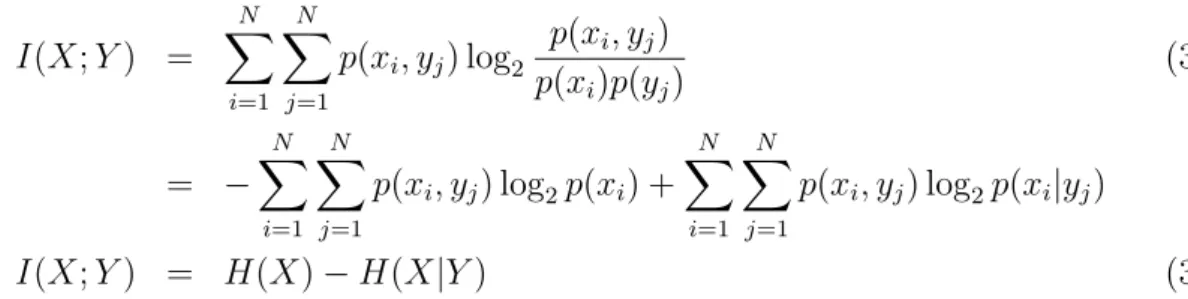Universidade Federal do Rio Grande do Norte
Instituto do C´erebro
Curso de Mestrado em Neurociˆencias
Disserta¸c˜
ao de Mestrado
Medida de uma Codifica¸c˜ao de Est´ımulos Sonoros por Potenciais
P´os-Sin´apticos e Potenciais de Campo Local Usando Teoria da
Informa¸c˜ao
Mestranda
Juliana Martins de Assis
Orientador
Adriano Bretanha Lopes Tort, Dr.
Juliana Martins de Assis
Medida de uma Codifica¸c˜ao de Est´ımulos Sonoros por Potenciais
P´os-Sin´apticos e Potenciais de Campo Local Usando Teoria da
Informa¸c˜ao
Disserta¸c˜ao apresentada ao Programa de P´os-Gradua¸c˜ao em Neurociˆencias da Uni-versidade Federal do Rio Grande do Norte como pr´e-requisito para obten¸c˜ao do t´ıtulo de Mestre em Neurociˆencias.
´
Area de concentra¸c˜ao: Neurociˆencias.
Linha de pesquisa: processamento de sinais aplicado `as Neurociˆencias.
Dr. Adriano Bretanha Lopes Tort (Orientador)
Juliana Martins de Assis
Medida de uma Codifica¸c˜
ao de Est´ımulos Sonoros por
Potenciais P´
os-Sin´
apticos e Potenciais de Campo Local Usando
Teoria da Informa¸c˜
ao
Apresentada em 3 de Julho de 2014
Banca Examinadora:
Prof. Dr. Jean Faber Ferreira de Abreu
Instituto de Ciˆencia e Tecnologia - Engenharia Biom´edica Universidade Federal de S˜ao Paulo
Profa
. Dra
. Kerstin Erika Schmidt Instituto do C´erebro
Universidade Federal do Rio Grande do Norte
Prof. Dr. Adriano Bretanha Lopes Tort Instituto do C´erebro
Universidade Federal do Rio Grande do Norte
Resumo
O processo de codifica¸c˜ao ´e um aspecto fundamental do funcionamento cerebral. A transforma¸c˜ao de est´ımulos sensoriais em respostas neurofisiol´ogicas tem sido objeto de estudo em diversas ´areas da Neurociˆencia. Um dos modos mais utilizados para medir a eficiˆencia de um c´odigo neural ´e pelo uso de medidas advindas da Teoria da Informa¸c˜ao,
como a informa¸c˜ao m´utua. Utilizando estas ferramentas, estudos recentes mostraram
que no c´ortex auditivo tanto registros de potenciais de campo local (LFPs - local field
potentials) quanto os tempos de disparos de potenciais de a¸c˜ao codificam informa¸c˜ao
sobre est´ımulos sonoros. Todavia, n˜ao h´a estudos aplicando ferramentas da Teoria da Informa¸c˜ao para investigar a eficiˆencia de c´odigos que utilizem potenciais p´os-sin´apticos
(PSPs - postsynaptics potentials), isoladamente e em conjunto com an´alises de LFPs.
Estes sinais est˜ao relacionados visto que os LFPs s˜ao em parte constru´ıdos pela a¸c˜ao
conjunta de v´arios PSPs. A presente disserta¸c˜ao reporta medidas de informa¸c˜ao m´utua
entre respostas de PSPs e LFPs obtidas no c´ortex auditivo prim´ario de ratos anestesiados e est´ımulos sonoros de frequˆencias distintas. Nossos resultados mostram que respostas de PSPs possuem informa¸c˜ao sobre est´ımulos sonoros, em n´ıveis compar´aveis e mesmo maiores que as respostas de LFPs. Tamb´em encontramos que PSPs e LFPs codificam informa¸c˜ao sonora independentemente, uma vez que a an´alise conjunta destes sinais n˜ao mostrou sinergia nem redundˆancia.
