Abstract— Neural responses are the fundamental expressions of any neural activity. Information carried by a neural response is determined by the nature of a neural activity. In majority of cases the underlying stimulus that triggers it remains largely unknown. Previous studies to reconstruct the stimulus from a neural response show that the high non-linearity of neural dynamics renders inversion of a neuron a challenging task. This paper presents a numerical solution rather than an analytical one to reconstruct stimuli from Hodgkin-Huxley neural responses. The stimulus is reconstructed by first retrieving the maximal conductances of the ionic channels and then solving the Hodgkin-Huxley equations for the stimulus. The results show that the reconstructed stimulus matches the original stimulus to a high degree of accuracy. In addition, this reconstruction approach also retrieves the neural dynamics for which an analytical solution does not currently exist. Constant-current and periodic stimuli are shown to be accurately reconstructed using this approach.
Index Terms—stimulus reconstruction, Hodgkin-Huxley neuron, neural response inverse, neural dynamics retrieval.
I. INTRODUCTION
The relationship between a neural response and its stimulus has been studied over the recent years to understand the encoding and decoding mechanisms adopted by neurons. Not much is known about how neurons specifically encode and decode information. It is thought that either the firing time or the rate of fire of a neuron carries specific neural response information [1-3]. A parallel line of research exists which aims to reconstruct the stimulus from a neural response. A stimulus represents a trigger for a neural activity which underlines any neural response. The ability to reconstruct a stimulus hence offers to retrieve stimulus parameters that can help extend our understanding of neuronal encoding /decoding.
Previous work on input reconstruction has been carried out across many fields like digital filters, neural networks, algorithms and complexity, and digital signal processing [4-13]. Similar approach can be considered for stimulus reconstruction however, due to the high non-linearity of Manuscript received March 18, 2010. Stimulus Reconstruction from a Hodgkin-Huxley Neural Response: A Numerical Solution
Mayur Sarangdhar is currently a PhD student within the Neural, Emergent and Agent Technologies Group, Department of Computer Science, University of Hull, Hull, East-Yorkshire, HU6 7RX, UK (phone: 01482 465253; e-mail: M.Sarangdhar@ 2006.hull.ac.uk).
C. Kambhampati is currently a Reader in the Department of Computer Science, University of Hull, Hull, East-Yorkshire, HU6 7RX, UK. (e-mail: [email protected]).
neural dynamics, it is very difficult to obtain an analytic solution. Periodic signals, unlike aperiodic signals, can be recovered using conventional filters [4]. Artificial neural networks are used to treat the Hodgkin-Huxley (HH) neuron as a black box and reconstruct the stimulus by learning the dynamics [5]. Other implementations use a reverse filter that predicts the sensory input from neuronal activity and recursive algorithms to reconstruct stimulus from an ensemble of neurons [6-7]. The principles of a Time Encoding and Decoding Machines for signal recovery have been explored to reconstruct a neural stimulus whereas, a more direct approach to recover stimulus focuses to make the HH neuron Input-Output (IO) equivalent to an Integrate and Fire (IF) neuron [8-13]. These approaches establish a relationship between the neural response and the stimulus but are not designed to capture or retrieve the neural dynamics. In other words, they offer some starting point for stimulus reconstruction but it is quite a challenge to analytically invert a neuron. However, it is possible to reconstruct stimulus from a neural response using numerical approximations and small time-steps for integration.
This paper aims to reconstruct constant-current and periodic stimuli by a) extracting the maximal conductances from a trace of neural response and b) solving the neural equations for the stimulus. To reconstruct the stimulus, it is imperative that linearization is carried out. This paper demonstrates the above approach using a Hodgkin-Huxley (HH) neuron [14] and Euler integration. The results show that for a small time-step , the accuracy of extracted maximal conductances is very high. Also, the reconstructed stimulus matches the original stimulus accurately. As reconstruction of the stimulus involves solving the neural equations, this approach can replicate the neural dynamics, the time-dependent changes in the voltage-gated ionic channels of Na+, K+ and Cl-. This technique, though computationally demanding, offers a local solution to the problem of inverting a neural response.
