos que indicam alterações de mielinização, é
plausível supor que o comprometimento da conectividade entre regiões encefálicas distantes
afetam a integração de informações multimodais. Evidências recentes também apontam para
disfunções em circuitos locais. A combinação entre dificuldade de comunicação social e rituais
motores com estados de hiper-sensibilidade sensorial, emocional e seletividade atencional alterada
apresentadas pelos autistas podem ser o resultado do encurtamento e desorganização de certas
projeções. Alguns autores chegam a propor que no cérebro autista os sinais são
insuficiente-mente diferenciados do ruído ou da informação irrelevante e, assim o conteúdo de informação
relevante está reduzido (Baron-Cohen e Belmonte,2005). Algumas evidências neuropatológicas
e funcionais apontam para uma combinação de hiper-conectividade em circuitos locais com
hipo-conectividade de longa distância (Belmonte et al., 2004a,b). Esta idéia é consistente não
1.4. Modelo de autismo por exposição pré-natal a ácido valpróico 6
auditiva, percepção visual, impulsividade e empatia (Teitelbaum et al.,1998;Plaisted et al.,2003;
Milne et al.,2002; Just et al.,2004). Esta hipótese poderia também explicar tanto as ilhas de
super-habilidades e interesses restritos, como a dificuldade dos autistas em integrar elementos
perceptuais em uma unidade.
A rede cerebral denominada Default Mode Network (DMN) tem extrema importância na
cognição social e é formada pelo córtex cingulado posterior, córtex pré-frontal medial, córtex
temporal lateral, lobo parietal inferior e hipocampo (Buckner et al.,2008). Achados consistentes
mostram que pacientes autistas apresentam déficits na comunicação entre os nós dessa rede
(Assaf et al.,2010;Weng et al.,2010). Uma hipoconectividade funcional na DMN seria coerente
com os relatos comportamentais de pacientes autistas com deficiências em teoria da mente,
empatia e interação social. Além da DMN, diversas outras redes implicadas em processamento
de informações sociais e emocionais parecem alteradas nos pacientes autistas (Uddin et al.,2013;
von dem Hagen et al.,2012; Supekar et al., 2013;Keown et al.,2013). Isto poderia explicar a
dificuldade de integração de informações sócio-emocionais apresentada por estes indivíduos.
1.4
Modelo de autismo por exposição pré-natal a ácido valpróico
Diversos modelos experimentais têm sido propostos para o estudo do autismo, sendo os modelos
em roedores os mais comuns. Como em qualquer modelo de doenças do sistema nervoso é
importante ter claro suas principais características como validade de face, validade de construto e
validade preditiva. Como validade de face se entende que o modelo deve apresentar características
comportamentais e fisiológicas (moleculares, celulares e eletrofisiológicas) mais próximas possível
do transtorno observado em humanos. Validade de construto se refere aos mecanismos celulares e
moleculares por trás da etiologia da doença. Já a validade preditiva trás a informação do quanto
uma estratégia de tratamento aplicada no modelo animal é possível de reproduzir seus efeitos
em humanos. Assim, os modelos em roedores apresentam vários graus de validade, sendo que a
presença de comportamentos sociais alterados e estereotipados - os endofenótipos mais comuns
do transtorno em humanos - o que garante em diferentes graus sua validade de face.
Em roedores, temos (1) os modelos baseados em alterações genéticas específicas envolvendo
neurexina-1, neuroligina-3, neuroligina-4, reelina, MeCP2, TSC2 e FMR1 (Etherton et al.,2009;
Patterson,2011;Waltereit et al.,2011); (2) os modelos de linhagens geneticamente selecionadas
1.4. Modelo de autismo por exposição pré-natal a ácido valpróico 7
T+tf/J (BTBR) (Scattoni et al.,2008), (3) os modelos baseados em lesões de estruturas límbicas
como a amígdala e o hipocampo ventral (Daenen et al.,2002; Wolterink et al.,2001) e (4) os
modelos baseados em exposição a drogas ou agentes patogênicos durante a gestação (Rodier
et al.,1997;Dufour-Rainfray et al.,2010).
O modelo que decidimos estudar é induzido por exposição de ratos ao ácido valpróico (VPA)
no período embrionário. Este é um modelo que agrega tanto uma consistente validade de face
quanto validade de construto. Uma vez que produz alterações comportamentais semelhantes
àquelas observadas em crianças nascidas de mães que receberam VPA durante a gravidez, uma
droga usada para estabilização de humor e anti-epiléptica (Rodier et al.,1997;Williams et al.,
2001). O trabalho que trouxe a tona a correlação entre uso de VPA durante a gestação e o autismo
foi o de Christianson et al. (1994). Estes autores observaram que crianças que apresentavam
síndrome do valproato fetal (caracterizada por traços fenotípicos de má formação) recebiam com
maior frequência diagnóstico de TEA. O período de sensibilidade à droga e o surgimento de
má-formações levaram à hipótese de que o TEA pudesse ser o resultado de alguma interferência
com os processos de desenvolvimento durante o período embrionário. De fato, além do VPA,
a exposição do embrião a outros agentes patogênicos durante o primeiro trimestre de gestação
parece também estar relacionado a um maior número de casos de autismo.
Em 1996, Rodier et al. (1996) propuseram um modelo animal para estudo das alterações
de espectro autista baseado na administração pré-natal de VPA em ratas prenhasRodier et al.
(1996). Desde então, este modelo tem sido reproduzido e recentemente foi caracterizado com
sucesso por nosso grupo de pesquisa. Em resumo, ratas prenhas, com controle do dia da
fecunda-ção, recebem uma injeção de VPA, em dose única na data de fechamento do tubo neural de sua
prole (E12). Como resultado, os filhotes apresentam uma série de alterações comportamentais
que se assemelham em muito àquelas observadas em pacientes autistas. Dentre estas alterações
se observa um retardo no desenvolvimento pós-natal, maior atividade locomotora, menor
intera-ção social, aumento do comportamento esterotipado (processos de auto-limpeza), aumento de
ansiedade e baixa sensibilidade à dor. Ao contrário de drogas como a talidomida, que apresentam
efeitos diferentes quando administradas em ratos e em humanos, o VPA tem mostrado resultados
semelhante aos observados na espécie humana, sendo por isso um modelo bastante validado na
literatura para o estudo das bases neurofisiológicas do autismo (Schneider e Przewşocki,2005;
Markram et al.,2007;Bambini Júnior,2010;Sousa,2013).