Palavras-chave. Codifica¸c˜ao, Potencial P´os-sin´aptico, Potencial de Campo Local,
Abstract
Coding process is a fundamental aspect of cerebral functioning. The sensory stimuli transformation in neurophysiological responses has been a research theme in several areas of Neuroscience. One of the most used ways to measure a neural code efficiency is by the use of Information Theory measures, such as mutual information. Using these tools, recent studies show that in the auditory cortex both local field potentials (LFPs) and action potential spiking times code information about sound stimuli. However, there are no studies applying Information Theory tools to investigate the efficiency of codes that use postsynaptics potentials (PSPs), alone and associated with LFP analysis. These signals are related in the sense that LFPs are partly created by joint action of several PSPs. The present dissertation reports information measures between PSP and LFP responses obtained in the primary auditory cortex of anaesthetized rats and auditory stimuli of distinct frequencies. Our results show that PSP responses hold information about sound stimuli in comparable levels and even greater than LFP responses. We have also found that PSPs and LFPs code sound information independently, since the joint analysis of these signals did neither show synergy nor redundancy.
Agradecimentos
Agrade¸co em primeiro lugar aos meus pais, Aida Maria e Francisco Marcos, pelo apoio incondicional, n˜ao s´o durante o Mestrado como nas fases que lhe precedem, sem as quais n˜ao haveria chegado at´e aqui. Agrade¸co tamb´em aos meus irm˜aos, Juana Tereza e Joa-quim Marcos, que mesmo `a distˆancia tem sido muito efetivos em me motivar. Agrade¸co aos meus tios Gotardo, Evanceli, e `as primas Gabi e Carol, que t˜ao bem me acolheram em Natal, por um bom tempo. Meu agradecimento sincero tamb´em a toda a turma da Capoeira Brasil pelas excelentes vivˆencias neste tempo de Mestrado (ax´e!).
Agrade¸co tamb´em a todos os colegas e funcion´arios do Instituto do C´erebro, que me receberam muito bem e com os quais estabeleci v´ınculos amistosos. Agrade¸co especial-mente a Rodrigo Pav˜ao pelas ideias sobre aplica¸c˜ao de Teoria da Informa¸c˜ao, a Bryan Souza e a V´ıtor Santos, pela ajuda em alguns programas, como tamb´em a Camile Cˆorrea, Kelly Sores e tantos outros pelo companheirismo. Agrade¸co a Ana Maria, que desde o come¸co apontou para a possibilidade de fazer o mestrado no Instituto. Agrade¸co enfim, e n˜ao menos importantemente, ao professor Adriano Tort, pela paciˆencia com que me conduziu neste trabalho e por sua confian¸ca em mim.
Ep´ıgrafe
“carne alma
forma conte´udo
Sum´
ario
1 Introdu¸c˜ao 1
1.1 Potenciais P´os-Sin´apticos . . . 2
1.2 Potenciais de Campo Local . . . 4
1.3 C´ortex Auditivo . . . 5
1.4 Objetivos . . . 7
1.5 Estrutura do Trabalho . . . 7
2 Coleta dos dados 8 3 Conceitos de Teoria da Informa¸c˜ao 10 4 M´etodos de corre¸c˜ao de vi´es 15 4.1 M´etodoShuffling . . . 17
4.2 M´etodoPanzeri-Treves (PT) . . . 17
4.3 M´etodo da extrapola¸c˜ao quadr´atica (QE) . . . 18
4.4 Simula¸c˜oes comparativas entre os m´etodos . . . 19
5 An´alise dos dados 21 5.1 Suaviza¸c˜ao das curvas . . . 21
5.2 Retirada de Amostras Discrepantes . . . 23
5.3 N´umero de Amostras N˜ao Discrepantes . . . 24
5.4 Obten¸c˜ao de Amplitudes e Latˆencias . . . 29
6 Resultados 30 6.1 Caracteriza¸c˜ao das amplitudes e latˆencias de PSPs e LFPs . . . 30
6.2 An´alise separada da informa¸c˜ao sensorial por PSPs ou LFPs . . . 39
6.3 Compara¸c˜ao entre informa¸c˜oes sensoriais por PSPs e por LFPs . . . 43
6.4 Codifica¸c˜oes conjuntas de PSPs e LFPs . . . 45
6.5 Influˆencia da anestesia nas codifica¸c˜oes . . . 46
7 Conclus˜ao e Considera¸c˜oes Finais 49 7.1 Conclus˜oes . . . 49
7.2 Considera¸c˜oes finais . . . 50
A Tabela de anestesia por c´elula 57
B Observa¸c˜ao dos Registros 58
Lista de Figuras
1.1 Esquema ilustrativo de sinapse excitat´oria ionotr´opica . . . 3
1.2 Exemplo de PSP . . . 4
1.3 Exemplo de LFP . . . 5
1.4 N´ucleos de retransmiss˜ao do sistema auditivo . . . 6
2.1 Exemplo de aparelho de registro . . . 9
3.1 Diagrama para entropia . . . 12
4.1 Exemplo de emprego do estimador naive . . . 16
4.2 Exemplo de extrapola¸c˜ao quadr´atica para estima¸c˜ao de entropia . . . 19
4.3 Compara¸c˜ao entre os m´etodos de corre¸c˜ao de vi´es . . . 20
5.1 Exemplos de registros . . . 22
5.2 Filtragem de registros . . . 23
5.3 Sinais discrepantes . . . 24