II. NEURONAL MODEL AND SYNAPSE
A. The neuron model
The computational model and stimulus for an HH neuron is replicated from [15]. The differential equations of the model are the result of non-linear interactions between the membrane voltage V and the gating variables m, h and n for
, and .
Stimulus Reconstruction from a
Hodgkin-Huxley Neural Response: A Numerical
Solution
i L L K K Na Na I V V g V V n g V V h m g dt dv C ) ( ) ( ) ( 4 3 (1) n n n h h h m m m n dt dn h dt dh m dt dm ) ( ) ( ) ( (2) 80 / ) 65 ( 10 / ) 35 ( 18 / ) 65 ( 10 / ) 55 ( 20 / ) 65 ( 10 / ) 40 ( 125 . 0 ] 1 /[ 1 4 ] 1 /[ ) 55 ( 01 . 0 07 . 0 ] 1 /[ ) 40 ( 1 . 0 V n V h V m V n V h V m e e e e V e e V (3)
The variable V is the resting potential of the membrane and
Na
V , VKand VLare the reversal potentials of theNa , K
channels and leakage. The values of the reversal potentials
. 5 . 54 , 77 ,
50mVV mVV mV
VNa K L The conductance
for the ionic channels are 2
/
120m S cm
gNa , gK 36m S/cm2 andgL 0.3m S/cm2. The capacitance of the membrane is
2
/ 1 F cm
C .
B. The synaptic current
An input spike train give by [16] is considered to generate the pulse component of the external current.
n f a
i t V t t
U() ( ) (4)
where, tfis the firing time and is defined as
T
t
t
n f nf( 1) ( ) (5) 0
) 1 (
f
t (6)
T represents the ISI of the input spike train and can be varied to generate a different pulse current. The spike train is injected through a synapse to give the pulse currentIP.
) (
)
( a syn
n
f syn
P g t t V V
I (7)
syn syn V
g , are the conductance and reversal potential of the synapse. [32] define the function as
), ( ) / ( )
(t t e t/ t (8)
where, is the time constant of the synapse and (t)is the Heaviside step function. Va 30mV, syn 2m s ,
2 / 5 .
0 mScm
gsyn andVsyn 50mV.
C. The total external current
The total external current applied to the neuron is a combination of static and pulse component
P S
i I I
I (9)
where, ISis the static and Ipis the pulse current, is the
random Gaussian noise with zero mean and standard deviation 0.025.
On injection of a periodic or sinusoidal stimulus the steady state response of a neuron is no longer preserved [17-25]. The self-excited oscillations of the HH neuron [14] may become chaotic when a sinusoidal stimulus is applied with proper choices of magnitude and frequency [20-21, 25-26]. Physiological experiments on squid giant axons [18-19] and Onchidium neurons [22] have confirmed the occurrence of chaotic oscillations. It is understood that distinct sinusoidal stimuli induce different chaotic oscillations which result in dissimilar neural responses [27-29].
III. STIMULUS RECONSTRUCTION
Let be the neural response of the HH neuron to a synaptic stimulus and ionic conductances , and . Assuming that is unknown and only the neural response and the reversal potentials are known, the aim is to reconstruct the stimulus such that and are identical. Therefore the target is to retrieve , and and get without any information of .
A. Extracting Maximal Conductances
Equations (1-3) show that the gating variables , and only depend on the instantaneous voltage at time . The instantaneous voltage at time is given by
t L L K t K t Na Na t dt V t v g dt V t v t n g dt V t v t h t m g dt t I C v t v 0 0 4 0 3 0 ' ) ) ' ( ( ' ) ) ' ( ( ) ' ( ' ' ) ) ' ( ( ) ' ( ) ' ( ' ' ) ' ( 1 ) 0 ( ) ( (10) To retrieve the three ionic conductances, linear equations in three unknowns need to be solved. The formulation of the equations is proposed as an algorithm in [30]. Given a small voltage trace , select three times , As the voltage trace is known over all , is known for
.
