RAFAELA COBUCI CERQUEIRA
O papel das relações sociais e da atenção social durante tarefas
cooperativas em
Callithrix jacchus
Natal
RAFAELA COBUCI CERQUEIRA
O papel das relações sociais e da atenção social durante tarefas
cooperativas em
Callithrix jacchus
Natal 2012
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre em Psicobiologia.
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Cerqueira, Rafaela Cobuci.
O papel das relações sociais e da atenção social durante tarefas cooperativas em Callithrix jacchus / Rafaela Cobuci Cerqueira. - Natal, RN, 2012.
59 f. : Il.
Orientadora: Profa. Renata Gonçalves Ferreira.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia. 1. Callithrix jacchus – Dissertação 2. Cognição social – Dissertação. 3. Cooperação – Dissertação. I. Ferreira, Renata Gonçalves. II. Universidade Federal do Rio Grande do Norte. IV. Título.
Título: O papel das relações sociais e da atenção social durante tarefas cooperativas em Callithrix jacchus
Autora: Rafaela Cobuci Cerqueira
Data da defesa: 30 de março de 2012
Banca Examinadora:
_________________________________ Profa. Briseida Dôgo de Resende
Universidade de São Paulo, SP
__________________________________ Profa. Maria de Fátima Arruda de Miranda Universidade Federal do Rio Grande do Norte, RN
__________________________________ Profa. Renata Gonçalves Ferreira
Dedico este trabalho à
memória do meu avô Manoel
Cobucci e à da minha
gatinha Natalina, dois dos
maiores exemplos de força
AGRADECIMENTOS
A primeira pessoa a quem eu quero agradecer é a professora Renata, por ter me dado essa oportunidade e acreditado em mim. Muito obrigada por toda a paciência, pelos ensinamentos, e também pelo carinho e compreensão em todos os momentos.
Agradeço imensamente também ao professor John, que me acolheu em Natal, me dando oportunidade para ficar na cidade e começar tudo.
Agradeço aos professores do programa de Pós Graduação em Psicobiologia pelos ensinamentos e pelos empréstimos de livros, artigos e teses.
À professora Regina Macedo pelas valiosas observações na minha qualificação.
À banca examinadora pela disponibilidade para contribuir com meu trabalho e pelas críticas e sugestões para melhorá-lo.
Ao CNPq pela bolsa de mestrado.
Aos funcionários do Núcleo de Primatologia pelo cuidado com os saguis.
Aos colegas de mestrado pelas ótimas aulas, conversas e discussões de livros e artigos.
Ao pessoal do LabCrono, por terem me ajudado quando cheguei em Natal e por não se importarem com a minha presença nas suas salas.
Aos amigos Dani e Ricardo pelo apoio desde o início.
À Eloysi e Suedja pela indispensável ajuda e ótima companhia durante os experimentos. Aos amigos Kalina, Denise, Flávia, Rodrigo, Sol, Silvia e Ione por todo apoio e carinho e pela enorme ajuda com os animais de rua nos inúmeros resgates durante o mestrado.
À minha amiga Clarissa, simplesmente por ser amiga.
Devo agradecer também aos saguis que ficaram comigo até o final do estudo e todos os que participaram de alguma etapa desse trabalho.
Aos meus pais e irmão que sempre me apoiaram em todas as decisões que eu tomei, não me contrariando nunca, nem quando eu inventei de fazer mestrado nessa lonjura.
À minha cunhada Juliana que sempre tinha uma receita especial pra mim. Aos meus tios e primos por torcem por mim sempre.
Um agradecimento especial a todos os bichos que passaram pela minha vida antes e durante o mestrado, e que me ensinaram tanto, principalmente aqueles que ficaram comigo: Look, Natalina e Tigrão (agora em pensamento), Miguel, Bela, Princesa, Gabi, Rapunzel e Nala, as cinco últimas sincronizando seus ritmos circadianos ao meu ritmo de estudo até o fim para me fazer companhia o tempo todo.
SUMÁRIO
RESUMO ... 1
ABSTRACT ... 2
INTRODUÇÃO ... 3
1. Relações Sociais e Comportamentos cooperativos ... 5
2. Cognição Social em Callithrix jacchus ... 10
Habilidades Cooperativas ... 11
Olhares sociais em tarefas cooperativas ... 14
OBJETIVOS ... 17
HIPÓTESES E PREDIÇÕES ... 18
METODOLOGIA ... 20
Animais e alojamento ... 21
Materiais ... 22
Procedimento experimental dos testes cooperativos ... 24
Teste de cooperação ... 25
Teste de prosocialidade ... 26
Teste controle ... 26
1. Relações Sociais ... 27
Coleta de dados ... 27
Análise de dados ... 28
2. Atenção Social ... 30
Coleta de dados ... 30
RESULTADOS ... 33
1. Relações Sociais ... 33
Primeiro set ... 33
Segundo e terceiro sets ... 34
2. Atenção Social ... 35
Primeiro set ... 35
Segundo e terceiro sets ... 37
DISCUSSÃO ... 40
1. Relações Sociais ... 40
2. Atenção Social ... 42
CONCLUSÃO ... 48
RESUMO
ABSTRACT
Several studies on nonhuman primates show that the relationships between individuals strongly influence the expression of cooperative behavior, both in natural environment and in captivity settings. Recent studies suggest that cooperative breeders present outstanding performance in tasks involving social cognition, such as cooperative tasks with experimental apparatuses. In experimental research on this subject it is crucial to differentiate between real cooperation (or communicative cooperation, mediated by social attention) and by-product cooperation that results from simultaneous actions of individuals. The present study assessed, in Callithrix jacchus, a cooperative breeder species, if social relationships and social attention between subjects are important factors during performance in cooperative tasks. During the experimental procedure the animals participated in three different cooperative tasks: cooperation task, prosocial task and control task. Diverging from the literature, matrix
INTRODUÇÃO
Endêmica do Brasil, a espécie Callithrix jacchus, conhecida como sagui comum, está entre os menores antropóides (o indivíduo adulto pesando entre 300 e 400 g, Araujo et al. 2000) e faz parte da família Callitrichidae de primatas do Novo Mundo que habitam principalmente a América Central e a do Sul (Rylands et al. 2009).
São reconhecidos atualmente sete gêneros na família: Saguinus, Leontopithecus, Callimico, Callithrix, Cebuella, Callibella, e Mico (Cortés-Ortiz 2009). Esses animais são
diurnos, arborícolas (Stevenson & Rylands 1988; Rylands et al. 1993) e alimentam-se principalmente de frutos, artrópodes, e gomas (Nash 1986), sendo sua ecologia comportamental fortemente influenciada pela grande variação sazonal na disponibilidade de recursos na maioria dos habitats neotropicais que eles habitam.
A análise do comportamento cooperativo em primatas tem revelado que dois fatores são importantes para a execução de tarefas cooperativas: as condições sociais e a cognição (para revisão, Werdenich & Huber 2002) o que significa que tanto as competências sociais (quem coopera com quem) quanto a capacidade individual (o entendimento da situação) são relevantes (Chalmeau & Gallo 1996b). A tolerância social tem sido identificada como um importante fator que influencia a ocorrência desses comportamentos nos primatas (para revisão, Anderson 2007) e os indivíduos tendem a formar pares cooperativos com aqueles animais que eles têm mais afinidade social (Crofoot et al. 2011). Ainda, a formação de pares cooperativos pode ocorrer em um sistema de trocas, com os indivíduos trocando favores como catação por suporte em conflitos (Seyfarth & Cheney 1984; Di Bitetti 1997).
Este texto discute resultados de uma pesquisa voltada à análise destes dois fatores em Callithrix jacchus e está estruturado da seguinte forma. Primeiramente é feita uma descrição
de trabalhos que mostram uma ligação entre relações sociais e comportamentos cooperativos em primatas (tema 1). Em seguida, é apresentada uma revisão da literatura acerca das habilidades cognitivas do sagui comum, incluindo trabalhos que mostram suas habilidades cooperativas e trabalhos que oferecem suporte à idéia de que esses animais apresentam os elementos de cognição social para engajar em comportamentos cooperativos verdadeiros, mediados por atenção social (tema 2). Com esta base teórica foram elaborados experimentos visando explorar a importância das relações sociais e da atenção social para a realização de tarefas cooperativas em dois grupos familiares cativos de saguis comuns. São apresentados os objetivos, as hipóteses, a metodologia, os resultados e a discussão também de forma a identificar esses dois temas. Dois artigos estão sendo elaborados; o primeiro será submetido para a revista A Primatologia no Brasil e o segundo para a revista Behavioural Processes.
1. Relações Sociais e Comportamentos Cooperativos
Em uma abordagem sobre o tema, duas espécies do gênero Macaca foram estudadas em um estudo comparativo. A primeira, Macaca tonkena é uma espécie altamente tolerante quando comparada com a outra, Macaca mulatta, que vive em relações despóticas (Petit et al. 1992, para revisão Anderson 2007). Os autores utilizaram pedras pesadas sobre itens alimentares para testar a habilidade dos indivíduos cooperarem em cada uma das espécies, uma vez que um indivíduo sozinho não era capaz de retirar a pedra, mas precisava da ajuda de outro co-específico. A retirada da pedra por dois indivíduos foi mais comum na espécie mais tolerante.