5.4 N´umero de amostras de PSPs . . . 25
5.5 N´umeros de amostras de LFPs . . . 26
5.6 N´umeros de amostras de PSPs e LFPs . . . 27
5.7 Erro percentual m´edio e n´umero de amostras . . . 28
5.8 Obten¸c˜ao de amplitudes e latˆencias . . . 29
6.1 Exemplos de curvas de caracteriza¸c˜ao de aamplitudes . . . 31
6.2 Curvas de caracteriza¸c˜ao de amplitudes normalizadas para todas as c´elulas 32 6.3 Distribui¸c˜ao de est´ımulos de maior amplitude . . . 33
6.4 Exemplos de curvas de latˆencias . . . 34
6.5 Curvas normalizadas para todas as c´elulas (latˆencias) . . . 35
6.6 Distribui¸c˜ao de est´ımulos de menor latˆencia . . . 36
6.7 Scatter plots de m´edias de amplitudes e latˆencias de PSPs, e m´edias de amplitudes e latˆencias de LFPs e correla¸c˜oes de Pearson. . . 37
6.8 Scatter plots de m´edias de amplitudes de PSPs e latˆencias de LFPs, e m´edias de latˆencias de PSPs e amplitudes LFPs de cada c´elula e correla¸c˜oes de Pearson. . . 38
6.9 Scatter plotsde m´edias de amplitudes de PSPs e LFPs e latˆencias de PSPs e LFPs de cada c´elula e correla¸c˜oes de Pearson. . . 39
6.10 Compara¸c˜ao entre informa¸c˜ao de amplitudes e latˆencias . . . 40
6.11 Compara¸c˜ao entre informa¸c˜ao atrav´es de amplitudes ou latˆencias, conjunta ou isoladamente . . . 41
6.14 Compara¸c˜ao entre informa¸c˜ao conjunta de PSPs e de LFPs . . . 45
6.15 Independˆencia entre registros de PSP e LFP . . . 46
6.16 Influˆencia da anestesia na informa¸c˜ao . . . 48
B.1 PSPs de c´elulas 1 e 2 . . . 59
B.2 PSPs de c´elulas 3 e 4 . . . 60
B.3 PSPs de c´elulas 5 e 6 . . . 61
B.4 PSPs de c´elulas 7 e 8 . . . 62
B.5 PSPs de c´elulas 9 e 10 . . . 63
B.6 PSPs de c´elulas 11 e 12 . . . 64
B.7 PSPs de c´elulas 13 e 14 . . . 65
B.8 PSPs de c´elulas 15 e 16 . . . 66
B.9 PSPs de c´elulas 17 e 18 . . . 67
B.10 PSPs de c´elulas 19 e 20 . . . 68
B.11 PSPs de c´elulas 21 e 22 . . . 69
B.12 PSPs de c´elulas 23 e 24 . . . 70
B.13 PSPs de c´elulas 25 e 26 . . . 71
B.14 PSPs de c´elulas 27 e 28 . . . 72
B.15 PSPs de c´elulas 29 e 30 . . . 73
B.16 PSPs de c´elulas 31 e 32 . . . 74
B.17 PSPs da c´elulas 33 e LFPs da c´elula 1 . . . 75
B.18 LFPs das c´elulas 2 e 3 . . . 76
B.19 LFPs de c´elulas 4 e 5 . . . 77
B.20 LFPs de c´elulas 6 e 7 . . . 78
B.21 LFPs de c´elulas 8 e 9 . . . 79
B.22 LFPs de c´elulas 10 e 11 . . . 80
B.23 LFPs de c´elulas 12 e 13 . . . 81
B.24 LFPs de c´elulas 14 e 15 . . . 82
B.25 LFPs de c´elulas 16 e 17 . . . 83
B.26 LFPs de c´elulas 18 e 19 . . . 84
B.27 LFPs de c´elulas 20 e 21 . . . 85
B.28 LFPs de c´elulas 22 e 23 . . . 86
B.29 LFPs de c´elulas 24 e 25 . . . 87
B.30 LFPs de c´elulas 26 e 27 . . . 88
Cap´ıtulo 1
Introdu¸c˜
ao
A ideia vigente em Neurociˆencias ´e de que o c´erebro ´e um ´org˜ao de grande capacidade computacional, que constr´oi as experiˆencias sens´orias, regula pensamentos, emo¸c˜oes e atitudes [1]. No entanto, n˜ao se sabe ao certo como essa computa¸c˜ao ´e realizada e diversos experimentos tˆem sido feitos na busca por poss´ıveis codifica¸c˜oes e decodifica¸c˜oes. O termo codifica¸c˜ao ´e atribu´ıdo ao processo de transforma¸c˜ao de est´ımulos sens´orios em respostas neurais, observ´aveis experimentalmente. J´a a decodifica¸c˜ao consiste em utilizar as mesmas respostas neurais para reconstruir qual foi o est´ımulo apresentado ou qual atividade motora ou comportamental a ser feita pelo animal [2].
A busca por codifica¸c˜oes neurais tem sido feita nos c´ortices sensoriais de animais e humanos, tais quais o visual, o auditivo e o somatosensorial [3] [4] [5] [6]. O m´etodo mais comum de se estudar o processamento sensorial da informa¸c˜ao pelo c´erebro ´e atrav´es da estimula¸c˜ao repetida, atrav´es dos sinais f´ısicos apropriados, do receptor sensorial; e o subsequente estudo da ´area cortical implicada [7]. O estudo da ´area cortical implicada geralmente consiste na an´alise das respostas neurais registradas eletrofisiologicamente, as
quais podem ser invasivas (an´alise por spikes, por exemplo) ou n˜ao (registros
eletroence-falogr´aficos - EEGs).
Os potenciais de a¸c˜ao, spikes ou “disparos”, s˜ao os sinais mais utilizados na pesquisa
por c´odigos neurais, sendo os elementos constituintes em c´odigos temporais ou por taxa
[8]. Os c´odigos por taxa contam o n´umero despikes em v´arias amostras ou em um
inter-valo de tempo e fazem uma m´edia de sua ocorrˆencia, ao passo que os c´odigos temporais
observam a rela¸c˜ao precisa de tempo entre spikes individuais em neurˆonios distribu´ıdos
[9].