1.4. Modelo de autismo por exposição pré-natal a ácido valpróico 8
(a) (b) (c)
(d) (e)
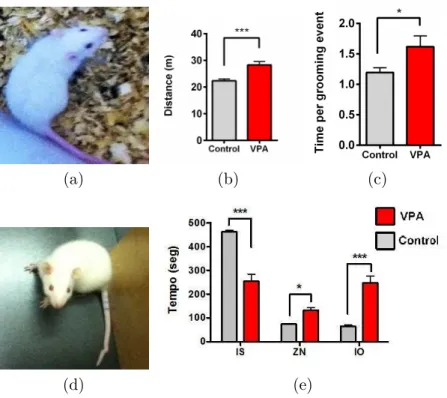
Figura 1.4: Diferenças entre animais controle e animais VPA. (a) Fotografia de um animal controle; (b) Distância média percorrida por animais de cada grupo em 5 minutos de livre exploração;(c) Duração de episódios de auto-limpeza;(d) Fotografia de um animal VPA onde nota-se má formação da cauda;(e)Tempo médio gasto por animal dos dois grupos em cada uma das 3 zonas: interação social (IS), zona neutra (ZN) e interação com objeto (IO).Sousa(2013)
elas, podemos citar a redução da capacidade de proteção contra estresse oxidativo (
Bambini-Junior et al., 2011), alteração na neurotransmissão NMDA (Rinaldi et al., 2007), aumento da
recaptação de glutamato (Silvestrin et al.,2013) e hipo-responsividade dopaminérgica (Hara
et al.,2015). Recentemente nosso grupo mostrou que além destas alterações, os animais expostos
ao VPA em útero apresentam alterações na circuitaria inibitória do córtex pré-frontal medial
(CPFm). Esta alteração se deve a uma redução, nas áreas do córtex cingulado anterior (Cg) e
pré-límbica (PL) do CPFm, do número de neurônios inibitórios que expressam parvalbumina
(PV+). Estas células constituem uma população de neurônios em cesto ou candelabro que têm
alta taxa de disparos e forte poder inibitório, pois inervam o corpo celular e o segmento inicial
do axônio de seus alvos, respectivamente. O que sugere uma hiper-excitabilidade local no CPFm
e uma possível disfunção na conectividade entre o hipocampo e o CPFm neste modelo (Brandão
e Romcy-Pereira,2015). Além disto,Sousa(2013) também observou alterações comportamentais
e na circuitaria inibitória do CPFm na geração de filhotes de pais tratados com VPA.
apresen-1.5. Circuitos neurais em modelos experimentais de autismo 9
tam alterações nas respostas eletrofisiológicas do córtex auditivo primário (A1) a tons puros.
Estes animais apresentam maior representação de frequências altas (10-50 kHz) em A1, assim
como limiar de intensidade sonora mais alto e campos receptivos auditivos menos específicos.
Estas características indicam um processamento auditivo aberrante nos animais VPA quando
comparados aos controles.
1.5
Circuitos neurais em modelos experimentais de autismo
Estudos recentes mostram que animais expostos ao VPA na fase embrionária apresentam
alte-rações em microcircuitos no CPFm, na forma de uma hiper-conectividade e hiper-plasticidade
na circuitaria cortical local quando comparada aos animais controle (Rinaldi et al.,2008). Estes
achados sugerem que tal padrão estrutural e funcional seria a representação no modelo animal do
que ocorre no córtex de indivíduos autistas. Esta hiperfuncionalidade local seria a consequência
de distúrbios na formação e maturação destas circuitarias (Markram et al.,2007). Anormalidades
na migração neuronal e/ou na maturação sináptica poderiam levar a uma organização aberrante
das conexões locais e das projeções de longa distância, decorrentes da exposição ao VPA durante
o desenvolvimento embrionário.
Estados comportamentais que indicam um certo grau de hiperexcitabilidade neuronal em
pa-cientes autistas, como ansiedade elevada, epilepsia, distúrbios do sono ou agressividade sugerem
um desbalanço excitatório-inibitório em circuitos neurais autistas (Gogolla et al.,2009;
Rubens-tein e Merzenich,2003). As causas deste desequilíbrio podem ser o aumento da neurotransmissão
glutamatérgica, a diminuição da neurotransmissão GABAérgica, ou mesmo a combinação dos
dois fatores. Estudos eletrofisiológicos em fatias cerebrais de animais VPA reforçam esta hipótese.
Rinaldi e colaboradores mostraram um aumento da neurotransmissão mediada por receptores
NMDA em fatias de animais VPA quando comparadas a fatias dos controles (Rinaldi et al.,2007).
Em outro estudo realizado porBanerjee et al.(2013), onde se caracterizou o sistema
GABAér-gico, os autores mostram um forte prejuízo da neurotransmissão inibitória pré e pós-sináptica
no modelo animal. Interessante notar que durante o desenvolvimento do sistema GABAérgico
cortical o neurotransmissor GABA deixa de ser excitatório (durante o período embrionário até
início pós-natal) e passa a ser inibitório a partir de P7-P10 (Rivera et al.,1999). Este fenômeno
se deve à diminuição da concentração do íon Cl− no meio intracelular por meio da expressão do
1.6. Comunicação hipocampo-córtex pré-frontal medial 10
foi mostrado que na área CA3 do hipocampo, a transição entre o papel excitatório-inibitório
do GABA está alterada em animais VPA (Tyzio et al., 2014). Nos neurônios embrionários, a
maior concentração de Cl− no meio intracelular no hipocampo controle força o fluxo de Cl− em
resposta a ativação de receptores GABA-A (efeito excitatório). Este gradiente, entretanto, sofre
uma queda abrupta e pontual no primeiro dia pós-natal (P0) em decorrência da liberação de
oxitocina (redução do efeito excitatório do GABA) e, finalmente, volta a ser reduzida aos níveis de
animal adulto entre os dias pós-natal 15 e 30 (P15 e P30).Tyzio et al.(2014) mostraram que esta
sequência não ocorre nos animais VPA, sugerindo que a disfunção da homeostase da sinalização
GABAérgica durante o desenvolvimento pós-natal poderia gerar uma circuitaria hiper-excitável
no hipocampo, onde os autores estudaram, e os comportamentos alterados no modelo VPA.
1.6
Comunicação hipocampo-córtex pré-frontal medial
Disfunções na comunicação entre regiões corticais de pacientes autistas parecem estar associada
a grandes dificuldades destes indivíduos em integrar informações e interpretar estímulos externos
(Matlis et al.,2015;Wolf et al.,2009). O fato de crianças autistas serem incapazes de interagir
reciprocamente com o seu meio social pode refletir uma deficiência do córtex pré-frontal em
interagir reciprocamente com outras regiões corticais (Godsil et al., 2013; Lewis et al.,2014).
Tem sido descrito que pacientes autistas apresentam disfunções no córtex pré-frontal e no lobo
temporal. Alguns estudos mostraram alterações anatômicas e moleculares nestas áreas (
Cour-chesne et al.,2011;Ziats e Rennert,2013). Ambas regiões fazem parte da DMN que apresenta
conectividade funcional entre seus nós reduzida nesses indivíduos (Assaf et al.,2010).
Recentemente alguns estudos têm mostrado alterações na neurotransmissão basal e
plasti-cidade sináptica entre o hipocampo e o córtex pré-frontal no modelo VPA (Sui e Chen, 2012;
Martin e Manzoni,2014). Entretanto estes estudos foram realizados em animais anestesiados e
nenhum deles abordou o acoplamento funcional entre estas duas áreas durante tarefa de natureza
social (Codagnone et al.,2015). O córtex pré-frontal é a segunda principal saída de informações
do hipocampo em direção ao córtex. Sua conectividade com o hipocampo é monossinaptica e
fundamental para a execução de tarefas de memória de trabalho, e plasticidade sináptica
envol-vida na formação de memórias de longo prazo (Jay et al.,2004;Romcy-Pereira e Pavlides,2004).