Em seu trabalho com chimpanzés, Melis et al. (2006) mostraram que indivíduos que se alimentam em proximidade espacial e de forma pacífica são mais propensos a cooperar em uma tarefa que requer ação simultânea de dois animais para obtenção de recompensas, concluindo que a tolerância age como uma importante limitação durante a realização dessas tarefas na espécie. Em outro estudo (Hare et al. 2007), a tendência para cooperar foi comparada entre bonobos e chimpanzés, os primeiros bem mais tolerantes que vivem em relações sociais mais pacíficas que os últimos. Foi identificado que os bonobos eram mais propensos a se alimentarem juntos e pacificamente que seus parentes mais próximos e que apresentavam maior tendência a cooperar quando a recompensa obtida era facilmente monopolizável (Hare et al. 2007). Essa diferença não existiu, entretanto, quando a recompensa podia ser dividida. Os autores concluem que, mesmo quando dois indivíduos são capazes de entender que é necessária a ajuda de outro animal para a resolução de uma tarefa, a cooperação pode não ocorrer como resultado de baixa tolerância social (Hare et al. 2007).
cooperativo está diretamente relacionado com a proximidade espacial entre os indivíduos, sendo o suporte oferecido para o animal com maior índice de proximidade com o interventor.
Estudos com macacos prego revelam também que além de apresentarem comportamentos cooperativos na natureza, como a coalizão (Ferreira et al. 2006) e transferência de alimento (Ferreira et al. 2002) esses animais também cooperam em tarefas que utilizam aparatos (Chalmeau et al. 1997b; Visalberghi et al. 2000; Mendres & de Waal 2000; de Waal & Berger 2000; Hattori et al. 2005).
Os calitriquídeos também se destacam nessas tarefas (Werdenich & Huber 2002; Hauser et al. 2003, Cronin et al. 2005), e são uma escolha evidente para estudos sobre cooperação e as relações sociais ligadas a esse comportamento. Eles apresentam um sistema de cuidado cooperativo dos infantes (Stevenson & Rylands 1988), partilham alimento (Ferrari 1987), incluindo partilha ativa (para revisão, Menezes 2010), são altamente tolerantes uns com os outros (Ferrari 1987; Kasper et al. 2008) e mantém coesão espacial e comportamental com seus parceiros sociais (Digby & Barreto 1993). Para Cronin & Snowdon (2008), o sistema social cooperativo dessa família e o forte vínculo que pode ser formado entre pares de indivíduos podem contribuir para seu sucesso na realização de tarefas cooperativas, destacando-os entre outras espécies mais competitivas ou sem fortes relações sociais.
cooperativo dos infantes e a alta tolerância da espécie foram identificados como os responsáveis por tal resultado (Burkart & van Schaik 2012).
Em um estudo sobre os principais fatores que influenciam o comportamento cooperativo em Callithrix jacchus em uma tarefa instrumental, Werdenich & Huber (2002) investigaram quais condições sociais poderiam determinar a ocorrência desse comportamento, e deram ênfase na análise de tolerância inter-individual e distribuição de papéis para cooperar. As duplas que foram bem sucedidas no teste de cooperação foram aquelas que, em uma fase anterior de treinamento, o subordinado assumia o papel de ativo puxando uma corda do aparato para o dominante receber uma recompensa. Os animais dominantes não forçaram os subordinados a tal atividade e compartilharam a recompensa com seus parceiros, ressaltando a importância da tolerância dos primeiros em relação aos segundos para a ocorrência de cooperação. Os autores reconheceram que a cooperação depende, portanto, de fatores sociais como a classificação na hierarquia, a distribuição específica de papéis na execução da tarefa e a tolerância de indivíduos dominantes com relação aos subordinados.
Outro medidor de afinidade social é a catação. Um exemplo clássico de trabalho que correlaciona a catação com o comportamento cooperativo de suporte em conflitos é aquele de Seyfarth & Cheney (1984), onde os autores estudaram fêmeas de macacos vervet (Chlorocebus aethiops) e testaram, com a ajuda de gravações de vocalizações, se a catação realizada por um indivíduo A em um indivíduo B antes do teste aumentaria a possibilidade do indivíduo B atender a solicitação de suporte do animal A. Seus resultados mostraram que, para indivíduos não aparentados, os animais pareceram mais propensos a dar suporte àqueles que haviam os catado recentemente. Posteriormente a este trabalho, diversos outros estudos evidenciam um padrão de troca de catação por favores sociais (Henzi & Barrett 1999; Barrett et al. 2012). Cheney et al. (2010) utilizaram metodologia semelhante àquela do trabalho
comportamento semelhante às fêmeas de macacos vervet. A associação entre catação e comportamentos cooperativos também foi identificada em bonobos (Legrain et al. 2010) e chimpanzés (Foster et al. 2009).
Dunbar (1991) sugere que a catação nos primatas do Novo Mundo tem função principal de higiene, ao contrário do que ocorre nos primatas do Velho Mundo, que, segundo o autor tem função principalmente social. Contrária à idéia de Dunbar, diversos estudos já mostraram que a catação é uma importante ferramenta para medir relações em espécies do Novo Mundo (Ateles geoffroyi: Ahumada 1992; Cebus apella: Di Bitetti 1997; Alouatta seniculus: Sanchez-Villagra et al. 1998; Callithrix jacchus: Lazaro-Perea et al. 2004). Di
2. Cognição Social em Callithrix jacchus
Muitos primatas vivem em grupos de dois ou mais indivíduos. Esses grupos podem ser de organização social simples, como em algumas espécies de prosímios que só se agregam para dormir, ou de organização social complexa como a que ocorre nos grandes símios e em espécies da família dos calitriquídeos (Smuts et al. 1987). Independente da forma de vida social, problemas cognitivos no domínio social são constantes e para sobreviver é necessário saber lidar com eles.
A hipótese da inteligência social sugere que uma maior cognição evoluiu como uma conseqüência da necessidade de manobras comportamentais para lidar com as complexidades da vida social (Byrne & Whiten 1988), e propõe que o tamanho do encéfalo (mais especificamente o neocortex) também evoluiu nesse sentido (para revisão, Shettleworth 2010). Apesar de apresentarem encéfalos relativamente pequenos, tanto em volume absoluto quanto em tamanho absoluto e relativo do encéfalo funcional, estudos recentes têm destacado muitas habilidades cognitivas no domínio físico e principalmente no domínio social em calitriquídeos (para revisão, Huber & Voelkl 2009). Estudos já demonstraram que algumas espécies são capazes de se beneficiar de influências sociais que ajudam no aprendizado sobre novos tipos de alimentos (Vitale & Queyras 1997; Caldwell, et al. 1999; Caldwell & Whiten 2003; Yamamoto & Lopes 2004; Voelkl et al. 2006; Schiel & Huber 2006) e aprender novas técnicas de forrageio por meio da imitação (Bugnyar & Huber 1997; Voelkl & Huber 2000; Caldwell & Whiten 2004). Alguns destes estudos apresentam evidências de que o aprendizado social é uma importante forma de aprendizado em saguis (Huber & Voelkl 2009).
& Huber 2002; Burkart et al. 2007) e muito do que sabe a respeito de comportamentos cooperativos depende do conhecimento sobre o sistema de cuidado cooperativo da prole em Callithrix jacchus (Koenig & Rothe 1991).
Habilidades cooperativas
O sistema de criação cooperativa da prole em Callithrix jacchus constitui, por si só, um importante exemplo de comportamento cooperativo na espécie. Quando os ajudantes são parentes dos filhotes, a ajuda pode ser explicada por seleção de parentesco (Hamilton 1964) e quando não há parentesco com a prole, existem outros potenciais benefícios para os ajudantes cuidadores, dentre eles o acesso aos recursos do grupo em que vive e o aprimoramento das habilidades de cuidado à prole que lhes podem ser úteis no futuro (Emlen 1991; Tardif 1997). Apesar de todos os indivíduos não reprodutores se envolverem no cuidado cooperativo, nem todos os membros do grupo contribuem da mesma maneira. Sabe-se que os indivíduos mais velhos transportam os infantes mais freqüentemente (Yamamoto & Box 1997) e partilham mais alimento com eles (Ferrari 1987).
defesa de território, mesmo tendo custos, fornece benefícios relacionados a decisões de dispersão, sendo possível aos ajudantes avaliar potenciais parceiros bem como aumentar sua familiaridade com grupos vizinhos.
Em laboratório, o mesmo estudo que verificou a influencia de fatores sociais na cooperação em aparatos experimentais, submeteu os animais a uma tarefa cooperativa que requeria divisão de tarefas e coordenação de atividades: um parceiro deveria puxar a corda do aparato experimental para que o outro pudesse alcançar a recompensa. As díades formadas foram capazes de cooperação ao coordenarem seus comportamentos no tempo e no espaço (Werdenich & Huber 2002).
