UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
AVALIA
Ç
ÃO DE HÍBRIDOS SIMPLES BRAQUÍTICOS DE MILHO SUPER DOCE
(
Zea mays
L.) PORTADORES DO GENE SHRUNKEN-2 (
sh
2sh
2) UTILIZANDO O
ESQUEMA DIALÉLICO PARCIAL
CARLOS ALBERTO ARAGÃO
Tese apresentada à Faculdade de
Ciências Agronômicas UNESP - Campus
de Botucatu, para a obtenção do título
de Doutor em Agronomia - Área de
Concentração em Agricultura
.
UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
AVALIA
Ç
ÃO DE HÍBRIDOS SIMPLES BRAQUÍTICOS DE MILHO SUPER DOCE
(
Zea mays
L.) PORTADORES DO GENE SHRUNKEN-2 (
sh
2sh
2) UTILIZANDO O
ESQUEMA DIALÉLICO PARCIAL
CARLOS ALBERTO ARAGÃO
Orientador: Prof. Dr. NORBERTO DA SILVA
Tese apresentada à Faculdade de
Ciências Agronômicas UNESP - Campus
de Botucatu, para a obtenção do título
de Doutor em Agronomia - Área de
Concentração em Agricultura
.
BIOGRAFIA DO AUTOR
Carlos Alberto Aragão, filho de João Francisco Aragão e Maria Valdenice Aragão,
nasceu na cidade de Vitória da Conquista, Estado da Bahia, a 04 de Setembro de 1970.
Diplomou-se em Agronomia pela Universidade Estadual do Sudoeste da Bahia
(UESB), no ano de 1996.
Em setembro de 1996, iniciou o curso de Mestrado em Agronomia, Área de
Concentração Genética e Melhoramento de Plantas, no Departamento de Biologia da
Universidade Federal de Lavras (UFLA), concluindo em setembro de 1998.
DEDICATÓRIA
Aos que me ensinaram os primeiros passos,
que são os responsáveis pela minha existência,
pela existência dessa conquista,
que sempre acreditaram em mim
e em momento algum, deixaram de me apoiar
DEDICO:
Meus pais, João Francisco Aragão “in memorian”
Maria Valdenice Aragão
Gratidão eterna.
Irmãos, Salete Aparecida Aragão, José Valdemir Aragão, João Francisco Filho Aragão,
Márcio Aragão, Ana Lúcia Aragão e minha esposa Bárbara França Dantas, companhia de
todos os momentos.
Meus agradecimentos.
A meu pai
João Francisco Aragão "in memorian", a minha
AGRADECIMENTOS
- À Faculdade de Ciências Agronômicas/UNESP, Campus de Botucatu pela
oportunidade da realização do Curso.
- À Coordenadoria de Apoio e Pesquisa ao Ensino Superior (CAPES) pela concessão
da Bolsa de estudos para o meu Curso de Doutorado.
- Ao Professor Dr. Norberto da Silva pela orientação e apoio.
- Aos Professores do Curso de Pós-graduação Maurício Dutra Zanotto, Sílvio José
Bicudo, João Nakagawa, Cláudio Cavariani e Rumy Goto pelos ensinamentos e
convívio.
- Aos Funcionários da Fazenda Experimental São Manuel, da SAKATA/Sudameris, e
do Departamento de Agricultura pelo apoio durante a execução dos experimentos e
coletas dos dados.
- Ao Engenheiro Agrônomo Wanderlei Barbosa, pelo apoio no experimento instalado
em Piracanjuba-GO.
- Aos Professores Maurício Dutra Zanotto, Sílvio José Bicudo, Celso Luis Marino e
Cláudio Lopes de Souza Júnior, pelas valiosas sugestões na Defesa da Tese.
- Aos grandes amigos Marcelo, Elza, Rodrigo, Luca, Eduardo, Patrícia, Drolle,
Recco, Morais, Marcinha, Edwin, Munir, Juliano, Moizés, Edmilson, Caetano,
companheiros de todos as horas.
- Aos meus familiares pelo apoio, carinho e compreensão em todos os instantes.
- A minha querida esposa Bárbara por todo amor compartilhado.