Let functions fj(t),j 1,2,3be defined as
' ) ) ' ( ( 1 ) ( ' ) ) ' ( ( ) ' ( 1 ) ( ' ) ) ' ( ( ) ' ( ) ' ( 1 ) ( 0 3 0 4 2 0 3 1 dt V t v C t f dt V t v t n C t f dt V t v t h t m C t f t L t K t Na (11)
and defined as
t
dt
t
I
v
t
v
t
b
0'
)
'
(
)
0
(
)
(
)
(
(12)Hence, ) ( ' ) ( ' ) ( ' )
(t g f1t g f2 t g f3 t
b Na K L (13)
If t
dt
t
I
0'
)
'
(
is a known analytic function, the value of)
(
t
variables, , and are obtained by integrating the HH equations. If , and are the gating-variables’ estimates and is the resultant approximation of , then the retrieving maximal conductances can be defined as a solution to the linear system
N
i
x
t
f
t
b
j j i ji
)
(
)
,
1
...
(
3
1 '
(14) This is an overdetermined system of linear equations in the form . An approximate solution can be obtained by using the full set of data generated during the integration of the HH equations and treating (14) as a linear least squares problem.
Hence, the best fit solution in the linear least squares sense is obtained by solving
2 1 3 1
)
(
'
)
(
min
N i j j i j ix
b
t
f
t
x
(15)If is the matrix whose entries are and ,
2
min
A
x
b
x (16)
As the equations are linear in , a solution is obtainable.
B. Reconstructing the stimulus
The approach defined above requires the knowledge of both the voltage and the external stimulus , for all time . In principle, it is unrealistic to know the stimulus for all times and in majority cases, the stimulus remains unknown. Therefore, retrieving the maximal conductances using the equations (11-16) is specific when all parameters are known.
However, it is possible to reconstruct the stimulus entirely without the knowledge of corresponding for a neural response . As the type of the neuron and the reversal potential for Na+, K+ and Cl- is known, we propose that the neural stimulus can be reconstructed without the knowledge of the original stimulus .
1. Record any neural response whose stimulus, say , requires to be reconstructed
2. Inject a supra-threshold stimulus, for a small time duration
3. Record the corresponding voltage trace generated,
4. Retrieve the maximal conductances using equations (11-16) and as the external stimulus
5. Using the approximated maximal conductances, , and , solve the HH equations using the recorded neural response and the stimulus as the only unknown to get the reconstructed stimulus
The HH equations can be re-written as
dt dv C V t v g V t v t n g V t v t h t m g t I L L K K Na Na ) ) ( ( ' ) ) ( ( ) ( ' ' ) ) ( )( ( ' ) ( ' ' ) (
' 3 4
(17)
where, , and are the approximated maximal
conductances calculated from and , and are the estimates of the gating variables , and respectively. As is known for all times , the rate of change of voltage ( ) can be numerically approximated.
' ' ' ' ' ' ' ' ' ' ) ( ' ' ) ( ' ' ) ( ' n n n h h h m m m n dt dn h dt dh m dt dm (18) 80 / ) 65 ( ' 10 / ) 35 ( ' 18 / ) 65 ( ' 10 / ) 55 ( ' 20 / ) 65 ( ' 10 / ) 40 ( ' 125 . 0 ] 1 /[ 1 4 ] 1 /[ ) 55 ( 01 . 0 07 . 0 ] 1 /[ ) 40 ( 1 . 0 V n V h V m V n V h V m e e e e V e e V (19)
This approach provides a local solution to reconstructing the neural stimulus of a HH neuron and also approximates the gating variables. In addition to the retrieval of stimulus parameters, it also estimates the neural dynamics which are important represent the open-close mechanism of ionic gates.