O acoplamento entre essas regiões foi inicialmente descrito em diferentes estados
1.7. Medo e extinção do medo 11
Estes autores mostraram que neurônios registrados no CPFm disparam preferencialmente em
uma determinada fase do ritmo teta hipocampal (Siapas et al.,2005). Além disso, a coerência
entre oscilações teta das duas regiões é elevada durante tarefas de tomada de decisão e cunho
emocional (OŠNeill et al.,2013). O interessante é que animais com maiores taxas de acerto na
tarefa apresentaram maior coerência entre as áreas, sugerindo que o acoplamento entre o CPFm
e o hipocampo está associado ao aprendizado. Estudos recentes têm mostrado que modelos de
ansiedade e esquizofrenia apresentam redução tanto na coerência em teta quanto no acoplamento
entre essas duas áreas (Sigurdsson et al.,2010).
1.7
Medo e extinção do medo
A dificuldade de interpretação e expressão de mensagens de caráter emotivo são características
comuns em pacientes autistas (Boraston et al.,2007;Charbonneau et al.,2013;Sato et al.,2012).
Em particular, a identificação e manifestação de medo é bastante alterada. Esses indivíduos
apresentam fobias (Mayes et al.,2013), problemas em identificar expressões de medo ou de raiva
(Wallace et al.,2008), condicionamento ao medo exacerbado (South et al.,2011) e dificuldades
em esquecer eventos de carga emocional aversiva (Gaigg et al.,2008).
O pequeno número de estudos psicofisiológicos examinando paradigmas de medo condicionado
clássico em seres humanos com transtorno do espectro autista têm rendido resultados discordantes
(Salmond et al.,2003; Gaigg et al.,2008; South et al.,2011; Bernier et al.,2005). Uma parte
destes estudos reportam diferenças entre a aquisição de medo condicionado entre indivíduos
autistas e indivíduos controle, enquanto outra não reporta nenhuma diferença. Vale salientar
que um dos grandes problemas em estudos com humanos é a grande heterogeneidade encontrada
entre indivíduos com transtornos do espectro autista.
Em ratos foi descrito que animais tratados com ácido valpróico são mais ansiosos e apresentam
memórias ligadas ao medo com níveis anormalmente elevados e de maior persistência, com
tendência a sobre-generalizar o medo a diferentes estímulos sensoriais e maior dificuldade de
extinguir ou esquecer (Markram et al.,2008).
1.8
Vocalizações ultrassônicas de ratos
Os ratos são capazes de vocalizar e ouvir dentro de uma faixa de frequência cujos limites são
1.8. Vocalizações ultrassônicas de ratos 12
kHz, os roedores podem ouvir sons de frequência superiores a 50 kHz. Diversos estudos apontam
para grande relevância etológica das vocalizações como sinais sociais para os ratos (Knutson
et al.,2002;Brudzynski,2005,2013;Burgdorf et al.,2008). Por isso o estudo dessas vocalizações e
de suas representações neuronais constitui uma ferramenta bastante interessante para a pesquisa
sobre comunicação, emoção e interação social em ratos.
Os ratos adultos desenvolvem dois tipos básicos de vocalizações ultrassônicas (VUS) que são
denominados genericamente Ş20 kHzŤ e Ş50 kHzŤ (Brudzynski,2013;Wöhr e Schwarting,2013).
As vocalizações de 20 kHz são emitidas durante estados emocionais negativos e situações aversivas,
enquanto chamadas de 50 kHz são emitidas em estados emocionais positivos (Brudzynski e Pniak,
2002;Knutson et al.,2002).
Tem sido sugerido que um dos papéis das vocalizações de 20 kHz é o de alertar a atenção de
toda a colônia de coespecíficos acerca da presença de predadores (Blanchard et al.,1991;Litvin
et al.,2007) ou outro perigo (Brudzynski e Ociepa,1992;Brudzynski,2001). As vocalizações de
22kHz são também correlacionadas com o comportamento de congelamento (do inglês,freezing)
e outros indicadores de medo (Lee et al.,2001;Choi e Brown,2003).
Por meio de experimentos de playback foi mostrado que os chamados de 20 kHz podem
induzir inibição locomotora (Wöhr e Schwarting,2007) e a atividade neuronal em áreas do cérebro
envolvidas na regulação do medo, como a amígdala e a substância cinzenta periaquedutal(Wöhr
e Schwarting,2013). Descobertas recentes indicam ainda que os ratos estão predispostos a formar
associações de memória entre 22kHz e estímulos aversivos (Allen et al.,2007;Endres et al.,2007;
Bang et al.,2008;Furtak et al.,2007) e que esses chamados desempenham um papel importante
na transmissão social do medo (Kim et al.,2010).
As vocalizações de alta frequência (50kHz) ocorrem em contextos comportamentais naturais
de conteúdo emocional positivo, incluindo jogo juvenil (Knutson et al.,1998), atividade social
exploratória (Brudzynski e Pniak,2002; Assini et al., 2013), acasalamento(McGinnis e
Vaku-lenko,2003) e antecipação de estímulos gratificantes (Knutson et al.,1998). Estas vocalizações
também são emitidas na consolidação de contatos sociais (ou seja, interações não-combativas
de abordagem, proximidade individual e afeto) e na antecipação de tais contatos (Brudzynski
e Pniak,2002; Willey e Spear, 2012;Willey et al.,2009). Eles podem desempenhar um papel
no aumento da coesão do grupo social e cooperação (čopuch e Popik,2011) e na estimulação e
coordenação de comportamento social (Thomas et al.,1983; Galef e Jeimy, 2004;Brudzynski,
1.8. Vocalizações ultrassônicas de ratos 13
Contrariamente aos chamados de 20kHz, o playback de chamados de 50kHz tende a aumentar
a atividade locomotora do animal e causar comportamentos aproximativos (Wöhr e Schwarting,
2007; Burgdorf et al.,2008). Também foi mostrado que ratos treinados para escolher um tipo
de vocalização efetuam a autoadministração de playback dos chamados de 50kHz de frequência
modulada e evitam o playback de vocalizações de 20 kHz (Burgdorf et al.,2008). Uma vez que
estes chamados de 50kHz são expressos durante a antecipação da cópula (McIntosh et al.,1978;
Barfield e Thomas,1986;Bialy et al.,2000) , jogo e brincadeira (Knutson et al.,1998), antecipação
de estímulos gratificantes e no contexto de auto-estimulação elétrica do feixe prosencefálico medial
(Burgdorf et al.,2000), foi postulado que estes chamados são um marcador sensível para respostas
não-condicionadas e condicionadas positivas (Knutson et al.,1999) ePanksepp e Burgdorf(2003)
Capítulo 2
Justificativa
O modelo experimental de utilizando VPA durante o período gestacional tem sido demonstrado
ter alta validade de face e permitir estudos tanto das bases neuropatológicas quanto
neuro-funcionais durante o desenvolvimento. A despeito do recente interesse por este modelo como
instrumento de compreensão dos aspectos básicos da fisiopatologia do autismo, a maioria dos
estudos experimentais têm se concentrado nos aspectos comportamentais, histológicos e
celula-res. Recentemente alguns estudos eletrofisiológicos mostraram alterações eletrofisiológicas em
animais anestesiados. Entretanto, ainda se faz necessário uma avaliação eletrofisiologia animais
durante diferentes estados comportamentais e interações sociais. Portanto, neste estudo
preten-demos contribuir para o preenchiemnto desta lacuna através da implementação de estratégias
experimentais de avaliação comportamental associadas a eletrofisiologia in vivo, além de avaliar
o acoplamento funcional entre o hipocampo e o CPFm no modelo VPA de autismo em animais
Capítulo 3
Objetivos
Este trabalho tem como objetivo a implementação de estratégias metodológicas para o estudo
de comportamentos sociais em animais modelo de autismo, bem como, caracterizar, por meio de
registros eletrofisiológicos extracelulares, a comunicação entre o hipocampo e o córtex pré-frontal.