Medidas de associa¸c˜ao entre vari´aveis ambientais e neurofisiol´ogicas podem ser fei-tas para compreender quais sinais neurofisiol´ogicos, ou quais de seus aspectos, possuem maior relevˆancia para a codifica¸c˜ao interna. Como geralmente se observa grande vari-abilidade nas respostas neurofisiol´ogicas e nas vari´aveis externas `as quais os seres vivos
est˜ao submetidos, s˜ao usadas medidas como correla¸c˜ao e informa¸c˜ao m´utua, adequadas
no tratamento de vari´aveis aleat´orias [10]. Esta ´ultima apresenta a vantagem de capturar
dependˆencias n˜ao lineares entre as vari´aveis aleat´orias, o que n˜ao ´e verdadeiro para a correla¸c˜ao de Pearson.
´
1.1
Potenciais P´
os-Sin´
apticos
Os neurˆonios s˜ao c´elulas que geralmente apresentam uma diferen¸ca de potencial negativa em sua membrana, em rela¸c˜ao ao meio extracelular, chamada de potencial de repouso. Esse fato se deve essencialmente `a presen¸ca de canais que permitem a entrada e a sa´ıda de ´ıons da c´elula de maneira diferenciada e devido `as diferentes concentra¸c˜oes desses ´ıons nos meios intra e extracelulares.
´
E bem estabelecido em Neurociˆencias que a principal forma de comunica¸c˜ao entre neurˆonios ´e a sinapse (h´a pesquisas, por´em, de outras formas poss´ıveis [11]). A sinapse pode ser el´etrica ou qu´ımica. As sinapses el´etricas s˜ao mais r´apidas, por possu´ırem pouco ou nenhum espa¸co entre os neurˆonios envolvidos, est˜ao presentes no hipot´alamo, no cerebelo e no c´ortex cerebral, permitindo atividades de sincroniza¸c˜ao da respira¸c˜ao, por exemplo. Por sua vez, a sinapse qu´ımica apresenta uma fenda sin´aptica atrav´es da qual a sinaliza¸c˜ao qu´ımica ´e feita, sendo mais lenta e representando a maioria das sinapses.
No caso da sinapse qu´ımica, o neurˆonio pr´e-sin´aptico passa por um potencial de a¸c˜ao,
ou spike, que nos terminais sin´apticos causa a entrada de c´alcio (Ca++
Figura 1.1: Esquema ilustrativo de sinapse excitat´oria ionotr´opica, isto ´e, que aumenta o potencial de membrana do neurˆonio p´os-sin´aptico, por meio dos neurotrans-missores que alteram a conforma¸c˜ao proteica de canal ligante-dependente, permitindo a
entrada de ´ıons positivos (Na+, neste caso). Fonte:
http://www.culturamix.com/cultura-/sinapse-artificial
Essas mudan¸cas na c´elula p´os-sin´aptica em rela¸c˜ao ao potencial de repouso, devido
`as sinapses, s˜ao chamadas de potenciais p´os-sin´apticos (PSPs, postsynaptic potentials),
podendo ser excitat´orios ou inibit´orios (despolariza¸c˜oes ou hiperpolariza¸c˜oes, respectiva-mente), dependendo do potencial de revers˜ao do receptor p´os-sin´aptico, o qual depende
do n´umero de canais perme´aveis aos ´ıons e da concentra¸c˜ao destes nos meios intra e
extracelulares. Em receptores ionotr´opicos, os PSPs excitat´orios podem ser gerados pela entrada dos ´ıons de s´odio (Na+
) e os PSPs inibit´orios podem ser gerados pela entrada de ´ıons de cloro (Cl−
) nas c´elulas, por exemplo. H´a contudo, o caso de c´elulas ciliares no ouvido interno, que transformam os sinais mecˆanicos do som em sinais el´etricos
neuro-nais, e para as quais o meio extracelular ´e mais abundante em pot´assio (K+) - sendo este
elemento o respons´avel pela despolariza¸c˜ao da membrana (diferentemente dos casos em que h´a influxo de Na+).
Caso ocorram PSPs excitat´orios, o potencial da membrana ficar´a temporariamente mais positivo em rela¸c˜ao ao que era. Se a despolariza¸c˜ao atingir um valor de voltagem chamado de limiar de disparo, o neurˆonio passa por uma fase de transmiss˜ao, ao longo da c´elula, de um sinal el´etrico, o j´a citado potencial de a¸c˜ao (spike). H´a, contudo, casos de neurˆonios que disparam potencial de a¸c˜ao ap´os uma hiperpolariza¸c˜ao, dependendo da dinˆamica de sua membrana [12].
´
E sabido que o n´umero de entradas sin´apticas recebidas por cada neurˆonio no sistema
sim-plesmente de PSP. Al´em disso, cov´em ressaltar que os registros utilizados neste trabalho tˆem dura¸c˜ao de 150ms, come¸cando 15ms antes da apresenta¸c˜ao do est´ımulo sonoro, e tˆem amplitude normalizada segundo estes primeiros 15ms (veja cap´ıtulo 2).