IV. COMPUTATIONAL RESULTS
A. Generating a Voltage Trace
Let be a small supra-threshold step current that evokes an action potential. The resultant voltage trace is sufficient to retrieve the maximal conductance values.
Fig.1: The voltage trace generated by a small step-current . This small trace of neural voltage is sufficient to retrieve the maximal conductances.
B. Retrieving Maximal Conductances
linear equations, an exact solution cannot be obtained for all values of .
Original /Retrieved
Table 1: Retrieved maximal conductance values for various values of . The conducances are highly accurate as becomes close to 0.
The relative error of the approximations decreases as becomes close to 0.
Relative error ( )
0.01 0.0037
0.001 0.00038
0.0001 0
Table 2: The relative error decreases as becomes close to 0.
The voltage traces reconstructed from the approximated conductances are shown in fig. 2. The estimated maximal conductance values produce a good fit to the original trace .
Fig.2: The reconstructed voltage trace using the approximated maximal conductance values for different time-steps . As becomes close to 0, the approximations approach the actual conductance values. For , the approximated conductance values are equal to the original values. Hence the trace generated by overlaps with the original trace .
C. Stimulus Reconstruction
The retrieval of maximal conductance values such that a good fit of the original voltage trace is produced indicates that the approximations are nearly accurate. Using equations (17-19), a linearised reconstruction of a stimulus can be obtained.
1) Constant-Current Stimulus
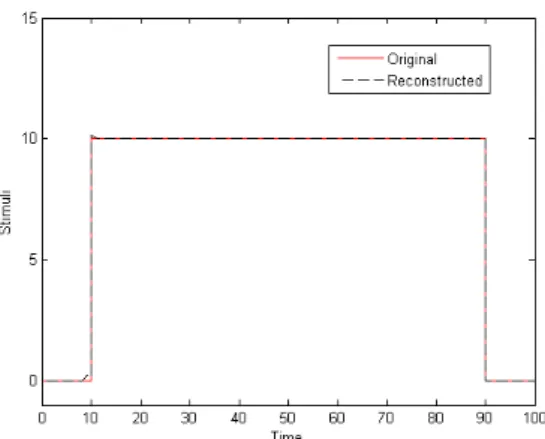
Let the HH neuron be stimulated by an unknown step-current such that it evokes a series of action potentials . The maximal conductances are approximated in Table 1. The reconstructed stimulus is shown in fig. 3.
Fig.3: The reconstructed stimulus is good fit to the original stimulus. The original stimulus is very well approximated if chosen is close to 0.
Fig.4: The approximations become less accurate with an increase in . Results show that if the time-step of Euler integration is sufficiently small i.e. , the maximal conductances can be accurately retrieved. The stimulus reconstructed using these maximal conductance values, is a near approximation of the original unknown stimulus.
2) Periodic Stimulus
If the HH neuron is stimulated by an unknown periodic stimulus , the resultant neural response is . Generating a trace voltage to retrieve the maximal conductance values, the unknown stimulus can be reconstructed using (17-19).
Fig.6: The approximation of the reconstructed stimulus become less accurate with an increase in ( ). The numerical approximation of the derivatives causes some jitters.
Fig. 7: Due to the presence of noise in original stimulus, the reconstructed stimulus cannot be an exact match. The jitters are due to the numerical approximation to the rate of change of voltage. However, the reconstruction is very close to the original stimulus for close to 0.
It is observed that the unknown stimulus can be predicted accurately if is small and close to 0. As a result, the computational time required by this approach is directly proportional to the choice of . However, this is approach provides a local solution to reconstructing unknown stimuli using the knowledge of the computational model of a neuron. It is also possible to retrieve the neural dynamics which cannot be retrieved by a purely analytical approach (fig. 8).
Fig.8: The reconstructed neural dynamics. This numerical solution can retrieve the gating variables and their time constants
.