3.1
Objetivos específicos
• Planejamento e organização desetupexperimental para registro de vocalizações simultâneo
a registros eletrofisiológicos e de vídeo;
• Planejamento e organização desetup para realização de um protocolo de condicionamento
ao medo. Na fase se extinção da memória aversiva, os estímulos sonoros são sincronizados
ao registro eletrofisiológico;
• Registrar potenciais locais de campo (LFP) no córtex pré-frontal e no hipocampo
simulta-neamente de animais VPA e controle;
• Avaliar os padrões oscilatórios hipocampais e corticais basais durante exploração de um
ambiente novo;
Capítulo 4
Materiais de Métodos
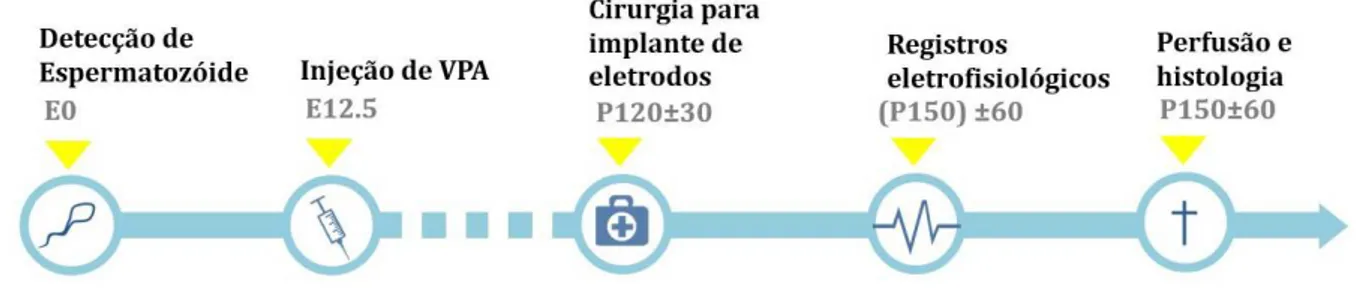
A linha temporal dos experimentos está representada simplificadamente na figura4.1. Animais
(P120±30) tratados com ácido valpróico e controles foram submetidos a cirurgias de implantes
de matrizes de eletrodos no hipocampo e no córtex pré-frontal medial (área pré-límbica). Todos
os animais tiveram ao menos sete dias de recuperação após a cirurgia e a partir de então foram
habituados ao experimentador por pelo menos mais 5 dias antes de serem submetidos aos testes
comportamentais. O conjunto de testes realizados foram: atividade exploratória, interação social,
condicionamento ao medo e extinção. Com exceção do condicionamento ao medo, durante todos
os outros experimentos foram registrados potenciais de campo locais (LFP) no hipocampo e no
córtex pré-frontal.
Figura 4.1: Esquema temporal do desenho experimental.
4.1
Modelo animal de autismo
Utilizamos ratos Wistar machos e fêmeas mantidos no Biotério Setorial do Instituto do Cérebro
da Universidade Federal do Rio Grande do Norte (UFRN). Fêmeas com ciclos de fertilidade
con-4.2. Confecção de matrizes de eletrodos 17
sanguineamente durante o período noturno (ou período de atividade). Na manhã seguinte ao
acasalamento, a fecundação e a determinação do primeiro dia gestacional (E0) foi confirmada
pela presença de espermatozóides na lavagem vaginal.
Passados 12 dias da fertilização (E12,5), data correspondente ao final do fechamento do tubo
neural nestes animais, um grupo de fêmeas recebeu uma injeção intraperitoneal de valproato de
sódio (VPA, 500 mg/kg, i.p.; Sigma, P4543) dissolvido em solução salina na concentração de 250
mg/mL. A administração de VPA na dose 500 mg/kg tem sido mostrada atingir níveis séricos
máximos da ordem de 900µg/mL em menos de 1 h, com meia-vida de eliminação plasmática
de 2,3 h (Binkerd et al., 1998). Essas proles foram utilizadas como grupo VPA. Outro grupo de
fêmeas grávidas receberam uma única injeção de salina em E12,5 e seus filhotes foram utilizados
como animais controles.
Após o nascimento, a prole permanece com a mãe por 21 dias durante a amamentação.
Passado este período, os animais machos são transferidos para gaiolas em grupos de dois a
quatro, onde ficam mantidos até a realização dos procedimentos experimentais. Todos os animais
são alojados em gaiolas de polipropileno (414 x 344 x 168 mm3) com raspa de madeira no assoalho
numa sala com temperatura (21±2°C), umidade e ciclo de claro-escuro (de 12-12h; ligado às 7:00h
e desligado às 19:00h) controlados com livre acesso à comida e água. Todos os procedimentos
experimentais realizados com os animais foram previamente aprovados pela Comissão de Ética
no Uso de Animais em Pesquisa (CEUA) da UFRN, sob o protocolo no. 044/2011. Apenas
animais machos foram utilizados no testes experimentais.
4.2
Confecção de matrizes de eletrodos
Para registrarmos potenciais locais de campo, utilizamos matrizes de eletrodos que foram
confecci-onadas em nosso laboratório no Instituto do Cérebro. Para tal, foram utilizados fios de tungstênio
de 50 µm de diâmetro revestidos com teflon. Os eletrodos foram alinhados verticalmente em duas
fileiras de 4 eletrodos de maneira que o espaçamento entre eles fosse de 300 µm (figura 4.2a).
O manuseio dos eletrodos foi feito com bastante cautela afim de evitar danos ao revestimento
e somente a ponta do eletrodo foi deixada exposta para aquisição o sinal elétrico intracerebral.
Por fim, as pontas opostas foram conectadas a um conector Würth (1,27mm,dual-socket 18
pinos de ouro) através de uma mini placa de circuito impresso de produção do nosso laboratório
4.3. Cirurgia para implante de eletrodos 18
(a) (b)
Figura 4.2: Matrizes de eletrodos.(a) as matrizes 2x4 implantadas no CPFm e no hipocampo foram confeccionadas com eletrodos de tungstênio 50µm e espaçamento de 300µm; (b)Foto de
uma matriz utilizada nos implantes.