Com relação à Callithrix jacchus, Burkart et al. (2007) sugerem que essa espécie também apresenta comportamento prosocial, provisionando alimento de maneira espontânea e altruísta a indivíduos não aparentados (Burkart et al. 2007). Em seu experimento, os animais foram isolados de seus grupos e confrontados com duas opções de bandeja numa tarefa cooperativa: uma contendo recompensa somente para o parceiro (0,1) e outra sem nenhuma recompensa (0.0). Foi observado que os animais puxavam mais a bandeja (0,1) quando um parceiro estava presente na gaiola adjacente à sua do que na condição controle, quando o indivíduo ativo estava sozinho e a gaiola adjacente estava vazia. O mesmo grupo de pesquisadores fez experimentos parecidos, mas dessa vez com todos os sagüis presentes no mesmo viveiro e os resultados encontrados também sustentam que esses animais doam alimento para seus parceiros espontaneamente e na ausência de solicitação (Burkart & van Schaik 2012).
Evidência adicional sobre as habilidades cooperativas no sagüi comum foi reportada por Kasper et al. (2008) em um trabalho que mostrou a ocorrência de transferência de alimento entre indivíduos adultos. Os autores observaram transferência boca a boca entre os animais que, durante a partilha, abriam as bocas uns dos outros para ter acesso à comida e essa transferência ocorreu tanto quando os indivíduos subordinados eram os possuidores do alimento quanto na direção contrária.
sócio cognitivas como o aprendizado social e tarefas que necessitam o entendimento da comunicação visual; ajustar sistemas de comunicação através da sinalização cooperativa; favorecer o comportamento prosocial em contextos outros que não o de cuidado da prole, e por fim, devem aumentar o desempenho em tarefas cooperativas que exigem habilidade para coordenar ações com outros indivíduos no tempo e no espaço (Burkart & van Schaik 2010). Os autores sustentam que as três últimas habilidades implicam numa maior capacidade para a resolução de problemas em tarefas cooperativas, o que se deve principalmente à cognição social característica e diferencial das espécies que apresentam cuidado cooperativo dos filhotes (Burkart et al. 2009; Burkart & van Schaik 2010).
Olhares Sociais em Tarefas Cooperativas
Os trabalhos que estudam a habilidade dos primatas de resolver tarefas cooperativas em cativeiro têm buscado compreender por que mecanismos proximais tais habilidades são mantidas (para revisão, Werdenich & Huber 2002; Noe 2006). Além das relações entre os indivíduos que são importantes determinantes para a realização dessas tarefas, como visto anteriormente, alguns estudos se interessam em conhecer as capacidades cognitivas subjacentes aos comportamentos realizados. Vários estudos têm demonstrado que os primatas utilizam diversos mecanismos cognitivos para monitorar os comportamentos de co-específicos em diferentes interações sociais (Tomasello & Call 1997; Tomasello 2000). Esses mecanismos cognitivos são considerados adaptações comportamentais de grande valia na complexa rede de demandas sociais na qual os indivíduos estão inseridos.
distinguir a cooperação verdadeira da cooperação que ocorre como resultado de ações individuais simultâneas.
Considera-se que, quando ocorre comunicação entre os animais, e quando esses estão atentos aos seus parceiros identificando a importância do papel desses durante a realização de uma tarefa, há a compreensão da tarefa cooperativa. Sem a comunicação ou a atenção social, ao contrário, a resolução de uma tarefa é considerada como um subproduto de atividades coincidentes (para revisão, Werdenich & Huber 2002). Para Chalmeau & Gallo (1996b) a troca de informação entre os indivíduos envolvidos na resolução de uma tarefa cooperativa é de extrema importância para o ajuste de seus comportamentos.
Vários estudos experimentais já identificaram que primatas do Velho Mundo são capazes de compreender tarefas de cooperação e entender o papel de outros indivíduos na realização destas (chimpanzés: Boesch & Boesch 1989; Chalmeau 1994; Chalmeau & Gallo 1996a; Chalmeau & Gallo 1996b; Hirata & Fuwa 2007; orangutangos: Chalmeau et al. 1997a). Os primatas do Novo Mundo também têm sido estudados e a cooperação verdadeira já foi reportada em algumas espécies tais como o macaco prego (Mendres & de Waal 2000; mas ver Chalmeau et al. 1997b) e o tamarin (Cronin et al. 2005).
tarefa, e puxavam mais a alavanca do aparato quando um parceiro estava presente do outro lado do que quando estava ausente. Os autores argumentam que esses resultados fornecem evidências de que os macacos prego entenderam a tarefa cooperativa e foram capazes de levar em conta a ação de seu parceiro para a realização da mesma.
Aos animais com cuidado cooperativo dos infantes, atribui-se um desempenho diferencial em tarefas que necessitam o entendimento da comunicação visual (Burkart et al. 2009; Burkart & van Schaik 2010). O trabalho de Cronin et al. (2005) defende que na espécie Saguinus oedipus da família Callitrichidae, o indivíduo é capaz de entender o papel de um parceiro durante tarefas cooperativas, modificando seu comportamento de acordo com a presença ou não de um outro animal em um aparato que requer que dois animais trabalhem simultaneamente para obter recompensas. Para esses autores, outras espécies de calitriquídeos devem ter desempenho similar em tarefas cooperativas desse tipo.
OBJETIVOS
Este estudo teve como objetivo estudar o comportamento cooperativo do sagui comum (Callithrix jacchus) em cativeiro durante a manipulação de aparatos em tarefas cooperativas. Os objetivos específicos foram:
1. Investigar se a tolerância social e a afiliação (medidas por meio da proximidade e do comportamento de catação) entre os indivíduos no contexto familiar estão relacionados com a execução de tarefas cooperativas (tarefas de cooperação e de prosocialidade) por eles.
HIPÓTESES E PREDIÇÕES
Hipótese 1a: A afiliação entre os indivíduos facilita a cooperação.
Predição 1a: Espera-se que os indivíduos que passem maior tempo em proximidade e em catação também cooperem mais por meio da coordenação de atividades, ou seja, deve haver uma correlação positiva entre essas relações e as puxadas no aparato no teste de cooperação.
Hipótese 1b: A afiliação entre os indivíduos favorece o comportamento prosocial.
Predição 1b: Espera-se que relações afiliativas anteriores facilitam o comportamento prosocial, ou seja, que os animais que realizam catação e ficam próximos com mais freqüência sejam os que mais doem alimento de forma prosocial aos seus parceiros.
Hipótese 1c: A catação e o comportamento prosocial entre os indivíduos ocorrem em um sistema de troca de favores.
Predição 1c: Espera-se que os animais que realizam catação com mais freqüência sejam os que mais recebam alimento de seus parceiros.
Hipótese 2a: A cooperação em saguis é verdadeira, ou seja, mediada por atenção social. Predição 2a: Espera-se uma correlação positiva entre a execução das tarefas (puxadas no aparato no teste de cooperação) e os olhares sociais (olhares entre os indivíduos envolvidos nas tarefas).
Hipótese 2b: O comportamento prosocial em saguis é verdadeiro, ou seja, mediado por atenção social.
METODOLOGIA
A literatura diverge na definição do termo “cooperação”, e termos amplamente usados
como cooperação, mutualismo, reciprocidade, altruísmo recíproco e simbiose são às vezes usados como sinônimos e outras vezes como diferentes categorias do comportamento cooperativo. Ainda, o que alguns autores consideram cooperação é considerado por outros como um subproduto de ações individuais coincidentes se não há compreensão da tarefa pelos animais envolvidos na realização da mesma (para revisão Noe 2006). Tais divergências têm dificultado conclusões acerca do assunto e a troca de informações entre os pesquisadores da área. Neste trabalho empregamos o termo cooperação fazendo referência a comportamentos onde os animais operam em conjunto, sendo a cooperação considerada verdadeira quando os indivíduos estão atentos ao parceiro para coordenar suas atividades para realizar uma tarefa (Mendres & de Wall 2000; Werdenich & Huber 2002).
A palavra prosocial é outro termo da literatura que possui significados diferentes de acordo com o autor que a utiliza ou com o campo de estudo em que ela é empregada. A definição de comportamentos prosociais aqui utilizada é a aquela de Burkart et al. (2009), que os define como aqueles comportamentos que produzem benefício a outros indivíduos (Burkart et al. 2009). Uma análise distal pode revelar que esses comportamentos são resultado de
seleção de parentesco, altruísmo recíproco, mutualismo, manipulação, etc. Porém, os meios evolutivos pelos quais eles são mantidos não são o foco do presente trabalho. Aqui, a análise é feita em nível próximo, com a investigação do papel das relações sociais e da atenção social para a ocorrência desses comportamentos, sendo o comportamento prosocial considerado verdadeiro quando há atenção e compreensão da tarefa pelos animais. Por fim, o termo atividades/tarefas/habilidades cooperativas será utilizado para fazer alusão a ambos os tipos
Primeiramente será descrita a metodologia do trabalho que foi comum aos estudos sobre os temas 1 e 2 e depois apresenta-se a metodologia específica para cada um deles.