0 0.05 0.1 0.15
−2 0 2 4 6 8 10 12 14 16
Tempo (s)
Potencial na membrana (unidades normalizadas)
PSP
Figura 1.2: Exemplo de PSP na membrana de um neurˆonio do c´ortex auditivo de um
rato anestesiado sujeito a est´ımulo sonoro dos 15 aos 40ms (linha vermelha).
1.2
Potenciais de Campo Local
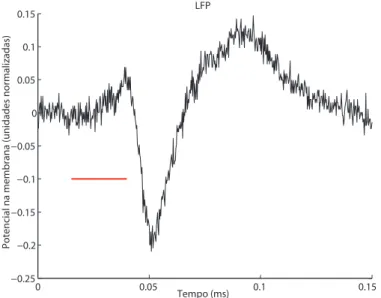
Potenciais de campo local (LFPs - Local Field Potentials) s˜ao registros eletrofisiol´ogicos obtidos invasivamente no c´erebro de animais sob experimento ou de pessoas sob
proce-dimentos cir´urgicos, por meio de eletrodos. Os LFPs s˜ao os sinais el´etricos registrados,
amplificados e filtrados a baixas frequˆencias (por exemplo,<500 Hz).
H´a o consenso de que os sinais de LFP apontem a atividade de uma popula¸c˜ao de neurˆonios, sendo gerados por oscila¸c˜oes lentas em suas membranas, as quais podem ser os potenciais p´os-sin´apticos ou altera¸c˜oes na membrana neuronal que ocorrem ap´os um
spike[14]. Devido `a contribui¸c˜ao da atividade s´ıncrona de v´arios neurˆonios sobre o sinal
de LFP, a pergunta de quantos neurˆonios est˜ao contribuindo n˜ao ´e de f´acil resposta e costuma-se referir ao espa¸co cortical que gerou o sinal. Essa regi˜ao varia em fun¸c˜ao da morfologia dos neurˆonios, de sua distribui¸c˜ao sin´aptica e da correla¸c˜ao da atividade sin´aptica entre eles, perpassando valores de poucas centenas de micrˆometros (µm) a v´arios mil´ımetros (mm) [15]. Outros fatores que influenciam o registro do LFP s˜ao o pr´oprio instrumento de medi¸c˜ao, o eletrodo, e suas caracter´ısticas, tais quais sua impedˆancia, o material de que ´e feito, sua geometria e superf´ıcie de contato.
de 150ms, come¸cando 15ms antes da apresenta¸c˜ao do est´ımulo sonoro, e tˆem amplitude normalizada segundo estes primeiros 15ms.
0 0.05 0.1 0.15
−0.25 −0.2 −0.15 −0.1 −0.05 0 0.05 0.1 0.15
Tempo (ms) LFP
Potencial na membrana (unidades normalizadas)
Figura 1.3: Exemplo de LFPobtido no c´ortex auditivo de um rato anestesiado sujeito
a est´ımulo sonoro dos 15 aos 40ms (linha vermelha).
1.3
C´
ortex Auditivo
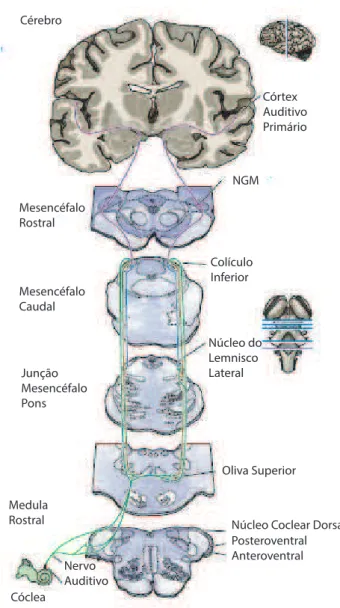
No sistema auditivo, a codifica¸c˜ao de est´ımulos sonoros ´e facilitada desde a c´oclea, o ´org˜ao sensitivo respons´avel pela audi¸c˜ao, por meio de c´elulas ciliadas. Estas c´elulas est˜ao organizadas tonotopicamente, i.e., estas c´elulas transdutoras est˜ao separadas segundo a frequˆencia sonora a que respondem mais intensamente, e essa divis˜ao ´e mantida at´e o c´ortex. O nervo auditivo estabelece a liga¸c˜ao entre as c´elulas ciliadas e o n´ucleo coclear,
o qual emite proje¸c˜oes para outros n´ucleos no tronco cerebral, a saber: complexo olivar
Cérebro
Córtex Auditivo Primário
NGM
Mesencéfalo Rostral
Mesencéfalo Caudal
Junção Mesencéfalo Pons
Núcleo Coclear Dorsal Posteroventral Anteroventral Oliva Superior Núcleo do Lemnisco Lateral
Colículo Inferior
Medula Rostral
Nervo Auditivo Cóclea
Figura 1.4: N´ucleos de retransmiss˜ao do sistema auditivo. Esquema simplificado,
da c´oclea ao c´ortex auditivo prim´ario (retirado da referˆencia [13]).