Os eletrodos foram submetidos ao processo de galvanização por eletrólise em uma solução
de ouro contendo nanotubos de carbono, com intuito de reduzir as impedâncias para faixa entre
50 e 100 kΩsegundo descrito emFerguson et al.(2009). Nesse processo, as pontas dos eletrodos
são imersas em uma solução de ouro enquanto uma corrente negativa os atravessa. Este fluxo de
elétrons faz com que o ouro se deposite nas pontas dos eletrodos. A galvanização, bem como as
medições de impedâncias de cada eletrodo, foram realizadas utilizando o equipamento NanoZ
(Neuralynx, USA). O equipamento gera microcorrentes para depósito do ouro e em seguida envia
sinusoidais de 1 KHz para avaliação das impedâncias. Este processo é repetido algumas vezes
até a obtenção dos valores desejados.
4.3
Cirurgia para implante de eletrodos
Ratos machos (idade 120±30 dias) experimentais e controles foram submetidos a cirurgias de
implante de duas matrizes de eletrodos 2x4 (fios de tungstênio revestidos comteflon de 50µm;
espaçamento horizontal: 300µm; impedância: ∼70 kΩ à 1 kHz). Uma matriz foi implantada no
hipocampo dorso-posterior (AP= -5.8; DV=3,2; ML=4,5) e a segunda matriz implantada no
CPFm (AP= +3,6; DV=3,8; ML=0,6). Neurônios dessas regiões se conectam
monossinaptica-mente (Hoover e Vertes,2007).
Para a cirurgia, os animais foram previamente anestesiados com uma mistura de cloridrato de
cetamina (100 mg/kg, i.p.; Agener) e cloridrato de xilazina (8mg/kg, i.p.) e, foram mantidos com
a temperatura corporal controlada na faixa de 37±2°C. Para manutenção, foram administradas
im-4.3. Cirurgia para implante de eletrodos 19
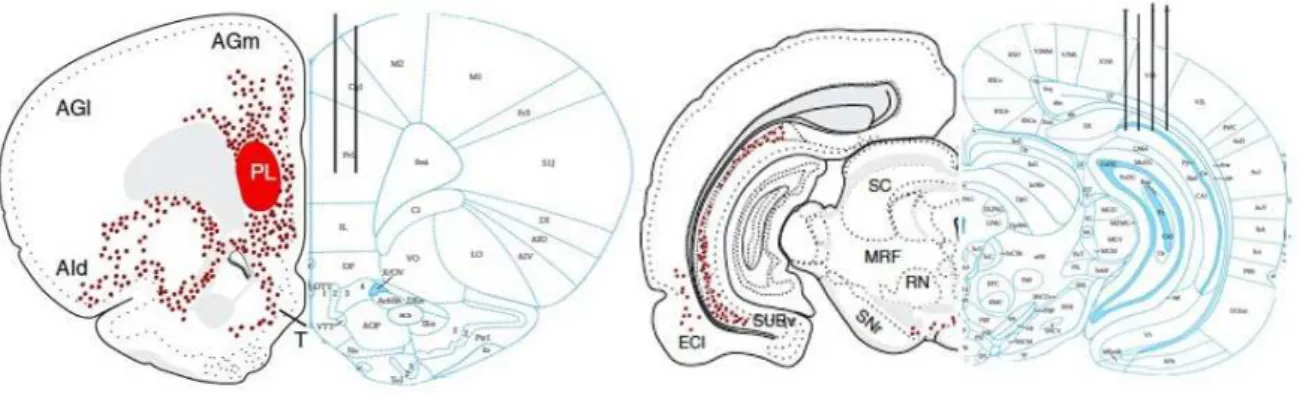
Figura 4.3: Representação do corte coronal do cérebro de um rato Wistar nas coordenadas (AP= -5.8; DV=3,2; ML=4,5) à direita e (AP= +3,6; DV=3,8; ML=0,6) à esquerda. Ambos hemisférios esquerdos apresentam em vermelho marcador retrógrado aplicado no córtex pré-frontal medial pré-límbico (Hoover and Vertes, 2007) mostrando conexão monossináptica entre as duas regiões onde foram implantados os eletrodos. As localizações dos implantes estão ilustradas nos hemisférios esquerdos dos cortes ilustrados.
plante foram realizados como previamente descrito no protocolo da CEUA, com todos os cuidados
necessários para garantir assepsia e ausência de dor (i.e., uso de materiais autoclavados, antibiótico
e analgésico pós-cirúrgico). Mais especificamente, no pré-operatório foi injetado um antibiótico
(Flotril® injetável 2,5%) intra-muscularmente, cloridrato de lidocaína sub-cutâneamente para
anestesia local da região de incisão e também injetado o anti-inflamatório/analgésico Maxicam
0.2%. Além disso, durante a cirurgia também injetamos salina (NaCl 0.9%) sub-cutâneamente
para prevenção de desidratação e mantivemos os olhos do animal úmidos com gel lubrificante. A
cabeça do animal foi desinfetada topicamente com Iodopovidona 10% antes da incisão e, após a
mesma, o crânio foi limpo com peróxido de hidrogênio 3%. Após a cirurgia de implante, aplicamos
na região da sutura uma pomada com propriedades antibióticas, anti-inflamatórias, anti-alérgicas,
anti-pluriginosas, anestésicas e anti-micóticas (Crema 6A, contendo dexametasona, neomicina,
bacitracina, griseofulvina e benzocaina). Finalizado o implante, os animais foram monitorados
até recobrirem atividade locomotora e então, devolvidos ao biotério onde ficaram em recuperação
por pelo menos uma semana antes do início dos registros eletrofisiológicos. Após o período de
4.4. Acoplamento hipocampo-CPFm durante atividade exploratória e interação social 20
4.4
Acoplamento hipocampo-CPFm durante atividade
explora-tória e interação social
Segundo dados anteriormente obtidos em nosso laboratório, ratos expostos ao VPA durante a
gestação apresentam alterações comportamentais caracterizadas por redução de interesse social,
aumento de comportamentos estereotipados e hiperatividade se comparados a animais controles
(Sousa,2013). A interação com um animal não-familiar leva à formação de uma memória social
em roedores. Durante este processo, tanto o hipocampo quanto regiões corticais são recrutadas
para a codificação e posterior consolidação desta memória episódica. Tendo isto em consideração,
nosso interesse foi avaliar a atividade oscilatória hipocampal e pré-frontal durante atividade
exploratória e interação social. Além disso, mesurar o grau de sincronização entre as duas áreas.