SUMÁRIO
LISTA DE QUADROS E TABELAS...VII
LISTA DE ABREVIATURAS...IX
1 RESUMO...1
2 SUMMARY...3
3 INTRODUÇÃO...5
4 REVISÃO DE LITERATURA...8
4.1 Particularidades do milho doce...8
4.2 A plantas do milho doce...10
4.3 Mutantes do milho doce...10
4.4 Utilização de híbridos...14
4.4.1 Heterose (vigor do híbrido)...14
4.4.2 Análise dialélica...17
5 MATERIAL E MÉTODOS...21
5.1 Material...21
5.2 Obtenção do material experimental...21
5.3 Localização dos experimentos...21
5.4 Épocas de semeadura...24
5.5 Delineamento experimental...25
5.6 Caracteres avaliados...25
5.7 Análises estatística e genética...29
5.7.1 Análise estatística...29
6 RESULTADOS E DISCUSSÃO...31
6.1 Resumo das análises de variância...31
6.1.1 Análises individuais...31
6.1.2 Análises dos três locais...32
6.2 Médias dos diferentes caracteres avaliados nos híbridos, comparadas pelo teste
Tukey, nos três locais...42
6.3 Análise da capacidade combinatória...77
6.3.1 Estimativas médias da capacidade geral de combinação (CGC)...81
6.3.2 Estimativas médias da capacidade específica de combinação (CEC)...84
7 CONCLUSÕES.... ...96
LISTA DE QUADROS E TABELAS
1. QUADRO 1 Composição química de cultivares de milho...9
2. TABELA 1 Quadrados médios obtidos da análise de variância de seis caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 1 (São Manuel – SP)...34
3. TABELA 2 Quadrados médios obtidos da análise de variância de cinco caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 1 (São Manuel - SP)...35
4. TABELA 3 Quadrados médios obtidos da análise de variância de seis caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 2 (SAKATA Sudameris - SP)...36
5. TABELA 4 Quadrados médios obtidos da análise de variância de seis caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 2 (SAKATA Sudameris - SP)...37
6. TABELA 5 Quadrados médios obtidos da análise de variância de seis caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 3 (Arisco - GO)...38
7. TABELA 6 Quadrados médios obtidos da análise de variância de seis caracteres
avaliados em híbridos simples de milho super doce e respectivas significâncias pelo
teste F, obtidos no Local 3 (Arisco - GO)...39
8. TABELA 7 Quadrados médios obtidos da análise de variância conjunta de seis
caracteres avaliados em híbridos simples de milho super doce e respectivas
significâncias pelo teste F, obtidos nos três locais de instalação dos experimentos...40
9. TABELA 8 Quadrados médios obtidos da análise de variância conjunta de seis
caracteres avaliados em híbridos simples de milho super doce e respectivas
significâncias pelo teste F, obtidos nos três locais de instalação dos experimentos...41
10. TABELA 9 Médias da produção total (PTCst/kg) corrigida para estande de híbridos de
milho super doce em três locais de plantio...43
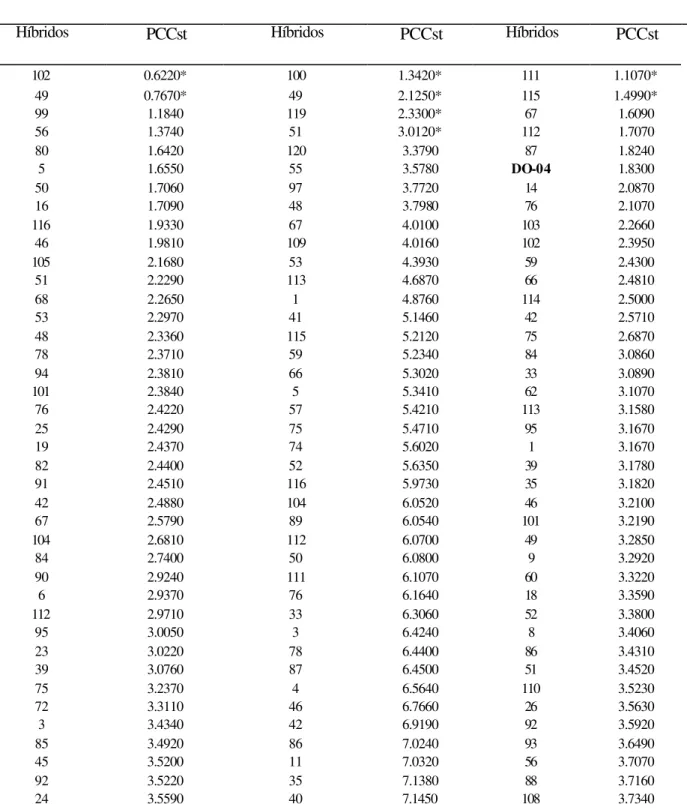
11. TABELA 10 Médias da produção comercial com palha (PCCst/kg) corrigida para
estande de híbridos de milho super doce em três locais de plantio...47
12. TABELA 11
Médias do peso de 5 espigas comerciais com palha (P5Eccp/kg) de
13. TABELA 12 Médias do peso de 5 espigas comerciais com palha (P5Eccp/kg) de
híbridos de milho super doce em três locais de plantio...54
14. TABELA 13
Médias da produção de espigas comerciais despalhadas corrigida para
estande e 76% de umidade (PC5Esp-76%/kg) de híbridos de milho super doce em três
locais de plantio...58
15. TABELA 14
Médias do comprimento de espigas (CoE/cm) de híbridos de milho super
doce em três locais de plantio...63
16. TABELA 15