V. CONCLUSIONS
The neural dynamics of the HH neuron have been the subject of research for many years now. The dynamics put
forth by Hodgkin and Huxley have been well studied and replicated by many researchers. In much the same way, inverting the HH neural equations has attracted interest in recent years. The equations of the HH neuron are highly non-linear due to the incorporation of probability of the gating variables which regulate the open-close mechanism of ionic channels.
Previous research has addressed the problem of inverting this non-linear neuron by using digital filters, neural networks, algorithms and complexity, and digital signal processing. Other approaches point to the use of reconstruction algorithms, time encoding/decoding machines or an IF neuron. These approaches establish a relationship between the neural response and the stimulus but they are not designed to capture or retrieve the neural dynamics.
The approach described in this paper provides a numerical solution to reconstruct an unknown neural stimulus. An unknown stimulus can be numerically reconstructed by
1. Recording any neural response whose stimulus, say , requires to be reconstructed 2. Injecting a supra-threshold stimulus, for a
small time duration
3. Recording the corresponding voltage trace generated,
4. Retrieving the maximal conductances using equations (11-16) and as the external stimulus
5. Using the approximated maximal conductances, , and , solve the HH equations using the recorded neural response and the stimulus as the only unknown to get the reconstructed stimulus
It is observed that the accuracy of maximal conductances retrieved by solving an overdetermined system of linear equations depends on the time-step ( ) of Euler integration. A small value of can reproduce almost exact maximal conductances. Accurate maximal conductance values help reconstruct a near-fit approximation of the original stimulus. Due to the nature of numerical approximation and the inherent non-linearity in the neural dynamics, the reconstructed stimulus shows some jitters. Also, it is noticed that if the original stimulus carries any noise, an exact match of the stimulus cannot be reconstructed. However, the reconstructed stimulus still matches the original stimulus to a high degree of accuracy. The choice of is very important and there is a trade-off between computational time and accuracy. The accuracy increases with a decrease in .
REFERENCES
[1] Rinzel J (1985). Excitation dynamics: insights from simplified membrane models. Theoretical Trends in Neuroscience Federal Proceedings, Vol. 44, No. 15, 2944-2946.
[2] Panzeri S, Schultz SR, Treves A, Rolls ET (1999). Correlations and the encoding of information in the nervous system. Proc. R. Soc. Lond. B 266, 1001-1012.
[3] Gabbiani F, Metzner W (1999). Encoding and Processing of Sensory Information in Neuronal Spike Trains. The Journal of Biology, 202, 1267-1279.
[4] Das A, Folland R, Stocks NG, Hines EL (2006). Stimulus reconstruction from neural spike trains: Are conventional filters suitable for both periodic and aperiodic stimuli? Signal Processing, 86 (7), 1720-1727.
[5] Saggar M, Mericli T, Andoni S, Miikkulainen R (2007). System identification for the Hodgkin-Huxley model using artificial neural networks. IEEE International Joint Conference on Neural Networks Orlando, FL, AUG 12-17, 2239-2244.
[6] Stanley GB, SeyedBoloori A (2001). Decoding in neural systems: Stimulus reconstruction from nonlinear encoding. Proceedings of the 23rd Annula International Conference of the IEEE Engineering in Medicine and Biology Society, Vols 1-4, Vol. 23, 816-819.
[7] Stanley GB (2001). Recursive stimulus reconstruction algorithms for real-time implementation in neural ensembles. Neurocomputing, 38, 1703-1708.
[8] Lazar AA, Pnevmatikakis EA (2009). Reconstruction of Sensory Stimuli Encoded with Integrate-and-Fire Neurons with Random Thresholds. EURASIP Journal on advances in Signal Processing. Article no. 682930.
[9] Lazar AA (2007). Information representation with an ensemble of Hodgkin-Huxley neurons. Neurocomputing, vol. 70 (10-12), 1764-1771.
[10] Lazar AA (2007). Recovery of Stimuli Encoded with Hodgkin-Huxley Neurons, Computational and Systems Neuroscience Meeting, COSYNE 2007, Salt Lake City, UT, February 22-25.