Estudos recentes investigaram medidas de dependˆencia entre est´ımulos e spikes no
c´ortex auditivo. Foi mostrado que a latˆencia das respostas despikes e a contagem de
spi-kes codificam est´ımulos sonoros [16]. Contudo, ainda n˜ao foi feita a an´alise da codifica¸c˜ao de est´ımulos auditivos por meio de PSPs e de como esta se compara com a codifica¸c˜ao realizada pelos LFPs. Como se acredita que o LFP seja resultado da a¸c˜ao coletiva de v´arios neurˆonios em uma vizinhan¸ca, ´e razo´avel ponderar se a dependˆencia de LFPs a est´ımulos sonoros ´e maior que a dos sinais individuais de PSPs. A an´alise de LFPs e
PSPs atrav´es da informa¸c˜ao m´utua, uma medida de dependˆencia, aplicada a diferentes
1.4
Objetivos
A presente disserta¸c˜ao prop˜oe-se a estudar registros intra (PSP) e extra (LFP) celulares do c´ortex auditivo de ratos anestesiados enquanto sujeitos a est´ımulos sonoros, a fim de quantificar a informa¸c˜ao m´utua entre estes est´ımulos e as respostas destes dois tipos de sinais. Al´em disso, verificar-se-´a se os sinais individuais de neurˆonios (PSPs) e os de conjunto de neurˆonios (LFPs) possuem rela¸c˜oes de sinergia ou redundˆancia. Para ambos os sinais, ser˜ao utilizados aspectos das respostas como amplitude e latˆencia.
1.5
Estrutura do Trabalho
O cap´ıtulo 2 descreve a obten¸c˜ao dos registros utilizados, enquanto o cap´ıtulo 3 apresenta uma abordagem resumida dos conceitos de Teoria da Informa¸c˜ao que ser˜ao relevantes na an´alise dos registros. Por sua vez, o cap´ıtulo 4 explora algumas t´ecnicas para retirada de vi´es nas estimativas feitas com os conceitos do cap´ıtulo 3. O cap´ıtulo 5 apresenta o tratamento feito com os registros para realizar estimativas. Finalmente, os resultados
obtidos nas estima¸c˜oes de informa¸c˜ao m´utua entre est´ımulos sonoros e PSPs/LFPs, al´em
Cap´ıtulo 2
Coleta dos dados
O banco de dados deste projeto foi obtido atrav´es da estimula¸c˜ao do c´ortex auditivo de ratos anestesiados por meio da apresenta¸c˜ao de 32 tons breves (25ms) de mesma amplitude (65dB) e frequˆencias distintas separadas logaritmicamente, apresentados em ordem fixa, pseudoaleat´oria, a uma taxa de dois por segundo e provenientes de mesma localiza¸c˜ao espacial (6cm `a direita do animal). As frequˆencias dos tons eram puras, variando de 2kHz a 46731Hz, sabe-se que neste intervalo as frequˆencias s˜ao aud´ıveis para ratos (a referˆencia [21] traz o intervalo aud´ıvel de 500Hz a 64kHz a 60dB, para ratos). Os registros foram
feitos emCold Spring Harbor Laboratory, em Nova York, E.U.A., de acordo com National
Institutes of Health guidelines e aprovadas pelo Cold Spring Harbor Laboratory Animal
Care and Use Committee. Esses dados est˜ao `a disposi¸c˜ao no site http://crcns.org/ [22],
o qual foi elaborado para compartilhar dados de alta qualidade e valor cient´ıfico em Neurociˆencias, a fim de facilitar pesquisas na neurociˆencia computacional. Os dados s˜ao uma contribui¸c˜ao de Michael DeWeese, da Universidade de Calif´ornia, Berkeley, atuando no laborat´orio de Anthony Zador.
A descri¸c˜ao detalhada dos procedimentos adotados para aquisi¸c˜ao ´e dada na referˆencia
[17]. De maneira resumida, os registros foram obtidos porpatch clamp in vivode 33 c´elulas
do c´ortex auditivo prim´ario de ratos, em regi˜oes diferentes deste c´ortex, de maneira a in-cluir diferentes regi˜oes de seu mapa tonot´opico, e em cerca de 17 animais (DeWeese, comunica¸c˜ao pessoal). O bloqueador de canal de s´odio r´apido, QX-314, tamb´em capaz de bloquear outras condutˆancias evocadas, foi adicionado `a solu¸c˜ao intracelular para evi-tar potenciais de a¸c˜ao. Dos 33 registros, 29 apresentam tamb´em registros extracelulares de LFPs, obtidos por outro eletrodo separado a uma distˆancia aproximada de 0,5mm da c´elula clampeada (a Figura 2.1 mostra um exemplo de aparelho para registro simultˆaneo
de PSP porpatch clampe de LFP por eletrodo,in vivo). Os registros foram feitos
continu-amente, sendo depois separados em matrizes de dura¸c˜ao de 150ms, come¸cando 15ms antes da apresenta¸c˜ao do est´ımulo - estas matrizes foram os registros utilizados neste trabalho. A anestesia utilizada foi pentobarbital (65mg/kg) em 16 dos registros, sendo que em 3 destes tamb´em foi usado diazepam (5mg/kg). Nos demais 17 registros foi utilizado ure-tano (1,5g/kg), ap´os cirurgia feita com ketamina (60mg/kg) e medetomedina (0,5mg/kg). A lista de c´elulas numeradas e separadas de acordo com a droga utilizada para registro encontra-se na tabela A.1, anexo A. Ap´os se encontrarem profundamente anestesiados, os animais foram colocados em uma conten¸c˜ao naso-orbital que mantinha as orelhas livres. Anest´esico local foi aplicado ao escalpo e uma craniotomia e durotomia 1x2mm foi feita
sobre o c´ortex auditivo esquerdo. A temperatura dos animais foi mantida a 37◦
C atrav´es
Figura 2.1: Exemplo de aparelho de registro simultˆaneo de PSP por patch clamp e
de LFP por eletrodo, in vivo (imagem retirada da referˆencia [23]).