O experimento foi realizado com três ratos controles e três ratos experimentais implantados
com eletrodos no CPFm e no hipocampo. Em sessões individuais, cada animal foi conectado
ao cabo de registro (contendo um pré-amplificador Űheadstage) e submetidos aos paradigmas
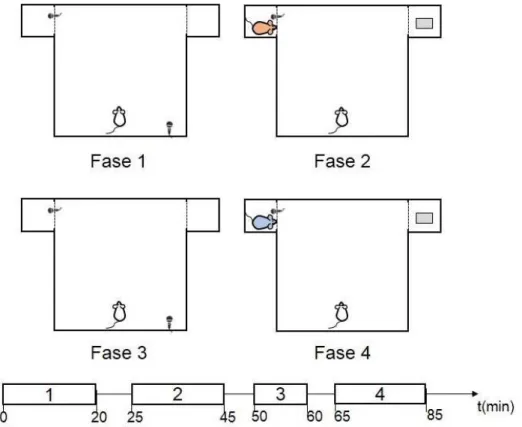
descritos nesta seção. Este experimento é realizado em uma sequência de quatro fases distintas
numa arena retangular principal (86x57x40cm3) com dois pequenos compartimentos laterais
(17x17x40 cm3) (figura4.4). Nas fases 1 e 3, os compartimentos laterais permaneceram vazios e
o animal pode explorar livremente por toda arena principal. As divisórias entre a arena principal
e os compartimentos auxiliares foram confeccionadas em madeira com revestimento preto similar
às paredes da arena principal. Já nas fases 2 e 4, um animal familiar ou não-familiar é colocado
em um dos compartimentos e um objeto branco de tamanho similar a um rato Wistar no outro.
Neste caso, as divisórias utilizadas entre a arena principal e os compartimentos auxiliares foram
confeccionadas de acrílico transparente com orifícios circulares de 1,5 cm espaçados por 2 cm
para permitir a interação entre os animais. A sequência de apresentação de animal familiar ou
não-familiar foi randomizada, bem como o lado do compartimento em que são colocados. As
quatro fases são separadas por intervalos de cinco minutos quando o animal é retirado da arena
principal e posto em sua caixa de origem. Com exceção da terceira fase que tem duração de 10
minutos, todas as fases têm duração igual a 20 minutos.
Os animais familiares são ratos Wistar pareados por sexo e idade e compartilham a mesma
gaiola com os animais experimentais por pelo menos 5 dias antes do início dos experimentos de
interação social. As gaiola de manutenção dos animais no biotério são divididas ao meio com
4.4. Acoplamento hipocampo-CPFm durante atividade exploratória e interação social 21
Figura 4.4: O experimento é dividido em quatro fases. Em todas as fases, o animal pode explorar o compartimento central livremente. Nas fases 1 e 3, os compartimentos laterais encontram-se vazios. Já nas fases 2 e 4, um dos compartimentos comporta um animal e o outro um objeto branco de tamanho similar a um rato. A linha temporal do experimento é ilustrada na parte de baixo da figura.
maneira, os animais podem interagir através de vocalizações, pistas olfativas e contato visual,
sem risco de danos ao capacete do animal implantado. Os animais não familiares são ratos Wistar
pareados por sexo e idade pertencentes a outras gaiolas também mantidos em pares.
Por pelo menos 5 dias antes do dia de experimentação, os animais foram manuseados
diari-amente pela manhã por pelo menos 15 minutos cada. Durante as sessões experimentais, foram
realizados registros eletrofisiológicos contínuos, registro de áudio ultrassônicos e filmagem do
comportamento sincronizada com os registros. A iluminação do ambiente foi realizada por um
conjunto de leds infra-vermelhos. Após o teste, os animais retornaram às suas caixas de origem
e foram levados ao biotério. O aparato comportamental foi limpo com uma toalha umedecida
com água.
A aquisição de dados neurais foi realizada através do sistema Omniplex (Plexon Inc.)
utili-zandoheadstagede alta impedância de entrada com ganho de 20X (HST/32V-G20). Os registros
eletrofisiológicos e os dados comportamentais de posicionamento do animal foram sincronizados
4.4. Acoplamento hipocampo-CPFm durante atividade exploratória e interação social 22
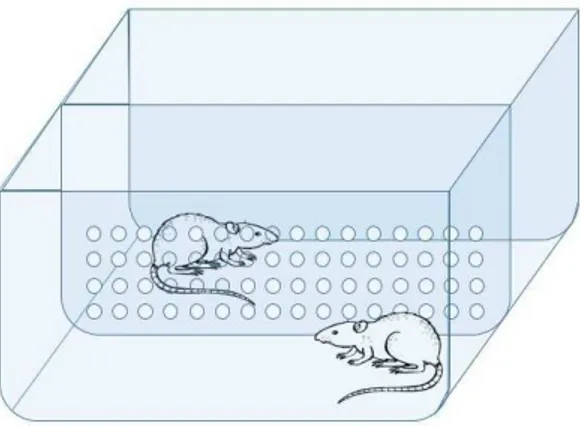
Figura 4.5: Os animais familiares dividiram gaiola com os animais experimentais durante 3 dias anteriores aos experimentos. A gaiola era dividida por uma placa de acrílico transparente e com orifícios circulares de 1,5 cm de diâmetro
CM16/CMPA foram utilizados para registrar as emissões vocais dos animais. A posição dos mesmos foi pensada de maneira a possibilitar futura identificação do rato emissor. Conforme
ilustrado na figura 4.6, um deles foi colocado em direção ao animal aprisionado no pequeno
compartimento enquanto a direção do segundo foi paralela a parede mais longa do aparato de
maneira que o som emitido pelo animal aprisionado chegue bastante atenuado a entrada deste
microfone.
Figura 4.6: Esquema representativo dosetupdo experimento de interação social. Utilizando um
sistema de aquisição de dados NI USB-6531, os registros ultrassônicos dos microfones foram sincronizados aos registros de vídeo e eletrofisiológicos do sistema daPlexon.
4.5. Condicionamento pelo medo 23
desenvolvido a fim de sincronizar o início dos registros de áudio aos registros de vídeo e neurais.
Quando executado no notebook, um pulso elétrico era enviado a uma saída digital do sistema de
aquisição de dadosNational Instruments NI USB-6351 concomitantemente ao início do registro
das vocalizações. A saída digital doNI USB-6531 foi conectada a entrada digital daPlexon e,
desta forma, podemos sincronizar os registros sonoros aos registros de vídeo e eletrofisiológicos
posteriormente.
4.5
Condicionamento pelo medo
O condicionamento pelo medo é um paradigma comportamental bastante utilizado para estudar
essa emoção em modelos animais. Portanto, se apresenta como um bom mecanismo de estudo
de alterações emocionais presentes no autismo. Este protocolo de condicionamento ao medo que
descreveremos nesta seção foi testado em dois animais sendo um controle e outro modelo de
autismo. Neste paradigma, um estímulo sonoro previamente não aversivo (CS+) é apresentado
algumas vezes em conjunto a um choque de baixa intensidade ao animal. O animal, então, passa
a demonstrar reações características de medo (congelamento, aumento de frequência cardíaca)
ao ouvir aquele som mesmo quando não associado a um choque. Em indivíduos com estresse
pós-traumático e ansiedade generalizada existe uma grande probabilidade de ŞgeneralizaçãoŤ
comportamental a estímulos não aversivos diversos. A generalização se refere ao fato da resposta
comportamental ser exacerbada mesmo quando seguindo um estímulo inofensivo que não tenha
sido previamente apresentado em conjunto com estímulos aversivos.
Os experimentos de condicionamento discriminativo pelo medo são realizados em um aparato
cujo assoalho consiste em barras de metal por onde é dado choques de intensidade em torno de
0,3 mA. Com exceção da parede frontal da caixa que é transparente, todas as paredes são opacas
em tonalidade cinza. Após cada sessão as caixas foram limpas com solução de ácido acético 1%.