Médias do diâmetro de espigas (
φ
E/mm) de híbridos de milho super doce
em três locais de plantio...66
17. TABELA 16 Médias da altura de plantas (AlP/m) de híbridos de milho super doce em
três locais de plantio...71
18. TABELA 17 Médias da altura de inserção (AlE/m) espiga de híbridos de milho super
doce em três locais de plantio...74
19. TABELA 18 Quadrados médios das capacidades combinatórias (Geral e Específica) e
respectivas significâncias pelo teste F, obtidos da análise conjunta de cinco caracteres
avaliados em híbridos simples de milho super doce nos três locais de instalação dos
experimentos...79
20. TABELA 19 Quadrados médios das capacidades combinatórias (Geral e Específica) e
respectivas significâncias pelo teste F, obtidos da análise conjunta de cinco caracteres
avaliados em híbridos simples de milho super doce nos três locais de instalação dos
experimentos...80
21. TABELA 20 Estimativas dos efeitos da Capacidade geral de combinação (CGC),
obtidos da análise conjunta de cinco caracteres avaliados em híbridos simples de milho
super doce nos três locais de instalação dos experimentos...82
22. TABELA 21 Estimativas dos efeitos da Capacidade geral de combinação (CGC),
obtidos da análise conjunta de cinco caracteres avaliados em híbridos simples de milho
super doce nos três locais de instalação dos experimentos...83
23. TABELA 22
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para produção total corrigida para estande, obtidos da análise conjunta avaliados em
híbridos simples de milho super doce nos três locais de instalação dos
experimentos...85
24. TABELA 23
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
25. TABELA 24
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para peso de 5 espigas comerciais com palha, obtidos da análise conjunta avaliados em
híbridos simples de milho super doce nos três locais de instalação dos
experimentos...88
26. TABELA 25
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para peso de 5 espigas comerciais sem palha, obtidos da análise conjunta avaliados em
híbridos simples de milho super doce nos três locais de instalação dos
experimentos...89
27. TABELA 26
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para produção de espigas comerciais sem palha corrigida para estande e 76% de
umidade, obtidos da análise conjunta avaliados em híbridos simples de milho super
doce nos três locais de instalação dos experimentos...90
28. TABELA 27 Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para comprimento de espigas, obtidos da análise conjunta avaliados em híbridos
simples de milho super doce nos três locais de instalação dos experimentos...91
29. TABELA 28
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para diâmetro de espigas, obtidos da análise conjunta avaliados em híbridos simples de
milho super doce nos três locais de instalação dos experimentos...92
30. TABELA 29
Estimativas dos efeitos da Capacidade específica de combinação (CEC)
para altura de plantas, obtidos da análise conjunta avaliados em híbridos simples de
milho super doce nos três locais de instalação dos experimentos...94
31. TABELA 30 Estimativas dos efeitos da Capacidade específica de combinação (CEC)
LISTA DE ABREVIATURAS
1 RESUMO
EVALUATION OF BRAQUÍTIC SIMPLE HIBRIDS OF SUPER SWEET CORN (
Zea
mays
L.) WITH SHRUNKEN-2 (
sh
2sh
2) GENE, USING THE PARTIAL DIALLEL
SCHEME. Botucatu, 2002. 101p. Tese (Doutorado em Agronomia/Agricultura) -
Faculdade de Ciências Agronômicas, Universidade Estadual Paulista.