[11] Lazar, AA (2006). Time encoding machines with multiplicative coupling, feedforward, and feedback. IEEE Transactions on Circuits and Systems II – Express Briefs. Vol. 53 (8), 672-676. [12] Lazar AA (2004). Time encoding with an integrate-and-fire
neuron with a refractory period. Neurocomputing, Vol.58, 53-58 [13] Lazar AA, Simonyi EK, Toth LT(2006). A Real-Time Algorithm for Time Decoding Machines. 14th European Signal Processing Conference, September, 2006.
[14] Hodgkin A, Huxley A (1952). A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 117:500–544.
[15] Hasegawa H (2000). Responses of a Hodgkin-Huxley neuron to various types of spike-train inputs. Physical Review E, Vol. 61, No. 1.
[16] Park MH, Kim S (1996). Analysis of Phase Models for two Coupled Hodgkin-Huxley Neurons. Journal of the Korean Physical Society, Vol. 29, No. 1, pp. 9-16.
[17] Wang XJ, Buzsáki G (1996). Gamma Oscillation by Synaptic Inhibition in a Hippocampal Interneuronal Network Model. The The Journal of Neuroscience, 16(2)
[18] Guttman R, Feldman L, Jakobsson E (1980). Frequency Entrainment of Squid Axon Membrane. Journal of Membrane Biology, 56, 9-18.
[19] Matsumoto G, Kim K, Ueda T, Shimada J (1980). Electrical and Computer Simulations upon the Nervous Activities of Squid Giant Axons at and around the State of Spontaneous Repetitive Firing of Action Potentials. J. Phys. Soc. Jpn., 49, 906. [20] Aihara K, Matsumoto G, Ikegaya Y (1984). Periodic and
Non-periodic Responses of a Periodically Forced Hodgkin-Huxley Oscillator. Journal of Theoretica Biology, 109, 249-269.
[21] Matsumoto G, Aihara K, Ichikawa M, Tasaki A (1984). Periodic and Nonperiodic responses of membrane potentials in squid giant axons during sinusoidal current simulations. Journal of Theoretical Neurobiology, 3, 1-14.
[22] Hayashi h, Ishizuka S, Hirakawa K (1985). Chaotic responses of the pacemaker neuron. J. Phys. Soc. Jpn., 54, 2337.
[23] Holden AV (1987). Chaos. Manchester: Manchester University Press.
[24] Hasegawa H (2008). Information conveyed by neuron populations – Firing rate, fluctuations and synchrony. Neuroquantology, Vol. 6 (2), 105-118.
[25] Kaplan D, Glass L (1995). Understanding Non Linear Dynamics. Springer, New York.
[26] Wilson HR (1999). Spikes, Decisions and Actions – The dynamical foundations of neuroscience. Oxford University Press Inc., New York.
[27] Sarangdhar M, Kambhampati C (2008a). Spiking Neurons: Is coincidence-factor enough to compare responses with fluctuating membrane voltage? In World Congress on Engineering 2008 : The 2008 International Conference of Systems Biology and Bioengineering, London, U.K. 2-4 July 2008, Vol. 2, pp 1640-1645.
[28] Sarangdhar M, Kambhampati C (2008b). Spiking Neurons and Synaptic Stimuli: Determining the Fidelity of Coincidence-Factor in Neural Response Comparison. Engineering Letters, Volume 16 Issue 4, pp 512-517.
[29] Sarangdhar M, Kambhampati C (2009). Spiking Neurons and Synaptic Stimuli - Neural Response Comparison using Coincidence-Factor. In Advances in Electrical Engineering and Computational Science. Edited by Gelman L, Balkan N, Ao S. Published by Springer.
[30] Shepardson D (2009). Algorithms for inverting Hodgkin-Huxley type neuron models. PhD diss., Georgia Institute of Technology.
http://www.aco.gatech.edu/doc/Shepardson_thesis.pdf, accessed