Dois estímulos sonoros de diferentes frequências foram apresentados, um deles pareado ao choque
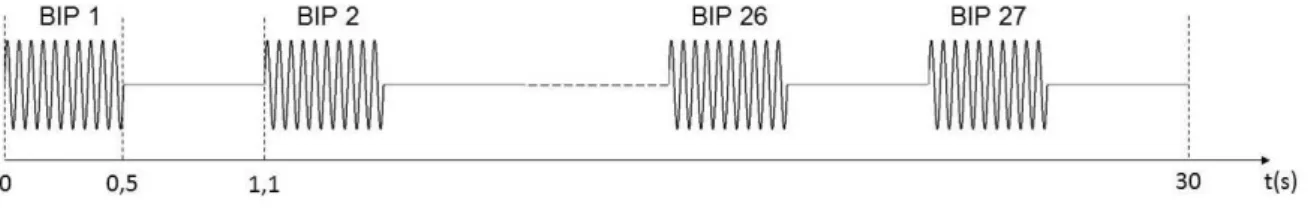
(CS+) e o outro não (CS-). Cada estímulo com duração de 30 segundos em uma sequência de
27 bips de 0,5 s (ver figura4.7). No caso do CS+, o últimobip foi co-terminado com um choque.
Osbips eram compostos por tons puros de 4 kHZ (CS+) e 10kHz (CS-).
O animal foi colocado no aparato e pôde explorar a caixa livremente por 2 minutos. Após
esse período, foram apresentados os estímulos sonoros cuja ordem de apresentação é a seguinte:
4.6. Extinção do medo 24
Figura 4.7: Cada estímulo sonoro, CS+ ou CS-, era composto por 27 bips sequenciais. A duração de cada um era de 500 ms e se repetiam a cada 1100 ms. A duração total de cada estímulo era de 30 s.
o experimento, foram realizados registros de VUS através de microfone ultrassônico e registro
de vídeo. No entanto, registros eletrofisiológicos não foram feitos para evitar danos ao
pré-amplificador presente noheadstage em decorrência da corrente elétrica aplicada no animal.
4.6
Extinção do medo
Um dia após o condicionamento pelo medo, realizamos a extinção dessa memória aversiva através
da apresentação seguidas vezes do estímulo sonoro CS+ sem estar pareado com o choque. A
extinção do medo foi realizada numa arena cujas características são bem diferentes daquelas
da arena do condicionamento. Suas paredes e assoalho são pretos, de madeira e limpos com
álcool 70%. Apresentamos os mesmos estímulos CS+ e CS- utilizados no condicionamento. A
apresentação dos CS- é importante para avaliarmos a ocorrência ou não a generalização. O
animal foi deixado explorar a caixa livremente por um período de dois minutos, e a seguir foi
apresentada uma sequência de quatro CS- seguidos por doze CS+. O intervalo de tempo entre
cada CS variou entre 30 e 120 segundos aleatoriamente. Durante o experimento, foram realizados
registros eletrofisiológicos contínuos, registro de áudio ultrassônicos e filmagem sincronizada com
os registros. A iluminação do ambiente foi realizada por um conjunto de leds infra-vermelhos.
Após o teste, os animais retornaram às suas caixas de origem e foram levados ao biotério.
4.7
Registro eletrofisiológico
A aquisição de dados foi realizada através do sistema Omniplex (Plexon Inc., USA)utilizando
headstage de alta impedância de entrada com ganho de 20X (HST/ 32V-G20). Os registros
eletrofisiológicos e os dados comportamentais de posicionamento do animal e velocidade de
locomoção foram sincronizados através do sistemaCinePlex (Plexon Inc., USA). O sinal de cada
4.8. Análise de dados 25
kHz para para análise de LFPs.
4.8
Análise de dados
A análise de dados foi realizada atavés do softwareCinePlex e de rotinas escritas por nós para
Matlab 2013b (MathWorks, Natick, MA).
4.8.1 Pré-tratamento do sinal
Utilizamos oCinePlex para realização das primeiras análisesoff-line dos dados. Essa ferramenta
permite visualização dos sinais registrados através do sistemaOmniplex em sincronia com o vídeo.
Alguns canais foram previamente excluídos das análises por sofrerem bastante interferência de
ruídos eletromagnéticos e/ou de artefatos de movimento. Apesar da medição pré-operatória de
impedância de todos os eletrodos, o manuseio da matriz durante a cirurgia pode danificar a
conexão dos fios de tungstênio à placa de circuito impresso ou até mesmo partir os eletrodos.
Através doCineplex é possível definir marcadores de eventos de interesse. Definimos, então,
intervalos onde apareciam artefatos de movimento nos canais não-excluídos. Esses marcadores
foram utilizados em rotinas do MatLab para seleção dos trechos analisados e mostrados nos
Resultados deste trabalho. Além disso, identificamos e sinalizamos uma série de
comportamen-tos: atividade exploratória, repouso, interação com animal e interação com objeto. Atividade
exploratória foi definida como movimentação de cabeça, pescoço e vibrissas mesmo se o animal
não estivesse propriamente ambulando. Repouso foi definido como imobilidade sem sem
pros-tração do corpo em posição de sono. Por fim, os trechos de interação foram determinados pela
aproximação do focinho do animal experimental com a grade que o separava do outro animal
(interação social) ou do objeto (interação com objeto).
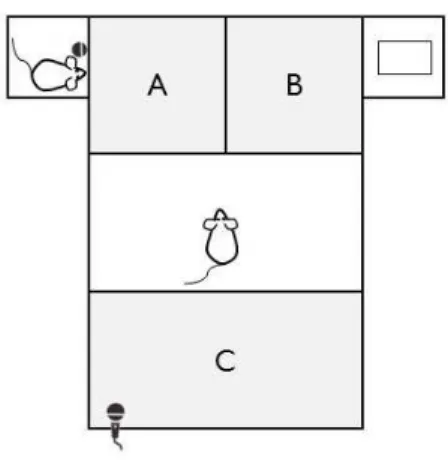
Definimos três zonas distintas dentro do aparato que foram diferenciadas pela distância às
regiões de interação com animal e objeto. As duas primeiras zonas A e B, adjacentes a gaiola
do animal e do objeto, respectivamente, foram estabelecidas como áreas quadradas com lado
de 28,5 cm. A zona C, localizada no lado oposto a ambas gaiolas, possuía área retangular de
4.8. Análise de dados 26
Figura 4.8: Três zonas foram delimitadas dentro do aparato: A, B e C. As duas primeiras, A e B, são quadradas com vértices iguais a 28,5 cm e a segunda retangular de área igual a 57x28,5 cm2. A zona A é adjacente a gaiola do animal de exposição, a zona B é adjacente a gaiola do objeto e, por fim, a zona C fica no lado oposto a ambas gaiolas.
4.8.2 Análise de LFPs
Densidade espectral de potência (PSD): Foi calculada através do método de Welch com janelas
Hamming de 1024 pontos e sobreposição de 50% do segmento de LFP. A normalização da PSD
foi resultado da divisão de cada ponto da PSD bruta pela soma de todos os pontos da PSD
excluindo aqueles correspondentes a faixa de frequência entre 58 e 62 Hz. Essa faixa de frequência
foi excluída das análises pois em alguns registros possuía muita energia proveniente de ruído da
rede de energia elétrica local.