Cap´ıtulo 3
Conceitos de Teoria da Informa¸c˜
ao
A Teoria da Informa¸c˜ao ´e uma ciˆencia relativamente recente se comparada com as demais
ciˆencias, foi estabelecida com a publica¸c˜ao do artigoA Mathematical Theory of
Commu-nication(“Uma Teoria Matem´atica da Comunica¸c˜ao”), de Claude Shannon, em 1948 [18].
O artigo procurou fixar medidas de comunica¸c˜ao que seriam - e s˜ao - importantes para resolver problemas de engenharia. Aspectos semˆanticos das mensagens foram ignorados em favor de outros aspectos, como por exemplo, de que um s´ımbolo enviado pelo emissor na mensagem seja o mesmo captado pelo receptor.
Uma das quantidades definidas na Teoria da Informa¸c˜ao ´e a entropia de um sinal
X, que pode ser denotada por H(X) (equa¸c˜ao 3.1). A entropia referida ao longo desta
disserta¸c˜ao ´e chamada de entropia de Shannon. O sinalX pode ser uma vari´avel aleat´oria
(v.a.) discreta de distribui¸c˜ao de probabilidade p(X). A entropia ´e uma medida da
incerteza de X, ou de qu˜ao dif´ıcil ´e prever seu resultado. Para v.a.s discretas, a entropia
´e m´axima quando sua distribui¸c˜ao ´e uniforme nosN valores queX pode assumir e dada
pela equa¸c˜ao 3.2. Equivalentemente, se X tiver maior chance de resultar em um valor
espec´ıfico em rela¸c˜ao aos demais, sua incerteza ser´a menor. F´ormula da entropia para vari´aveis discretas:
H(X) =−
N X
i=1
p(xi) log2p(xi). (3.1)
Entropia para vari´aveis discretas uniformes (m´axima entropia):
H(X) = −
N X
i=1
p(xi) log2p(xi)
= −
N X
i=1
1
N log2
1 N
= −log2
1 N
= log2N. (3.2)
Quando expressa pelo logaritmo na base 2, como na equa¸c˜ao 3.1, a medida de entropia
´e dada embits e no intervalo{H(X), H(X) + 1}se encontra o m´ınimo n´umero m´edio de
perguntas bin´arias necess´arias para se saber o valor de X.
Quando a v.a. X n˜ao pode assumir um determinado valor nunca, ou seja, p(X =
xk) = 0, h´a a conven¸c˜ao adotada de que 0 log20 = 0 no somat´orio da equa¸c˜ao 3.1, que ´e
O conceito de entropia pode ser extendido para pares de v.a.s, denominando-se entro-pia conjunta (equa¸c˜ao 3.3), e tamb´em para entroentro-pias condicionadas ao conhecimento de outra(s) v.a.(s) (equa¸c˜ao 3.4). A entropia conjunta mede a incerteza de v.a.s quando ob-servadas conjuntamente, j´a a entropia condicionada mede como o conhecimento de uma v.a. influencia a incerteza de outra.
H(X, Y) = −
N X
i=1
N X
j=1
p(xi, yj) log2p(xi, yj) (3.3)
H(X|Y) = −
N X
i=1
N X
j=1
p(xi, yj) log2p(xi|yj) (3.4)
A entropia conjunta pode ser escrita como:
H(X, Y) = −
N X
i=1
N X
j=1
p(xi, yj) log2p(xi, yj)
= −
N X
i=1
N X
j=1
p(xi, yj) log2p(yj|xi)p(xi)
= −
N X
i=1
N X
j=1
p(xi, yj) log2p(xi)− N X
i=1
N X
j=1
p(xi, yj) log2p(yj|xi)
= −
N X
i=1
p(xi) log2p(xi)− N X
i=1
N X
j=1
p(xi, yj) log2p(yj|xi)
= H(X) +H(Y|X) (3.5)
Assim, fica demonstrado que a entropia conjunta pode ser escrita por H(X, Y) =
H(X) +H(Y|X), a qual significa que a entropia conjunta de duas v.a.s ´e a soma da
entropia de uma delas e da entropia da outra condicionada `a primeira. Observe que, caso
o conhecimento da v.a. X nada acrescente para a determina¸c˜ao de Y, ou seja, caso as
vari´aveis X e Y sejam independentes, H(Y|X) = H(Y) e a incerteza conjunta ser´a a
soma das incertezas individuais. Contudo, em geral o condicionamento reduz a entropia (e a entropia conjunta ´e menor ou igual `a soma das entropias individuais).
A medida que utiliza conceitos mostrados at´e aqui e que ´e a mais utilizada na aplica¸c˜ao de Teoria da Informa¸c˜ao `as Neurociˆencias ´e a informa¸c˜ao m´utua. De maneira mais geral,
a informa¸c˜ao m´utua ´e uma medida semelhante a uma m´etrica, que indica o quanto duas
ou mais v.a.s s˜ao dependentes ou n˜ao (equa¸c˜ao 3.6) e sabe-se que ´e uma medida de dependˆencia mais geral que a correla¸c˜ao de Pearson, que indica apenas dependˆencias
lineares [19]. Mais precisamente, a informa¸c˜ao m´utua mede o quanto conhecer uma v.a.
I(X;Y) = N X
i=1
N X
j=1
p(xi, yj) log2
p(xi, yj) p(xi)p(yj)
(3.6)
= −
N X
i=1
N X
j=1
p(xi, yj) log2p(xi) + N X
i=1
N X
j=1
p(xi, yj) log2p(xi|yj)
I(X;Y) = H(X)−H(X|Y) (3.7)
A Figura 3.1 ilustra as rela¸c˜oes existentes entre entropia, entropia conjunta, entropia
condicionada e informa¸c˜ao m´utua para duas vari´aveis (X e Y).