Animais e alojamento
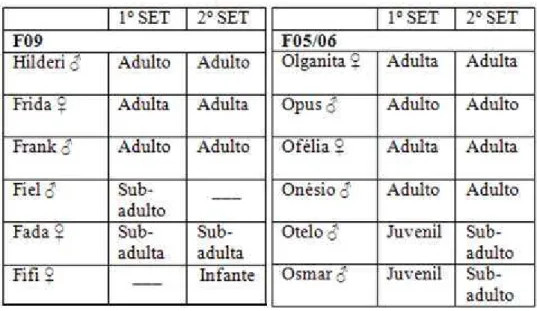
Foram estudados dois grupos familiares cativos de Callithrix jacchus do Núcleo de Primatologia da Universidade Federal do Rio Grande do Norte. Cada família estava alojada em um recinto de alvenaria com portões de ferro e piso de cimento, com 2x1x2 m, equipado com plataformas e troncos de madeira, comedouros, bebedouro e caixa ninho. Cada recinto era parcialmente coberto com telhas de cerâmica, permitindo que os sagüis ficassem expostos à temperatura e ciclo de luz naturais. Os animais mantinham contato visual, auditivo e olfativo com indivíduos de outras famílias do núcleo. A tabela 1 apresenta a composição das duas famílias que foram estudadas (F09 e F05/06), bem como o sexo e a classificação etária dos animais segundo Yamamoto (1993).
Tabela 1: Composição e classificação etária das duas famílias estudadas *.
Os animais eram identificados pela marcação dos tufos periauriculares, dorso, patas dianteiras e/ou patas traseiras com solução de ácido pícrico à 5%. Essa substância faz com que as partes do corpo adquiram coloração amarelada.
Materiais
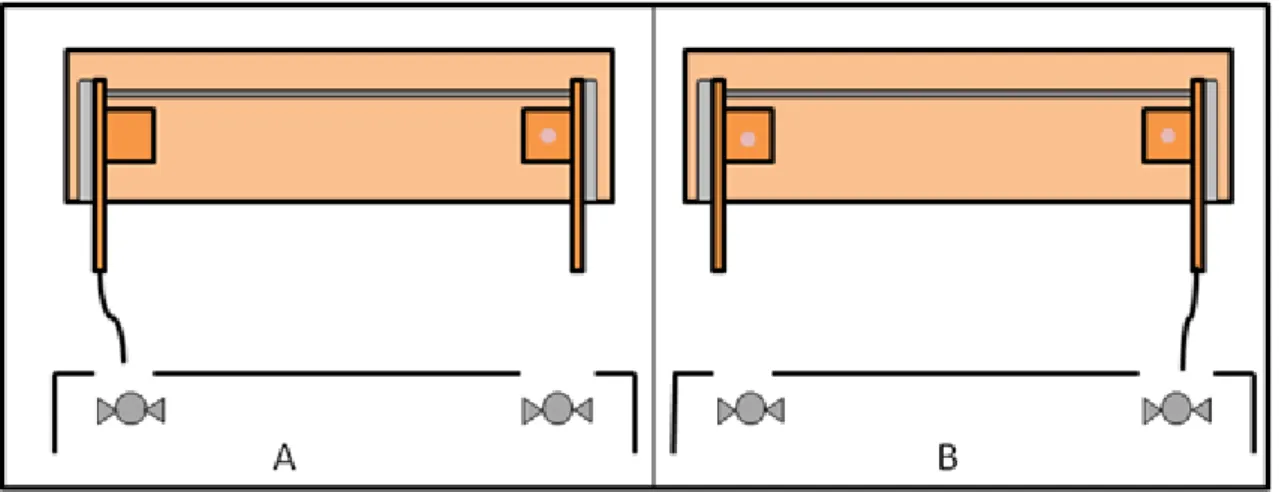
Dois aparatos experimentais idênticos foram utilizados na realização dos experimentos. Cada aparato experimental (figuras 1 e 2) consistiu de uma plataforma de madeira contendo duas alavancas separadas entre si por aproximadamente 40 cm. As duas alavancas eram presas em trilhos de alumínio, o que permitia que elas se movimentassem como gavetas, e estavam conectadas uma a outra por uma barra. Em uma das extremidades de cada alavanca estava conectada uma bandeja e cordas foram dispostas nas outras extremidades para permitir que os animais puxassem as alavancas e tivessem acesso às bandejas com as recompensas.
Foram feitos três diferentes testes experimentais. O primeiro teste foi denominado Teste de cooperação (figura 1), o segundo teste foi chamado Teste de prosocialidade (figura
Figura 1: Desenho esquemático do aparato experimental utilizado no Teste de cooperação. Neste teste, ambas as bandejas continham uma recompensa alimentar, de maneira que os dois animais envolvidos na tarefa cooperativa recebiam alimento.
Os reforços alimentares utilizados foram pedaços de banana com aproximadamente 0,5 cm2. Todos os experimentos foram gravados por meio de câmeras de vídeo que eram dispostas em tripés na frente dos viveiros durante os experimentos (figura 3). Computadores foram utilizados para posterior análise dos vídeos e dos dados.
Procedimento experimental dos testes cooperativos
Os experimentos foram feitos após uma fase de treinamento dos animais para operação dos aparatos. O treinamento foi feito de forma que ações individuais resultavam em obtenção individual de alimento, isto é, os saguis foram treinados a puxar as alavancas quando a barra que as conecta havia sido removida. Assim, quando um animal puxava uma alavanca, ele recebia a recompensa da bandeja e não era necessária a ação simultânea de puxar as alavancas para obtenção dos reforços alimentares.
Os animais foram treinados por duas semanas em sessões de aproximadamente 10 minutos. Nenhum indivíduo foi manipulado de modo a minimizar o estresse dos animais, por isso o treinamento de cada sagui ocorreu de forma aleatória, à medida que os animais estabeleciam contato e manipulação com os aparatos.
como as sessões experimentais foram conduzidos com todos os indivíduos de cada família presentes nos viveiros.
Figura 3: Disposição dos aparatos experimentais e das câmaras de vídeo na frente do viveiro dos animais.
Testes de cooperação
Após o treinamento, teve início a fase de teste de sincronismo de atividades para obtenção conjunta de recompensas alimentares (Teste de cooperação). Neste experimento foi testada a capacidade de cooperar dos animais.
Teste de Prosocialidade
Esse teste analisou a ocorrência de comportamento prosocial nos saguis, ou seja, se os animais eram capazes de executar uma tarefa em benefício de um parceiro. Assim como no teste de cooperação, dois aparatos (Figura 2a) ficaram disponíveis para os animais durante as sessões. Ao puxar uma alavanca, as duas bandejas chegavam ao alcance de cada animal no aparato, mas somente a bandeja do animal que não puxou continha a recompensa. O indivíduo que puxava e nunca recebia a banana podia ver que seu parceiro era beneficiado com a sua ação.
Teste Controle
Um terceiro teste experimental foi realizado para comparação e controle dos resultados obtidos nos dois primeiros testes. Tudo foi feito de modo similar ao teste de prosocialidade, mas aqui, os dois indivíduos recebiam a fruta a partir da ação de somente um animal que puxava a corda da alavanca.
Nos três testes, assim como no treinamento, as duplas de indivíduos de cada aparato (um animal na frente de cada bandeja) também se estabeleceram de maneira espontânea, já que os animais não foram manipulados e estavam todos juntos nos viveiros.
1. Relações sociais
Coleta de dados
O padrão de relação social de cada indivíduo foi registrado por meio de comportamentos afiliativos de catação e proximidade e os registros foram obtidos por meio do método focal instantâneo (Altmann 1974) com intervalos de 30 segundos entre registros, durante dez minutos. A catação foi definida como a exploração dos pêlos realizada com as mãos ou com a boca, que podia ocorrer entre dois ou mais indivíduos. Incluiu receber catação, quando o animal focal recebia catação de algum outro membro do grupo, e fazer catação, quando o animal focal dirigiu este comportamento a outro animal. A proximidade foi considerada quando o animal focal encontrava-se a até 15 cm de outro (s) indivíduo (s) (Lopes 2002). Esses dados das relações sociais foram coletados de duas maneiras:
- durante dez dias antes de ter início os testes cooperativos do primeiro set, e durante outros dez dias antes de ter início os testes cooperativos do segundo set. A idéia inicial era fazer um único registro, antes de submeter os animais aos experimentos com os aparatos, porém, a composição da família F09 variou entre o primeiro e o segundo sets, mas permaneceu a mesma durante o segundo e o terceiro (ver tabela 1). Além disso, houve um intervalo nos experimentos entre o primeiro e o segundo set, devido a reformas no Núcleo de Primatologia, e considerou-se, portanto, que as condições do primeiro set poderiam ser diferentes daquelas do segundo e do terceiro. Por causa desse intervalo, foram feitas duas coletas dos dados das relações sociais.
