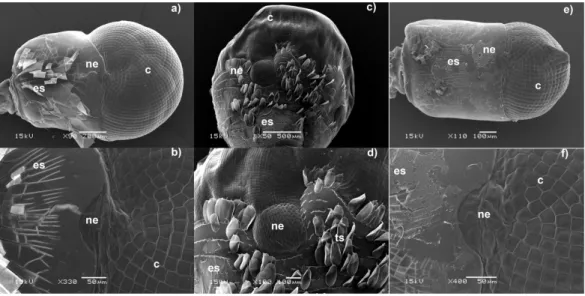
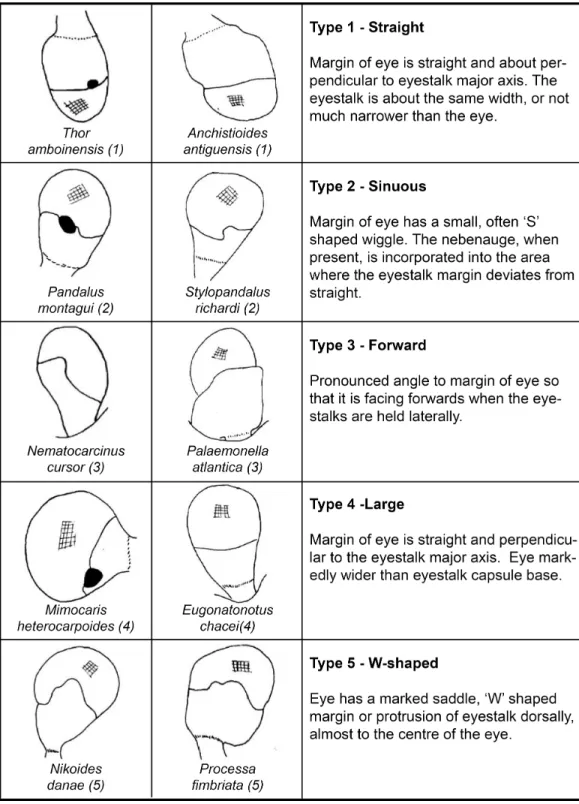
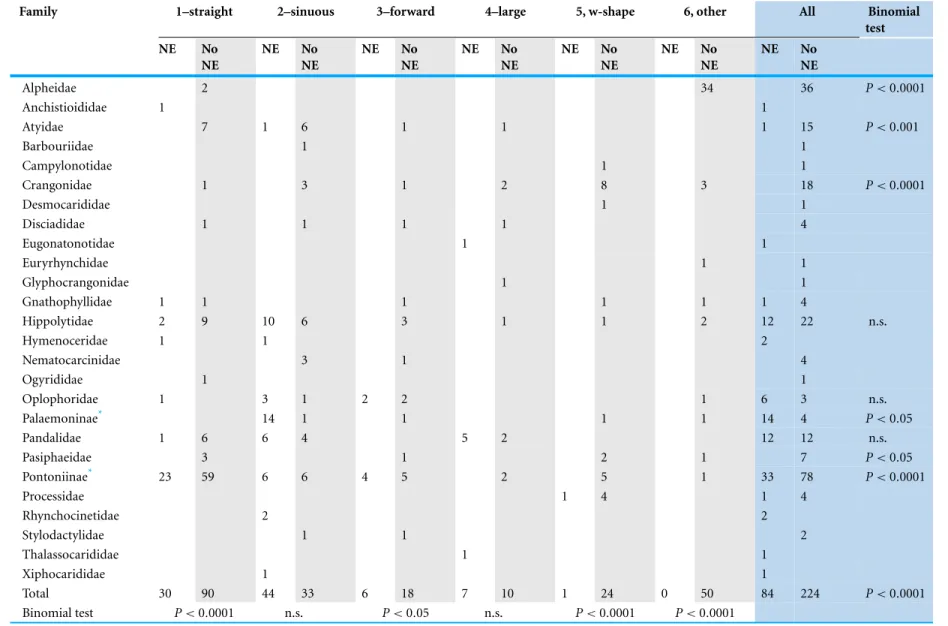
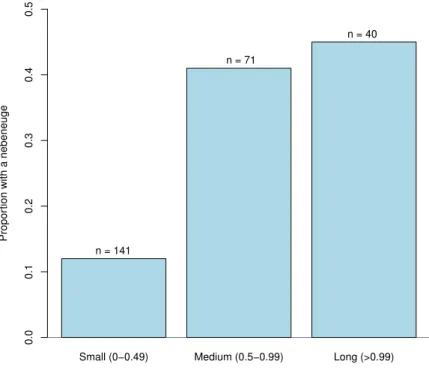
External morphology of eyes and Nebenaugen of caridean decapods–ecological and systematic considerations
Texto
Imagem
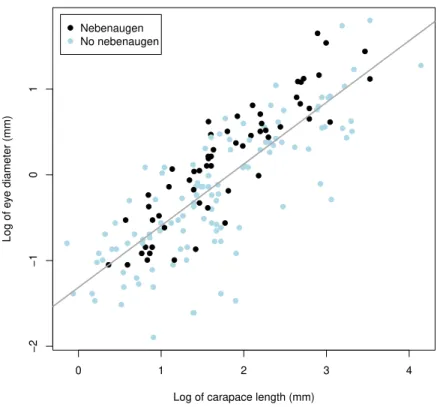
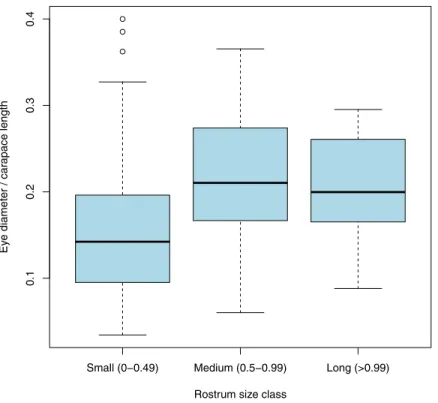
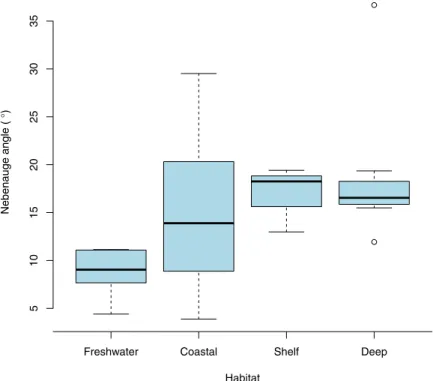
Documentos relacionados
Este modelo permite avaliar a empresa como um todo e não apenas o seu valor para o acionista, considerando todos os cash flows gerados pela empresa e procedendo ao seu desconto,
Em sua pesquisa sobre a história da imprensa social no Brasil, por exemplo, apesar de deixar claro que “sua investigação está distante de ser um trabalho completo”, ele
Despercebido: não visto, não notado, não observado, ignorado.. Não me passou despercebido
Caso utilizado em neonato (recém-nascido), deverá ser utilizado para reconstituição do produto apenas água para injeção e o frasco do diluente não deve ser
This log must identify the roles of any sub-investigator and the person(s) who will be delegated other study- related tasks; such as CRF/EDC entry. Any changes to
Além disso, o Facebook também disponibiliza várias ferramentas exclusivas como a criação de eventos, de publici- dade, fornece aos seus utilizadores milhares de jogos que podem
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
É nesta mudança, abruptamente solicitada e muitas das vezes legislada, que nos vão impondo, neste contexto de sociedades sem emprego; a ordem para a flexibilização como