Author: CARLOS ALBERTO ARAGÃO
Adviser: NORBERTO DA SILVA
2 SUMMARY
(
44
)421 x 320 and (
68
)657 x 320, which have great potential for sweet corn breeding
programs
3 INTRODU
Ç
ÃO
O milho doce é classificado como especial e destina-se
exclusivamente ao consumo humano. Na fase de grãos leitosos, isto é, milho verde, os
grãos são tenros e apresentam maior quantidade de sacarose, dextrinas e vitaminas em
relação ao milho verde comum (Storck & Lovato, 1991). Esse tipo de milho vem sendo
consumido em diversos países na forma de espiga cozida “in natura” ou congelada ou como
grãos enlatados. No Brasil várias agroindústrias fomentam a produção e comercializam o
milho doce enlatado, conserva. Estima-se atualmente uma área cultivada anual em torno de
12.000 ha, com uma produção de, aproximadamente, 84.000 t de espigas verdes.
Metropolitana, próximas à cidade do Rio de Janeiro, grande centro consumidor do produto.
Nos últimos anos, o consumo vem aumentado e a oferta do produto mantido constante,
dando margem à importação de outros Estados (Oliveira et al., 1990).
No Brasil, algumas empresas governamentais e particulares
vêm desenvolvendo programas de melhoramento genético para produção de cultivares
adaptados as nossas condições. O Centro Nacional de Pesquisa de Hortaliça (CNPH),
juntamente com o Centro Nacional de Pesquisa de Milho e Sorgo (CNPMS),
desenvolveram e lançaram em 1984, três cultivares: a ‘Superdoce’, a ‘Doce ouro’ e a
‘Doce-cristal’, e dois híbridos simples, ‘Lili’ e ‘Doce mel’ (Reifeschneider et al., 1984 e
1988). Com isso, estabeleceu um programa de sementes básicas deste material para atender
à demanda das companhias de sementes, das agroindústrias e dos demais usuários. No
entanto esses materiais apresentam algumas características indesejáveis, por serem de
espigas curtas e ciclo precoce, havendo necessidade então de obtenção de novos cultivares
com adaptação a diversas regiões do país e com boas características comercias, além de que
o número de cultivares recomendado para o nosso mercado, a época, ser ainda pequeno em
relação à crescente demanda (Parentoni et al., 1990).
Em face da demanda por cultivares de milho doce que reúnam
as características desejadas pelos produtores, industrias enlatadoras e consumidores, a
Faculdade de Ciências Agronômicas – FCA/UNESP - Botucatu, através de convênio com a
Indústria de Conservas Alimentícias (CICA), iniciou um programa de melhoramento de
milho doce a partir de 1989, selecionando materiais, com base no gene
shrunken-2
por
combinação de linhagens avançadas (S4), para obtenção de híbridos com bom potencial
produtivo.
4 REVISÃO DE LITERATURA
4.1 Particularidades do milho doce
O milho doce deve possuir características próprias: sabor
adocicado, pericarpo fino e endosperma com textura delicada. É um produto de alto valor
nutricional, como pode ser visualizado no Quadro1 (Pereira, 1987).
QUADRO1- Composição química de cultivares de milho doce e milho comum
Super-doce
(BR 400)
Doce-de-ouro
(BR 401)
Doce-cristal
(BR 402)
Milho comum
Sólidos solúveis (Brix)
Cinzas (%)
Proteínas
Lipídeos (%)
Amido (%)
Açúcares redutores
Açúcares totais
20.8
0.8
11.6
3.7
20.2
2.1
5.2
22.3
0.9
11.0
4.4
22.9
1.9
4.6
17.7
1.1
10.8
4.5
24.2
1.6
4.3
13.6
1.4
10.3
4.8
71.5
0.55
2.0
Adaptado de Pereira,1987.