- diariamente, no dia de cada experimento do segundo e do terceiro sets, na hora anterior a cada sessão experimental. Durante o primeiro set não foram registrados dados sociais nessas condições.
ela foi definida como um animal segurar a corda da alavanca fazendo a bandeja deslizar trazendo o reforço ao seu alcance. No teste de cooperação elas ocorriam sempre em duplas, enquanto que nos testes de prosocialidade e controle elas eram exercidas por um indivíduo somente, o emissor.
Análise de dados
Com relação aos dados coletados por dez dias antes do início dos testes, a análise dos dados do primeiro set foi feita separadamente daquela do segundo e do terceiro, que foram analisados juntos. Os motivos para as análises serem organizadas dessa maneira já foram descritos acima. Com relação aos dados coletados diariamente antes de cada sessão, o segundo e o terceiro sets também foram analisados juntos.
Para verificar a importância das relações sociais entre os animais para a cooperação e a prosocialidade, testes de correlação de matrizes foram realizados. Como as díades nos aparatos foram formadas aleatoriamente, houve muitas possibilidades de formação de duplas, e durante a análise dos dados isso leva a uma dependência estatística. Foi utilizado então o teste Tau Kr para correlação de matrizes, uma vez que tal teste leva em consideração a dependência dos dados (Hemelrijk, 1990a, b).
Para cada família, e para os três sets (o primeiro separado, e o segundo e o terceiro juntos) para o teste de cooperação foi criada uma matriz quadrada simétrica onde cada célula continha número de vezes que os animais puxaram juntos a corda. Para o teste de prosocialidade foi criada uma matriz quadrada assimétrica de puxadas, onde as linhas representavam o número de puxadas do indivíduo emissor (o que puxava a corda) para seu parceiro receptor em cada díade formada.
puxadas do teste de prosocialidade, e por isso, era assimétrica. Uma matriz de proximidade quadrada simétrica foi criada para correlação com as puxadas dos dois testes cooperativos.
A matriz de puxadas assimétrica do teste de prosocialidade foi transposta para ser correlacionada com a matriz de catação para se testar a hipótese sobre a troca de favores entre catação e doação de alimento (Hipótese 1c).
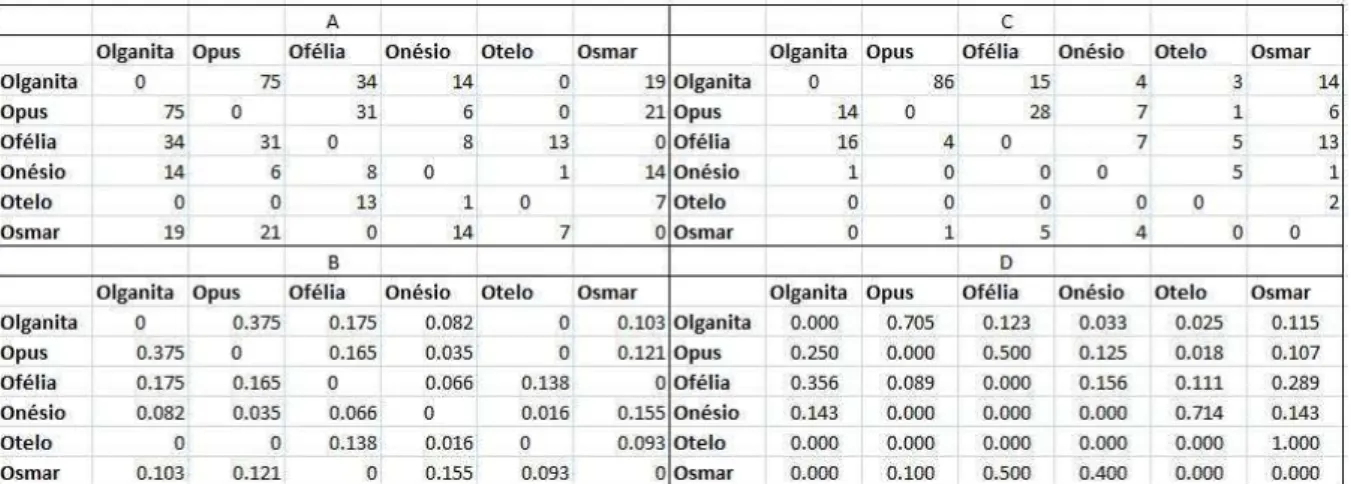
Como os indivíduos participaram em quantidades diferentes dos testes experimentais, todas as matrizes foram normalizadas para se retirar o possível efeito de um viés no número de puxadas que poderia existir se diferentes indivíduos tivessem diferentes motivações para participar das tarefas. Tanto para as puxadas quanto para as relações sociais, o índice de Jaccard foi utilizado como um método de normalização nas matrizes simétricas. Para as matrizes assimétricas a normalização foi feita dividindo-se o número de puxadas/catação do emissor A para o receptor B pelo número total de puxadas/catação do emissor A para todos os indivíduos receptores com os quais ele formou dupla. A figura 4 mostra um exemplo de matriz simétrica e outro de matriz assimétrica antes e depois das normalizações.
Figura 4: Exemplo de uma matriz simétrica antes (a) e depois (b) da normalização e de uma matriz assimétrica antes (c) e depois (d) da normalização.
2. Atenção social
Coleta de dados
A filmagem dos testes cooperativos permitiu o registro das três variáveis de interesse para o estudo do papel da atenção social nestes testes. São elas: as puxadas, os olhares sociais e a presença dos indivíduos em cada bandeja do aparato.
O olhar social foi definido como a movimentação da cabeça de um animal em direção ao parceiro que estava na outra bandeja no mesmo aparato, e a ele foi atribuída função de atenção social (Mendres & de Waal, 2000). As puxadas, como já foi dito, definiram a execução das tarefas e a presença de cada animal na bandeja foi quantificada a cada 30 segundos.
Análise de dados
prosocialidade a matriz criada foi assimétrica com o número de olhares dos indivíduos emissores (os que puxavam a corda) para seus parceiros receptores em cada dupla estabelecia. As matrizes de presença dos animais nas bandejas eram simétricas para o teste de cooperação e assimétricas para o teste de prosocialidade. Também aqui as matrizes simétricas e assimétricas foram normalizadas, da mesma maneira que descrito anteriormente.
Com a finalidade de verificar se a atenção social é um elemento importante para a ocorrência de cooperação e de comportamentos prosociais, para cada teste foi feita uma correlação entre a matriz de olhares sociais e a matriz de puxadas, e a matriz de presença na bandeja foi utilizada como co-variável. Por meio do programa Matrix Tester 2.2.4, foi aplicado o teste Tau Kr parcial, que mede a associação entre duas matrizes mantendo constante o efeito de uma terceira matriz (Hemelrijk, 1990a, b).
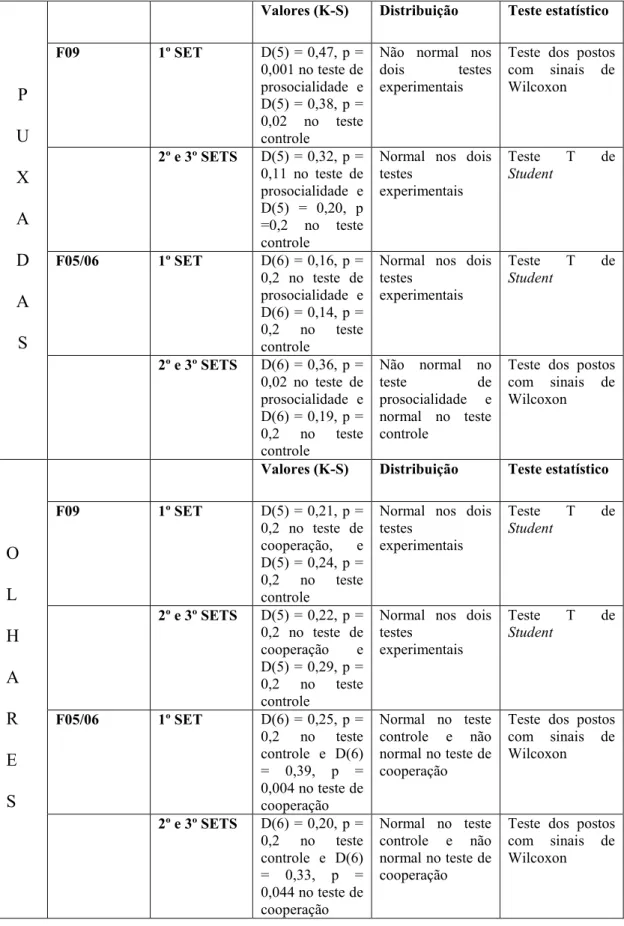
Tabela 2: Tipo de distribuição (normal ou não normal) dos dados de puxadas e olhares sociais nas duas famílias no primeiro e no segundo e terceiro sets e os testes estatísticos utilizados nas análises.