Coerência: A coerência é uma medida da correlação entre duas séries temporais. Em
neurofisi-ologia, onde dados eletrofisiológicos de campo (LFP) são registrados simultaneamente em várias
áreas, podemos usá-la como parâmetro estimativo da sincronia entre regiões cerebrais. Neste
relatório, utilizamos o método de Welch para cálculo da coerência entre o CPFm e o hipocampo
com janelamento hamming de 1024 pontos,overlapde 50% e Transformada Rápida de Fourier
Capítulo 5
Resultados
5.1
Exploração de um novo ambiente
Três animais controles e três animais experimentais tendo sido manipulados por no mínimo
cinco dias antecedentes foram deixados explorar livremente um novo ambiente durante a fase 1
do experimento descrito em4.4. Durante este período, foram registrados potenciais de campo
local (LFP) simultaneamente no córtex pré-frontal e no hipocampo ventral. Para realização
das análises dividimos o experimento em dois momentos: o primeiro enquanto o animal esteve
ativo em atividade de exploração com as vibrissas em movimento; e o segundo quando animal
esteve em repouso, no entanto, sem dormir. Para realização das análises mostradas nesse capítulo,
utilizamos o melhor canal de registro do hipocampo e o melhor do córtex pré-frontal, isto é,
aqueles menos sujeitos a ruídos elétricos e artefatos de movimento.
Podemos ver na figura 5.1, trechos representativos de LFPs de animais controles (5.1a e
5.1b) e animais VPA (5.1c e5.1d) em momentos de exploração. Já é bem descrito que durante
atividade locomotora, os ratos apresentam oscilação hipocampal em ritmo teta (entre 4 e 12 Hz)
sustentada. Mesmo através da representação do sinal no domínio do tempo, podemos observar
que assim como o grupo controle, o grupo VPA também apresenta aumento da potência na faixa
de teta. Percebe-se que a oscilação mais lenta de maior potência alcança seu valor de pico em
torno de 8 vezes dentro do período de 1 s representado, correspondendo a frequência de teta. Essa
característica fica mais clara, no entanto, quando observamos os dados no domínio de frequência.
A densidade espectral de potência normalizada apresentada na figura5.2refere-se ao conjunto
de trechos extraídos em que os animais estiveram em locomoção. A faixa de frequência da rede
5.1. Exploração de um novo ambiente 28
(a) (b)
(c) (d)
Figura 5.1: Trechos ilustrativos de LFP de animais controle (em preto) registrados no(a)Córtex pré-frontal medial e no(b) hipocampo; e animais VPA (em vermelho) registrados no(c)córtex pré-frontal medial e no(d)hipocampo. Os animais estavam em atividade exploratória
ao grupo controle e aqueles em vermelho ao grupo VPA. Sendo ambos da esquerda extraídos do
córtex pré-frontal medial, enquanto aqueles da direita vêm do hipocampo. Vemos que os dois
grupos apresentam um pico na faixa de teta em ambas regiões cerebrais. Outro pico em torno
de 18 Hz é presente tanto no hipocampo como no pré-frontal medial do grupo controle. Por
fim, notamos um pico na faixa de high gamma (62 Ű 90 Hz) no córtex pré-frontal dos animais
controle.
Figura 5.2: PSD do LFP dos animais em atividade exploratória. Os gráficos em cinza corres-pondem ao grupo controle (n=3) e aqueles em vermelho ao grupo VPA (n=3). Sendo ambos da esquerda extraídos do córtex pré-frontal medial, enquanto aqueles da direita vêm do hipocampo.
5.1. Exploração de um novo ambiente 29
em repouso em momento algum. Portanto, para as analises dos momentos em repouso, somente
dois animais de cada grupo foram utilizados. Vemos na figura5.3trechos representativos de LFPs
desses momentos e podemos notar que nesse caso as ondas são mais chatas. A potência de teta
claramente se mostra reduzida neste caso. No entanto, esse fato fica mais nítido se compararmos
as figuras5.4e 5.2que representam a PSD normalizada para os dois momentos distintos.
(a) (b)
(c) (d)
Figura 5.3: Trechos ilustrativos de LFP de animais controle (em preto) registrados no(a)Córtex pré-frontal medial e no(b) hipocampo; e animais VPA (em vermelho) registrados no(c)córtex pré-frontal medial e no(d)hipocampo. Os animais estavam em repouso.
5.2. Animais em interação social 30
5.2
Animais em interação social
Nesta seção abordaremos os resultados provenientes da etapa do experimento de interação social
(seção 4.4) quando cada animal fora colocado na arena principal enquanto um animal familiar
estava presente em um compartimento lateral da mesma. Os dois espaços eram separados por
uma parede de acrílico transparente e com orifícios circulares de 1,5 cm ao longo de toda ela.
Desta forma, os animais podiam interagir através da visão, do olfato e de vocalizações. Somente
dados provinientes de dois animais de cada grupo foram utilizados nessas análises. Durante esse
período, separamos os trechos de interação com o animal familiar conforme explicado em4.8.1.
Podemos ver na figura5.5, trechos representativos de LFPs de animais controles (5.5ae 5.5b) e
animais VPA (5.5c e 5.5d) nesses momentos. Ao analisarmos a PSD dos dois grupos durante a
interação social (figura5.6), não observamos nenhuma diferença significativa.
(a) (b)
(c) (d)
Figura 5.5: Trechos ilustrativos de LFP de animais controle (em preto) registrados no(a)Córtex pré-frontal medial e no(b) hipocampo; e animais VPA (em vermelho) registrados no(c)córtex pré-frontal medial e no(d)hipocampo. Os animais estavam interação social.
Afim de elucidar com mais clareza as diferenças entre as densidades espectrais de potência
entre os dois grupos de animais nos três momentos distintos: em exploração, em repouso e em
interação social; separamos o PSD por ritmos oscilatórios cerebrais: Teta, Beta,Low gamma
e High Gamma (figuras 5.7, 5.8,5.9 e 5.10). As barras correspondem ao valor médio da PSD
dentro da faixa de frequência de cada ritmo. Para comparar os resultados dos grupos, utilizamos
teste-t. Os resultados e comparações que realizaremos, no entanto, servem como caminho para
5.2. Animais em interação social 31
Figura 5.6: PSD do LFP dos animais em interação social. Os gráficos em cinza correspondem ao grupo controle (n=2) e aqueles em vermelho ao grupo VPA (n=2). Sendo ambos da esquerda extraídos do córtex pré-frontal medial, enquanto aqueles da direita vêm do hipocampo.
Conforme já mostrado em diversos estudos, o grupo controle apresenta aumento da potência
do ritmo Teta hipocampal quando em exploração se comparado quando encontra-se em repouso
(p=0,025) (figura 5.7). Nossos resultados mostram também uma diferença dessas medições nos
momentos de exploração e interação social para o grupo controle (p=0,00096). No entanto, estas
alterações não ocorrem no grupo VPA. Por fim, não foram encontradas diferenças entre os grupos
para o ritmo teta seja hipocampal seja cortical.