4.2 A planta do milho doce
O milho doce (
Zea mays
L. grupo saccharata) pertencente à família
gramínea, tribu Maydeae, do gênero Zea, é uma espécie de grande importância econômica e
social. Originário da América, provavelmente da região onde se situa o México, foi
domesticado num período entre 7.000-10.000 anos atrás. O homem herdou dos povos mais
antigos, cerca de 300 raças de milho, caracterizadas pelas mais diversas adaptações, tanto
para condições climáticas, como para vários usos do cereal. Apresentando também uma
grande quantidade de genes identificados, o que torna à espécie botânica de maior
diversidade genética existente na natureza (Bull & Cantarella, 1993).
A botânica e reprodução no milho doce são idênticas às do
milho comum. Diferencia-se apenas, no grão, que após a maturação fisiológica seca
vagarosamente e, uma vez seco, é vítreo e enrugado. Vítreo por causa da cristalização dos
açúcares que se encontram em maior concentração e enrugado devido à menor proporção
de amido no endosperma (Stork & Lovato, 1991).
4.3 Mutantes de milho doce
O caráter doce desse tipo de milho deve-se a presença
desses genes mutantes que acarretam uma mudança no metabolismo vegetal, que consiste
no bloqueio da conversão de açúcares em amido. A expressão de todos esses genes ocorre
no endosperma do grão (Springer et al., 1986, Laughnan, 1953).
Quando se dá o processo de secagem dos grãos de milho
doce ocorre a cristalização de açúcares, onde os grãos enrugam e o endosperma torna-se
vítreo (Tosello, 1978), enquanto que, em grãos maduros normais o grão é liso e amiláceo
(Zuber, 1965).
Parentoni et al. (1990), sugeriu uma classificação para milho
doce em dois grupos básicos:
- Milho doce: contendo o gene sugary
- Milho super doce: contendo o gene brittle, shrunken ou sugary enhancer
Os mutantes de milho doce em triplas combinações, como
aeduwx, não fizeram sucesso no mercado como os mutantes simples. Nos EUA, apenas um
híbrido triplo tem sido comercializado. Além disso, esses triplos mutantes não apresentam
aumento do teor de açúcar suficiente para justificar seu estabelecimento no comércio
(Kaukis & Davis, 1986).
A concentração de carboidratos dos grãos sugary é
praticamente a mesma dos grãos normais, isso ocorre porque o aumento contínuo do teor de
fitoglicogênio leva ao aumento da concentração de polissacarídeos totais.
De acordo Juvik & La Bonte (1988), sugary enhancer é um
duplo mutante, constituído do gene sugary e do gene sugary enhancer conjuntamente, em
homozigose recessiva (sususese). O gene sugary enhancer possui a vantagem de ter o teor
de açúcar mais elevado, mantendo as outras características desejáveis do milho sugary
como textura e estrutura do grão, germinação das sementes e vigor de plântulas, por isso
são considerados de melhor qualidade.
As dificuldades da utilização de populações com ambos os
genes associados é selecioná-las no genótipo sugary. Os efeitos fenotípicos do sugary
enhancer não são facilmente perceptíveis durante o processo de seleção de homozigotos
(Ferguson et al., 1979). La Bonte & Juvick (1990) fizeram a análise de sacarose e conteúdo
de caroteno total em uma população de grãos sugary (su-1) para sugary enhancer e
mostraram que grãos de coloração amarela clara eram um bom indicador do fenótipo de
grãos homozigóticos para o gene sugary enhancer. Além disso, o duplo mutante apresenta
uma perda menor de água durante o processo de secagem e mantém melhor qualidade por
4-5 dias em relação ao genótipo sugary.
O duplo mutante sugary enhancer (sususese) possui baixo
teor do amido nos grãos maduros, devido ao alto teor de maltose (Ferguson et al., 1979; La
Bonte & Juvick, 1990). No Brasil, ainda não existem cultivares ou híbridos comerciais com
este genótipo (Silva, 1994).
de amido e acúmulo do altos teores de açúcares, e por isso é denominado super doce.
Populações que carregam esse gene não permitem o acúmulo de polissacarídeos solúveis
em água (WSP) e com isso os grãos não apresentam textura pastosa no ponto de milho
verde como ocorre com os mutantes citados anteriormente, sugary e sugary enhancer
(Kaukis & Davis, 1986).