P U X A D A S
Valores (K-S) Distribuição Teste estatístico
F09 1º SET D(5) = 0,47, p = 0,001 no teste de prosocialidade e D(5) = 0,38, p = 0,02 no teste controle
Não normal nos dois testes experimentais
Teste dos postos com sinais de Wilcoxon
2º e 3º SETS D(5) = 0,32, p = 0,11 no teste de prosocialidade e D(5) = 0,20, p =0,2 no teste controle
Normal nos dois testes
experimentais
Teste T de Student
F05/06 1º SET D(6) = 0,16, p = 0,2 no teste de prosocialidade e D(6) = 0,14, p = 0,2 no teste controle
Normal nos dois testes
experimentais
Teste T de Student
2º e 3º SETS D(6) = 0,36, p = 0,02 no teste de prosocialidade e D(6) = 0,19, p = 0,2 no teste controle
Não normal no
teste de
prosocialidade e normal no teste controle
Teste dos postos com sinais de Wilcoxon O L H A R E S
Valores (K-S) Distribuição Teste estatístico
F09 1º SET D(5) = 0,21, p = 0,2 no teste de cooperação, e D(5) = 0,24, p = 0,2 no teste controle
Normal nos dois testes
experimentais
Teste T de Student
2º e 3º SETS D(5) = 0,22, p = 0,2 no teste de cooperação e D(5) = 0,29, p = 0,2 no teste controle
Normal nos dois testes
experimentais
Teste T de Student
F05/06 1º SET D(6) = 0,25, p = 0,2 no teste controle e D(6) = 0,39, p = 0,004 no teste de cooperação
Normal no teste controle e não normal no teste de cooperação
Teste dos postos com sinais de Wilcoxon
2º e 3º SETS D(6) = 0,20, p = 0,2 no teste controle e D(6) = 0,33, p = 0,044 no teste de cooperação
Normal no teste controle e não normal no teste de cooperação
RESULTADOS 1. Relações sociais
Primeiro set
No primeiro set os dados de afiliação foram coletados antes de terem início os testes experimentais, mas não antes do início de cada sessão experimental. Considerando que as relações sociais entre os primatas são consideradas importantes fatores que influenciam a ocorrência de comportamentos cooperativos (Hipóteses 1a e 1b), esperava-se que os animais que mais se catassem ou que passassem mais tempo em proximidade, cooperassem mais nos aparatos (Predição 1a). Ao contrário do esperado, não foi observado correlação positiva significante entre catação e puxadas em nenhuma das famílias (F05/06: Tau Kr = - 0,12, pl =
0,32; F09: Tau Kr = - 0,55, pl = 0,1) nem entre proximidade e puxadas (F05/06: Tau Kr =
0,21, pr = 0,16; F09: Tau Kr = - 0,11, pl = 0,42). No teste de prosocialidade, era esperado que
os animais doassem mais alimento àqueles indivíduos que eles mais catavam e também aos indivíduos com os quais eles passassem mais tempo em proximidade (Predição 1b), mas os resultados não seguiram essas suposições. Na correlação das matrizes de puxadas e catação, na F05/06 o valor do Tau Kr foi - 0, 27 com significância pl = 0,13 e na F09, o Tau Kr não
pôde ser calculado, devido ao grande número de células na matriz com valor zero. Na correlação de puxadas e proximidade, na F05/06 o valor de Tau Kr foi – 0,07 com significância pr = 0,35 e na F09 os resultados foram: Tau Kr = 0,24, pr = 0,51).
A hipótese 1c sugeria que a catação ocorria em um sistema de troca entre os animais e sendo assim, esperava-se que os animais que mais catassem seus parceiros seriam os que mais receberiam alimento desses no teste de prosocialidade (predição 1c). Os resultados mostraram que não ocorreu esse tipo de troca na F05/06 (Tau Kr = - 0,19, pl = 0,23) e na F09 o Tau Kr
Segundo e terceiro sets
A análise dos dados de afiliação que foram registrados por dez dias antes de ter início o segundo set foram correlacionadas com os dados de puxadas do segundo e terceiro sets somados, e também não oferecem suporte à predição 1a, ou seja, não houve correlação significante entre puxadas e catação no teste de cooperação (F05/06: Tau Kr = - 0,15, pl =
0,27; F09: Tau Kr = 0,2, pr = 0,22). Para o teste de prosocialidade também não foi encontrada
correlação como esperava-se com a predição 1b (F05/06: Tau Kr = - 0,2, pl = 0,20; F09: Tau
Kr = 0,85, pr = 0,01). A correlação entre puxadas e proximidade também não foi significante
em nenhum dos testes (Cooperação: F05/06: Tau Kr = 0,15, pr = 0,18; F09: Tau Kr = 0,2, pr =
0,33; Prosocialidade: F05/06: Tau Kr = 0,06, pr = 0,44; F09: Tau Kr = 0,31, pr = 0,23).
Também em desacordo com a hipótese 1c que sugeria que os animais poderiam trocar catação por alimento, a correlação de matrizes não foi significante na F09 (Tau Kr = - 0,22, pl
= 0,23) nem na F05/06 (Tau Kr = 0,05, pr = 0,39).
As correlações de matrizes com os dados de relações afiliativas que foram coletados diariamente, antes do início de cada sessão experimental também vão de encontro às nossas hipóteses iniciais. A correlação entre puxadas e catação não foi significante no teste de cooperação em nenhum das famílias (F05/06: Tau Kr = - 0,25, pl = 0,09; F09: Tau Kr = 0,15,
pr = 0,36), nem no teste de prosocialidade (F05/06: Tau Kr = - 0,15, pl = 0,19; F09: Tau Kr = -
0,55, pl = 0,09). As correlações entre puxadas e proximidade também não mostraram
significância estatística no teste de cooperação (F05/06: Tau Kr = - 0,07, pl = 0,42; F09: Tau
Kr = 0,14, pr = 0,37), e no teste de prosocialidade não houve significância na F09 (Tau Kr =
0,18, pr = 0,37) mas houve na F05/06, porém, com correlação negativa, indo de encontro ao
que foi previsto (Tau Kr = - 0,42, pl = 0,02). Sobre a hipótese 1c, também não foi encontrada
2. Atenção social
Primeiro set
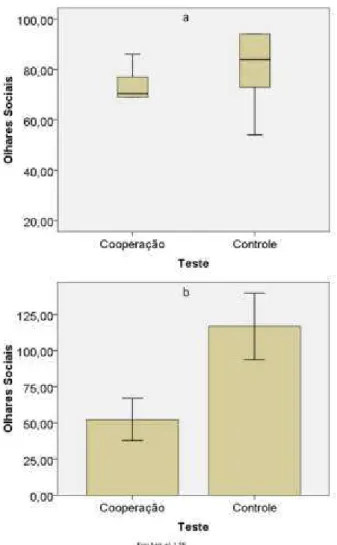
De acordo com a hipótese da cooperação verdadeira (Hipótese 2a), era esperado que em testes em que era necessária a ação simultânea de dois indivíduos ocorresse maior troca de olhares entre os animais do que em testes onde um indivíduo sozinho poderia resolver a tarefa (Predição2a´). No entanto, os resultados foram opostos a essa expectativa e a quantidade de olhares sociais por indivíduo foi maior no teste controle (figura 5). Os resultados evidenciaram diferença significante na F09 (média = 52,4, EP = 14,63 no teste de cooperação; média = 116,8, EP = 22,91 no teste controle; t (4) = -5,77, p = 0,002, r = 0,94) e na F05/06 (mediana = 70,5 teste de cooperação e mediana = 84 no controle, T = 0, p = 0,01, r = - 0,64).
Indo de encontro à predição 2a que previa atenção social para comunicação e coordenação de atividades no teste de cooperação, ou seja, uma correlação positiva entre a execução das puxadas no aparato e os olhares sociais, o teste de correlação de matrizes não mostrou significância na família F05/06 (Tau Kr parcial = 0,39; pr = 0,06) nem na família F09
(Tau Kr parcial = 0,12; pr = 0,43).
Se os indivíduos apresentassem comportamento prosocial verdadeiro, ou seja, mediado por atenção social (Hipótese 2b), a correlação entre puxadas e olhares sociais no teste de prosocialidade deveria ser positiva (Predição 2b), mas a análise não mostrou significância em nenhuma das famílias (F05/06: Tau Kr parcial = 0,04; pr = 0,40; F09: Tau Kr
Figura 5: Número de olhares sociais por indivíduo nos testes de cooperação e controle no primeiro set. a. Mediana e distância interquartílica na F05/06; b. Média e erro padrão na F09.