O gene shrunken-2 (sh-2) localiza-se no cromossomo
número 3 do milho (Glover & Mertz, 1987). Apresenta-se ligado ao gene "al", que dá a cor
da aleurona, dando uma média de crossing-over de 0,27, que é baixa. Por esse motivo,
dificilmente encontram-se grãos brancos nas espigas sh-2 (Mains, 1949).
Segundo Tracy (1994), O alelo mutante
shrunken
(
sh
),
condiciona altos teores de fitoglicogênio ou polissacarídeos solúveis em água e baixo
conteúdo de amido, por causa dos elevados teores de açúcares nesse tipo eles são chamados
de super doce.
O açúcar predominante no mutante
shrunken-2
, na fase de
grãos imaturos, é a sacarose que, segundo Tosello (1978) é o açúcar com o maior poder
adoçante. Os açúcares redutores maltose, glicose e frutose estão presentes em menores
quantidades.
Uma das vantagens do genótipo shrunken é permitir um
maior período de colheita e manter o teor de açúcar por um maior período pós-colheita, já
que a perda de água é mais demorada e o açúcar não é rapidamente convertido em amido
quando se compara com outros genótipos (Laughnan, 1953, Huelsen, 1954).
ao damping-off de pré e pós-emergência, pois o aumento do nível de açúcar deixa a planta
mais vulnerável ao ataque de patógenos e pragas do solo.
Segundo Kaukis & Davis (1986), o primeiro híbrido super
doce shrunken foi lançado nos EUA em 1961 e denominava-se ‘Illini Supersweet', mas só
em 1984 foi desenvolvido o primeiro híbrido de milho shrunken já adaptado para o
mercado de processamento.
No Brasil, as cultivares Agrocica Super Doce e Colorado
DO-0l, DO-02, DO-03 e DO-04 são exemplos de milhos super doces, contendo o gene
shrunken-2, além de populações e híbridos intervarietais desenvolvidos na Faculdade de
Ciências Agronômicas -FCA/UNESP- Botucatu.
4.4 Utiliza
ç
ão de Híbridos:
4.4.1 Heterose (vigor do híbrido)
O termo heterose foi originalmente proposto por Shull
(1908), para descrever o vigor de híbrido manifestado em gerações, derivadas do
cruzamento entre indivíduos genotipicamente divergentes, ou seja, a expressão genética dos
efeitos benéficos da hibridação. Em geral, o efeito principal esperado está relacionado a um
aumento substancial de produtividade. Porém, um grande número de outros caracteres
agronômicos economicamente importantes são, também, melhorados por meio da heterose.
esta hipótese a condição heterozigótica não seria uma condição essencial para que a
heterose se manifeste e admite a possibilidade da ocorrência de indivíduos homozigotos,
tão vigorosos quanto os heterozigotos.
A outra é a hipótese da sobredominância, que considera a
existência de alelos, com efeitos divergentes, porém favoráveis nos locos. Nesta hipótese, a
condição heterozigótica seria essencial para que a expressão do vigor se manifeste. Foi
proposta independentemente em 1908, por G.H Shull e E.M East. Posteriormente, foi
exposta por East, em 1936, de forma ampliada. Esta hipótese admite que a heterose, por si
só, é essencial à manifestação da heterose. Pressupõe a existência de alelos divergentes, no
mesmo loco, que produzem efeitos favoráveis, embora diferentes. Nesses locos, os
heterozigotos são superiores a ambos os homozigotos. Representando os dois alelos por a1
e a2, a combinação a1a2 é superior a a1a1 e a2a2. A superioridade do heterozigoto ocorre
porque cada alelo tem uma função e nele se somam as ações de a1 e de a2. Verifica-se
maior vigor nos indivíduos que têm maior número de locos heterozigóticos.
A heterose é função da diferença de freqüências alélicas dos
materiais envolvidos nos cruzamentos, e teoricamente espera-se que quanto mais
divergentes forem os materiais, maior será a heterose. Na prática nem sempre o cruzamento
de materiais mais divergentes resulta em maior heterose, pois, normalmente, esses materiais
diferem muito em adaptação e não apresentam heterose no nível esperado (Ferreira, 1992).
A heterose está normalmente mais ligada a diferenças
raciais e distância genética entre as populações. As populações, as quais em combinação
permitem que a divergência genética entre elas seja explorada em prol de uma maior
heterose, sem que haja perdas significativas quanto à adaptação dos materiais delas
gerados, constituem os "grupos heteróticos". Segundo Tsotsis (1972) citado por Bórem
(1997), a identificação desses grupos heteróticos pode determinar o sucesso de um
programa de híbridos e é um fator importante de qualquer programa de melhoramento de
milho de empresas privadas. Os sintéticos de base estreita que são feitos para
desenvolvimento de linhagens devem ser por grupos heteróticos e normalmente são
exclusivos das empresas que o desenvolveram.
Generalizando, o principal efeito esperado da heterose está
sem dúvida, relacionado a ganhos em termos de produtividade; um número relativamente
grande de características agronômicas e economicamente importantes são melhoradas e
exploradas com a utilização da heterose (Allard, 1971).
A heterose, ou o vigor de híbrido, pode ser utilizada com
vantagem, quando se tem um produto comercial de alto valor e o gasto com sementes por
área for pequeno, a exemplo do tomate e do pimentão, ou quando a semente híbrida é
produzida a preços relativamente baixos, como na caso do milho (Miranda, 1987).
4.4.2 Análise dialélica
Uma parcela importante do melhoramento é representada
pela hibridação das cultivares e linhagens, fato este, que possibilita a recombinação da
variabilidade existente, para produzir novos cultivares adaptados a diversas finalidades
(Ramalho et al., 1993). Diante do grande número de materiais disponíveis, a maior
dificuldade reside na escolha dos mais promissores para serem os genitores em um
programa de hibridação. Dessa forma, com o cruzamento entre indivíduos portadores de
fenótipos diferentes, espera-se gerar progênies superiores e conseqüentemente conduzir a
uma maior eficiência do programa. Contudo, cabe ressaltar que a hibridação poderá
conduzir tanto a indivíduos desfavoráveis quanto favoráveis (Allard, 1971).
Em programas de melhoramento, em que se opta pela
hibridação, o maior problema, e/ou a primeira grande dificuldade, corresponde à escolha
dos genitores a serem cruzados para formação da população híbrida, devido ao grande
número de materiais, normalmente disponíveis. Além disso, o melhorista terá ainda que
definir como realizar as hibridações e como as populações segregantes serão conduzidas
(Ramalho et al., 1993).
Diante do exposto torna-se evidente que o êxito de um
programa de melhoramento depende de uma escolha criteriosa e acertada dos genitores a
serem utilizados. É claro, outros fatores também influenciam nessa decisão. Dentre eles
pode-se citar a fonte de germoplasma disponível, as características a pode-serem melhoradas e o controle
genético dos caracteres (Fehr, 1987).
formarem híbridos promissores, é a de cruzamentos dialélicos (Ramalho et al., 1993). Este
método tem como finalidade expressar um conjunto de p(p-1)/2 híbridos resultantes do
acasalamento entre p progenitores (linhagens, variedades, clones, etc.), podendo incluir, além
dos respectivos pais, os híbridos recíprocos e, ou outras gerações relacionadas, tais como
F1’s, retrocruzamentos.
As metodologias de análise dialélica têm por
finalidade analisar o delineamento genético definido e prover estimativas de parâmetros
úteis na seleção de genitores para hibridação e no entendimento da natureza e magnitude
dos efeitos genéticos na determinação dos caracteres (Cruz & Regazzi, 1994).
São estimados os efeitos da capacidade combinatória,
pelo método proposto por Griffing (1956), das heteroses média, varietal e específica, pelo
método proposto por Gardner & Eberhart (1966), e os efeitos gênicos, como grau médio de
dominância, número de genes, limites de seleção, pelo método proposto por Jinks &
Hayman (1953).
Dentre os esquemas de cruzamentos dialélicos utilizados, o
que exibe todas as combinações possíveis é o da tabela dialélica completa, que corresponde
ao intercruzamento de
n
materiais, dois a dois, produzindo
n
2combinações possíveis. Essas
n
2combinações correspondem aos
n
materiais,
n(n-1)/2
híbridos simples e
n(n-1)/2
recíprocos dos híbridos simples. Porém a tabela dialélica completa não é o único esquema
de cruzamento utilizado, e algumas variações deste esquema foram introduzidas, como a
exclusão dos genitores, totalizando
n(n-1)
combinações, ou tratamentos experimentais, ou
n(n+1)/2
combinações, ou a não inclusão dos genitores e dos cruzamentos recíprocos,
utilizando apenas
n(n-1)/2
cruzamentos (Ramalho et al., 1993).