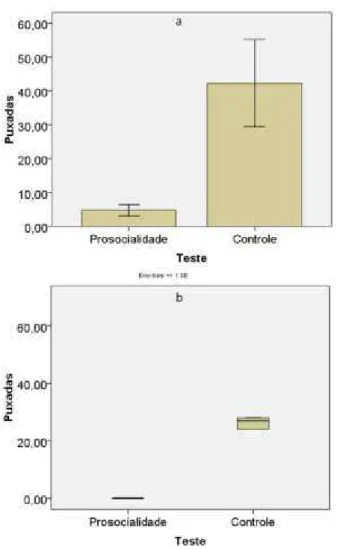
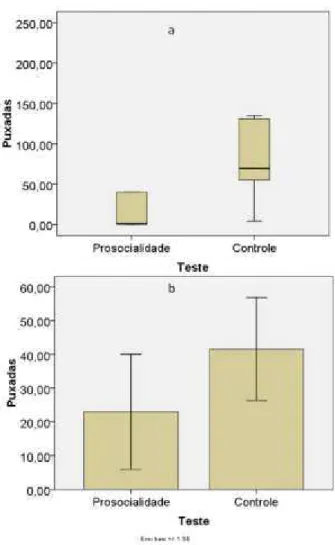
Figura 6: Número de puxadas por indivíduo nos testes de prosocialidade e controle no primeiro set. a. Média e erro padrão na F05/06; b. Mediana e distância interquartílica na F09.
Segundo e terceiro sets
Assim como no primeiro set, a matriz de olhares sociais não mostrou correlação significativa com a matriz de puxadas no teste de cooperação (F05/06: Tau Kr parcial = 0,004, pr = 0,53; F09: Tau Kr parcial = 0,29, pr = 0,19) nem no teste de prosocialidade
(F05/06: Tau Kr parcial = 0,43, pr = 0,06; F09: Tau Kr parcial = 0,23, pr = 0,22).
28,45 no teste de cooperação; média = 172,8, EP = 42,38 no controle; t(4) = - 0,27, p = 0,40, r = 0,14).
Figura 7: Número de olhares sociais por indivíduo nos testes de cooperação e controle no segundo e terceiro sets. a. Mediana e distância interquartílica na F05/06; b. Média e erro padrão na F09.
DISCUSSÃO 1. Relações sociais
A literatura apresenta dados para algumas espécies de calitriquídeos que mostram que durante a realização de atividades cooperativas esses animais não discriminam parentes e não parentes nem parceiros reprodutivos e não parceiros (Burkart et al. 2007; Cronin et al. 2009; Stevens 2010). Não há, entretanto, dados relacionando fatores sociais com prosocialidade ou cooperação em saguis. O presente trabalho testou se comportamentos afiliativos poderiam influenciar a ocorrência de comportamentos cooperativos nesses animais, e os resultados foram negativos. Não houve correlação entre a execução das tarefas cooperativas com a catação ou a proximidade, nem em dados sociais coletados por dez dias antes dos experimentos, nem em dados coletados diariamente, imediatamente antes de cada teste.
Sugere-se que as altas taxas de interações afiliativas que normalmente ocorrem no contexto familiar dos sagüis podem ter reduzido a possibilidade de seu comportamento durante as tarefas depender de interações de afiliação passadas. Com uma organização social típica dos calitriquídeos, os sagüis são indivíduos altamente tolerantes, e a catação pode ocupar mais de 30 por cento da fase ativa desses animais (Azevedo et al. 1997). Pode ser que relações positivas não tenham efeito sobre a escolha de parceiros cooperativos e outros fatores como a idade (como sugere o comportamento de um indivíduo chamado Onésio, descrito a seguir), a hierarquia ou o sexo podem ser mais determinantes. Uma análise post hoc do trabalho de Burkart et al. (2007) permitiu identificar, por exemplo, que a prosocialidade encontrada nos animais na análise agrupada não existiu entre as fêmeas.
indivíduo na bandeja ele o afastava e monopolizava a bandeja contendo a banana, e isso ocorria para todos os indivíduos com exceção de seu irmão mais velho, o Opus. Ao Opus era permitido pegar a recompensa e ao contrário do que ocorria com os outros animais, os quais eram afastados com mordidas, empurrões e outras agressões, poucos comportamentos agonísticos foram direcionados a ele (observações pessoais). Observa-se, portanto, que o comportamento competitivo foi predominantemente mais notável que o comportamento prosocial. Este último ocorreu somente em benefício de seu irmão mais velho o que pode revelar que existe um tipo de parceiro preferencial para a prosocialidade que em nossa amostra estaria sub-representado.
Outra possível explicação para a ausência de correlação entre os comportamentos afiliativos e a execução das tarefas, é a condição experimental, com todos os animais juntos no viveiro. Segundo Burkart & van Schaik (2012) vários animais presentes ao redor de um aparato experimental torna improvável a possibilidade dos animais escolherem parceiros para a realização de tarefas, e as puxadas podem ocorrer sem o controle de quem está na outra bandeja, não havendo formação de duplas preferenciais para a ocorrência de comportamentos cooperativos.
catação e a proximidade com os comportamentos cooperativos em Callithrix jacchus, e os resultados foram negativos, não tendo sido observada maior tendência a comportamentos cooperativos entre indivíduos com maiores taxas de comportamentos afiliativos, nem troca de catação por cooperação ou prosocialidade. Entretanto, outras análises precisam ser feitas para um melhor entendimento sobre o assunto.
A realização dos experimentos com os animais separados em duplas, organizados em todas as possibilidades de díades, permitiria uma comparação entre duas condições experimentais, e ainda, um estudo de longo prazo pode ser mais eficiente para identificar as relações entre os animais. Isso contribuiria para um melhor entendimento sobre a influência das relações sociais sobre os comportamentos cooperativos dessa espécie em cativeiro.
2. Atenção social
Muitos trabalhos já identificaram que espécies de primatas do Velho Mundo são capazes de entender o papel de outros indivíduos na realização de tarefas de cooperação (Chalmeau 1994; Chalmeau & Gallo 1996a, Chalmeau & Gallo 1996b; Chalmeau et al. 1997a; Hirata & Fuwa 2007) e verifica-se uma crescente discussão sobre se a cooperação entre os primatas do Novo Mundo seria uma cooperação acidental, ou se os indivíduos envolvidos também levam em conta o papel do parceiro na realização de uma tarefa em conjunto (Chalmeau et al. 1997b; Mendres & de Waal 2000; Visalberghi et al. 2000; Hattori et al. 2005; Cronin et al. 2005; Mendes 2009).
Os calitriquídeos são animais que têm se destacado em alguns estudos que testam suas habilidades cognitivas, principalmente no domínio social. Seu desempenho em tarefas cooperativas e de aprendizado social sustenta a idéia de que eles são atenciosos aos companheiros de grupo, são capazes de coordenar ações no tempo e no espaço para cooperar e fornecem alimento a co-específicos em diferentes situações.
Eles já se mostraram capazes de aprender (Snowdon 2001), cooperar (Werdenich & Huber 2002; Cronin et al. 2005), e prover alimento de maneira prosocial (Burkart et al. 2007; Cronin et al. 2010; Brurkart & van Schaik 2012). Algumas dessas características são semelhantes às que ocorrem nos grandes primatas e têm sido atribuídas à cognição social diferencial desses pequenos antropóides que deve ter relação com o sistema de cuidado cooperativo dos filhotes (Burkart et al. 2009; Burkart & van Schaik 2010).
saguis estudados puderam realizar atividades de cooperação em um nível elaborado com divisão de tarefas complementares (Werdenich & Huber 2002). Em ambos os trabalhos, entretanto, as condições experimentais eram diferentes das do presente estudo, o que torna a comparação delicada, como discutido adiante.
Com relação ao teste de prosocialidade, os resultados não corroboram a nossa predição de que as díades com maiores quantidades de olhares sociais apresentam também maiores taxas de comportamentos prosociais, ou seja, maiores números de puxadas para o parceiro receber a recompensa. Para Chalmeau & Gallo (1996a) os olhares entre os indivíduos são um importante indicador sobre o que os sujeitos aprendem numa situação social. Baseado nessa idéia, não se pode inferir aqui que os indivíduos estavam atentos aos seus parceiros e que essa atenção determinou a ocorrência de comportamento prosocial entre os saguis.
Sugerem-se duas possíveis explicações para esse resultado: 1) não houve entendimento da tarefa, e os indivíduos não estavam atentos, não compreendendo também o papel de receptor do parceiro. Isso não excluiria a existência de prosocialidade nos saguis estudados, o que estaria de acordo com a literatura sobre o assunto em Callithrix jacchus (Burkart et al. 2007; Burkart & van Schaik 2012), visto que a não ocorrência da prosocialidade seria devido à não compreensão do experimento e não à falta de tendências prosociais; 2) houve entendimento da tarefa, os indivíduos estavam atentos aos seus parceiros, porém, a atenção pode estar menos relacionada com a prosocialidade, e mais com competição. Assim, ao entender que seus parceiros seriam beneficiados a seu custo, os animais escolheram por agir de forma não prosocial.
que houve entendimento da tarefa, além de mostrar que a competição pode ter tido papel importante durante sua execução.
De acordo com a segunda explicação está o trabalho de Stevens (2010) com outro calitriquídeo, o tamarin, utilizando aparatos experimentais. O autor verificou que os animais sempre puxavam as alavancas quando recebiam alguma recompensa, mas raramente o faziam quando não recebiam, e na presença de um parceiro, os animais ativos puxavam muito menos a alavanca quando essa ação não resultava em recompensa para eles mesmos mas somente para o parceiro. Entretanto, comportamentos agonísticos não foram relatados, uma vez que os animais estudados estavam separados por uma grade.
O trabalho de Cronin et al. (2009) também com tamarins, está igualmente em acordo com o presente estudo. Em seu trabalho os animais tiveram a opção de escolher entre duas opções de bandeja (0,0 ou 0,1), e não demonstraram preferência pela bandeja com a opção prosocial (0,1) na presença de um parceiro, demonstrando baixa motivação para puxar a alavanca do aparato quando não obtinham recompensa pelo seu esforço. Os autores ainda observaram que os animais puxavam a opção prosocial menos ainda se o parceiro demonstrasse algum sinal de interesse pelo petisco (Cronin et al. 2009), sugerindo que há competição entre os animais.
Em vista disso, sobre as explicações aventadas acima, sugere-se que pode ter havido compreensão da tarefa, com ocorrência de atenção social, mas esta estava voltada para a competição. A competição no contexto alimentar já foi reportada em Callithrix jacchus (Tardif & Richter 1981; Lopes 2002; Queiroz 2003, citado por Menezes 2010), e neste trabalho, também foram registrados alguns eventos de roubo e agonismo durante os experimentos.
isolados de seus grupos familiares, em pequenas gaiolas, o que faz com que os animais encontrem poucas opções comportamentais. Somente um estudo fez experimentos com os animais todos em grupo no mesmo recinto (Burkart & van Schaik 2012). Apesar desse estudo não ter identificado diferença no desempenho dos animais quando em grupo ou quando separados em duplas (Burkart et al. 2007), a literatura sugere que diferentes condições experimentais podem gerar resultados variados. Neste trabalho os indivíduos estavam todos juntos no viveiro onde foram realizadas as tarefas, e duplas de parceiros foram formadas espontaneamente sem categorização de sexo, idade ou status social. Durante os experimentos alguns indivíduos brincavam, outros se catavam, enquanto alguns participavam das tarefas. Nessas condições os animais tendem a expressar comportamentos diferentes do que quando estão isolados. No estudo de Chalmeau et al. (1997b), por exemplo, os autores trabalharam com Cebus apella nas mesmas condições que o presente trabalho, e seus resultados para esses animais foram semelhantes aos aqui obtidos. Por outro lado, o trabalho também com macacos prego, mas com os indivíduos confinados a pequenas gaiolas, obteve resultados opostos, sustentando que os macacos prego utilizam-se dos olhares sociais para se comunicar com o parceiro e são capazes de entender o papel deste na realização de uma tarefa cooperativa (Mendres & de Waal 2000). Halsey et al. (2006) compararam o desempenho, em Callithrix jacchus, no teste de cordas paralelas em dois ambientes, e encontraram diferenças no
desempenho dos animais em cativeiro e em ambiente natural. Para os autores essa diferença se deve a dificuldades impostas na natureza (como a atenção a predadores) e que não ocorrem em cativeiro. A maneira como os animais se encontram durante um experimento pode, portanto, ter forte influência sobre os resultados obtidos.
CONCLUSÃO
Os comportamentos cooperativos constituem um assunto controverso e apesar do crescente número de trabalhos a esse respeito, muito ainda é necessário se estudar para um melhor conhecimento sobre o tema. Os estudos sobre esse assunto em primatas não humanos têm como principal objetivo esclarecer sobre esses comportamentos nos humanos e para isso, há muitos trabalhos com diferentes focos de análise. Pensando nos mecanismos (causas próximas) que sustentam os comportamentos cooperativos, as relações entre os indivíduos e a atenção subjacente a esses comportamentos estão entre os focos mais explorados, e os primatas do Velho Mundo sempre foram considerados mais interessantes para serem estudados.
REFERÊNCIAS BIBLIOGRÁFICAS
Ahumada, J. A. 1992. Grooming behavior of spider monkeys (Ateles geoffroyi) on Barro Colorado Island, Panama. International Journal of Primatology, 13, 33–49.
Altmann, J. 1974. Observational study of behavior: sampling methods. Behaviour, 49, 227-267.
Anderson, J. R. 2007, Animal behavior: tolerant primates cooperate best. Current Biology, 17, 7, 242-244.
Araújo, A., Arruda, M. F., Alencar, A. I., Albuquerque, F., Nascimento, M. C. & Yamamoto, M. E. 2000. Body weight of wild and captive common marmosets (Callithrix jacchus). International Journal of Primatology, 21, 2, 317-324.
Azevedo, C. V. M., Braga, K. R. G, Silva, L. C., Silva, R. P., Menezes, A. A. L. & Moreira, L. F. S. 1997. Influência de fatores sociais no ritmo circadiano de catação do sagüi comum (Callithrix jacchus) em cativeiro. In: A Primatologia no Brasil 5 (Org. por S. F. Ferrari, H. Schneider), pp. 269-277. Belém: Editora Universitária - UFPA.
Barrett, L., Henzi, S. P., Weingrill, T., Lycett, J. E. & Hill, R. A. 2012. Market forces predict grooming reciprocity in female baboons. Proceedings of the Royal Society B: Biological Sciences, 266, 665-670.
Boesch, C. & Boesch, H. 1989. Hunting behavior of wild chimpanzees in the Taï National Park. American Journal of Physical Anthropology, 78, 547-573.
Burkart, J. M. 2009. Socio-cognitive abilities and cooperative breeding. In: Learning from Animals? Examining the Nature of Human Uniqueness (Org. por L. S. Röska-Hardy &
E. M. Neumann-Held), pp. 123-141. New York: Psychology Press.
Burkart, J. M., Fehr, E., Efferson, C. & van Schaik, C. P. 2007. Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. PNAS, 50, 19762–19766.
Burkart, J. M., Hrdy, S.B. & van Schaik, C. P. 2009. Cooperative beeding and human cognitive evolution. Evolutionary Anthropology, 18, 175-186.
Burkart, J. M., Kupferberg, A., Glasauer, S. & van Schaik, C. P. 2011. Even Simple forms of social learning rely on intention attribution in marmoset monkeys (Callithrix jacchus). Journal of Comparative Psychology, 1-10.
Burkart, J.M. & van Schaik, C. P. 2010. Cognitive consequences of cooperative breeding in primates? Animal Cognition, 13, 1-19.
Burkart, J. M. & van Schaik, C. P. 2012. Group service in macaques (Macaca fuscata), capuchins (Cebus apella) and marmosets (Callithrix jacchus): a comparative approach to identifying proactive prosocial motivations. Journal of Comparative Psychology, 1-14.
Byrne, R. W. & Whiten, A. 1988. Machiavellian Intelligence: Social Expertise and the Evolution of Intellect in Monkeys, Apes and Humans. Oxford: Clarendon Press.
Cacchione, T. & Burkart, J. M. 2012. Dissociation between seeing and acting: insights from common marmosets (Callithrix jacchus). Behavioural Processes, 89, 52-60.
Caldwell, C. A. & Whiten, A. 2003. Scrounging facilitates social learning in common marmosets, Callithrix jacchus. Animal Behaviour, 65, 1085–1092.
Caldwell, C. A., Whiten, A. & Morris, K. D. 1999. Observational learning in marmoset monkey (Callithrix jacchus). In: AISB convention: symposium on imitation in animals and artifacts (Ed. por The Society for the Study of Artificial Intelligence and the
Simulation of Behaviour), (pp. 27-31), Edinburgh.
Chalmeau, R. 1994. Do chimpanzees cooperate in a learning task? Primates, 35, 385-392. Chalmeau, R. & Gallo, A. 1996a. What Chimpanzees (Pan troglodytes) learn in a
Cooperative Task. Primates, 37, 39-47.
Chalmeau, R. & Gallo, A. 1996b. Cooperation in primates: critical analysis of behavioural criteria. Behavioural Processes, 35, 101-111.
Chalmeau, R., Lardeux, K., Brandibas, P. & Gallo, A. 1997a. Cooperative Problem Solving by Orangutans (Pongo pygmaeus). International Journal of Primatology, 18, 23-32.
Chalmeau, R., Visalberghi, E. & Gallo, A. 1997b. Capuchin monkeys, Cebus apella, fail to understand a cooperative task. Animal Behaviour, 54, 1215-1225.
Cheney, D. L., Moscovice, L. R., Heesen, M., Mundry, R. & Seyfarth, R. M. 2010. Contingent cooperation between wild female baboons. PNAS, 107, 21, 9562-9566. Cortés-Ortiz, L. 2009. Molecular phylogenetics of the Callitrichidae with an emphasis on the
marmosets and Callimico. In: The Smallest Anthropoids, The Marmoset/Callimico Radiation (Org. por M. Ford, L. M. Porter & L. C. Davis), pp. 3-24, USA: Springer
Press.
Crofoot, M. C., Rubenstein, D. I., Maiya, A. S. & Berger-Wolf, T. 2011. Aggression, grooming and group-level cooperation in white-faced capuchins (Cebus capucinus): insights from social networks. American Journal of Primatology, 73, 821-833.