O esquema dialelo parcial, o qual corresponde ao
intercruzamento de dois grupos com
I
e
J
genitores, sendo avaliadas, experimentalmente, as
IJ
combinações híbridas e os dois grupos de genitores (Hallauer & Miranda Filho, 1982),
pode ser perfeitamente utilizado quando os outros esquemas, já mencionados, elevar muito
o número de cruzamentos.
O conceito de análise dialélica foi introduzido por Sprague &
Tatum (1942), sendo que desde então, foram propostos vários métodos para a análise das
tabelas dialélicas e, entre eles os mais empregados são os métodos de Jinks & Hayman
(1953), Griffing (1956) e Gardner & Eberhart (1966).
A análise dialélica desenvolvida por Griffing (1956), permite
estimar a capacidade geral e específica de combinação dos genitores envolvidos nos
cruzamentos dialélicos. O método pode apresentar variações de esquemas de cruzamentos,
valendo-se de quatro métodos experimentais:
Método 1- São avaliadas as
n
2combinações e inclui os parentais, os cruzamentos entre
esses parentais e seus recíprocos.
Método 2- São avaliadas as
n(n+1)/2
combinações correspondentes aos parentais e seus
cruzamentos entre esses parentais, excluindo-se os recíprocos.
Método 3- São avaliadas as
n(n-1)
combinações que correspondem aos cruzamentos e aos
recíprocos, excluindo-se os parentais.
Método 4- São avaliadas apenas as
n(n-1)/2
combinações correspondentes aos cruzamentos
Os métodos 1 e 3 permitem detectar variações genéticas dos dados
devidas as diferenças de comportamento de um dado cruzamento e de seu recíproco. Essas
variações devem-se a genes extracromossômicos, situados provavelmente nos plastídeos,
em plantas, sobretudo, nos cloroplastos e nas mitocôndrias. Esses genes são passados para
os descendentes exclusivamente através do óvulo. Assim quando se avalia um cruzamento
e também o seu recíproco, as diferenças genéticas observadas são atribuídas às
constituições genéticas do citoplasma do parental feminino de cada cruzamento (Ramalho
et al., 1993).
O procedimento de Griffing (1956) considera também os modelos
fixo e aleatório, dependendo, respectivamente, se os tratamentos experimentais fornecem
conclusões que lhe são particulares ou que podem ser extrapoladas para uma população em
que os tratamentos representam uma amostra. De acordo com Ramalho et al. (1993), no
modelo fixo apenas o efeito devido ao erro experimental é aleatório, enquanto no modelo
aleatório, apenas o efeito devido à média geral dos tratamentos experimentais é fixo. Para
esses dois modelos e os quatro métodos experimentais, podem ser efetuadas oito análises
diferentes.
5
MATERIAL E MÉTODOS
5.1 Material
Foram utilizadas e comparadas cento e vinte híbridos simples de
milho super doce portadores do gene shrunken-2 (
sh
2sh
2), e mais o híbrido Colorado
DO-04, como cultivar testemunha.
5.2 Obten
ç
ão do material experimental
Os híbridos foram obtidos a partir de um cruzamento em esquema
dialélico parcial envolvendo, no grupo 1 - quatorze linhagens braquíticas femininas (353,
365, 374, 386, 387, 421, 467, 656, 657, 662, 681, 750, 788 e 862) e, no grupo 2 - onze
linhagens braquíticas masculinas (280, 311, 320, 321, 331, 333, 522, 533, 606, 628, 636),
todas na quarta geração de endogamia (S4).
ano de 1989, tendo como objetivo obter cultivares de milho super doce com o gene
shrunken-2.
Rela
ç
ão dos híbridos e suas respectivas linhagens (genitores):
Híbridos Fêmea Macho Híbridos Fêmea Macho Híbridos Fêmea Macho Híbridos Fêmea Macho