Figura 3.1: Diagrama para entropia. Esquema para os conceitos de entropia, entropia
conjunta, entropia condicionada e informa¸c˜ao m´utua.
Uma das propriedades da entropia ´e nunca ser negativa, o que vale tamb´em para entropia condicionada [20]. Desse modo ´e poss´ıvel escrever:
I(X;Y) = H(X)−H(X|Y)
I(X;Y) ≤ H(X) (3.8)
I(X;Y) = H(Y)−H(Y|X)
I(X;Y) ≤ H(Y) (3.9)
Das inequa¸c˜oes 3.8 e 3.9, pode-se escrever:
I(X;Y)≤min(H(X), H(Y)), (3.10)
A qual significa que a informa¸c˜ao m´utua ´e limitada superiormente pelas entropias das
A informa¸c˜ao m´utua, assim como a entropia, tamb´em pode ser condicionada, conforme definida na equa¸c˜ao 3.11.
I(X;Y|Z) = H(X|Z)−H(X|Y, Z)
= X
x∈χ X
y∈Υ
X
z∈ζ
p(x, y, z) log2
p(x, y|z)
p(x|z)p(y|z) (3.11)
A entropia conjunta e a informa¸c˜ao m´utua podem ser extendidas para v´arias vari´aveis, segundo as regras da cadeia para entropia e informa¸c˜ao, equa¸c˜oes 3.12 e 3.13, respectiva-mente.
Regra da cadeia para entropia:
H(X1, ..., Xn) =
n X
i=1
H(Xi|Xi−1, ..., X1). (3.12)
Regra da cadeia para informa¸c˜ao:
I(X1, ..., Xn;Y) =
n X
i=1
I(Xi;Y|Xi−1, ..., X1). (3.13)
Al´em dos conceitos de entropia e informa¸c˜ao m´utua, outra defini¸c˜ao muito utilizada
em Neurociˆencias e que utiliza os conceitos j´a abordados ´e a de sinergia (equa¸c˜ao 3.14).
Syn(X;Y1, Y2) =I(X;Y1, Y2)−(I(X;Y1) +I(X;Y2)), (3.14)
Quando resulta em valor positivo, diz-se que h´a sinergia entre as v.a.s envolvidas, ou
que as v.a.sY1 eY2 agem sinergicamente fornecendo informa¸c˜ao sobre a v.a. X, de modo
que conjuntamente s˜ao mais informativas que quando analisadas individualmente. Por
outro lado, se o valor de Syn for negativo, ´e poss´ıvel afirmar que ocorre a situa¸c˜ao de
redundˆancia, e as v.a.s Y1 e Y2 trazem informa¸c˜oes similares sobre X.
Finalmente, outro conceito esclarecedor que a Teoria da Informa¸c˜ao traz ´e a
desigual-dade de processamento de dados. Para entendˆe-lo, definir-se-´a um processo de Markov,
denotado por X →Y →Z, para o qual pode-se escrever:
p(x, y, z) = p(x)p(y|x)p(z|y). (3.15)
A equa¸c˜ao 3.15 indica que em uma cadeia de Markov, a distribui¸c˜ao de probabilidade
da v.a. Z depende apenas deY, assim como a distribui¸c˜ao de probabilidade deY depende
apenas deX. Al´em disso, ´e poss´ıvel mostrar que as vari´aveisX eZ s˜ao condicionalmente
independentes em Y (p(x, z|y) =p(x|y)p(z|y)) [20].
A desigualdade de processamento de dados ´e o teorema que afirma que, se X →
Y → Z, ent˜ao I(X;Y) ≥ I(X;Z). ´E poss´ıvel intuir que o processamento de registros
eletrofisiol´ogicos dependa apenas da distribui¸c˜ao de probabilidade dos pr´oprios registros e que no c´ortex auditivo tais registros estejam condicionados aos est´ımulos sonoros. A desigualdade de processamento de dados ´e importante em Neurociˆencias porque mostra
que quaisquer processamentos ou estat´ısticas obtidas de registros associados `a v.a. X
poder˜ao informar no m´aximo tanto quanto os pr´oprios registros informariam. Em outras palavras, todas as manipula¸c˜oes que podem ser feitas com os registros podem apenas
Dessa maneira, as ferramentas da Teoria da Informa¸c˜ao abrem possibilidades de pes-quisa na busca por codifica¸c˜oes cerebrais que traduzam o ambiente externo, em especial
as que produzam os maiores valores de informa¸c˜ao m´utua entre est´ımulos sens´orios e
Cap´ıtulo 4
M´
etodos de corre¸c˜
ao de vi´
es
Conforme visto no cap´ıtulo 3, equa¸c˜oes 3.1 e 3.6, para calcular a entropia ou informa¸c˜ao m´utua entre vari´aveis aleat´orias, ´e necess´ario conhecer primeiro sua distribui¸c˜ao de pro-babilidade. Contudo, geralmente esta distribui¸c˜ao e/ou seus parˆametros n˜ao s˜ao conhe-cidos, da´ı a necessidade de amostras. Se X1, X2, . . . XN ´e uma amostra na vari´avel X
e θ ´e um parˆametro desconhecido da sua distribui¸c˜ao de probabilidade, por exemplo a
m´edia, uma fun¸c˜ao g(X1, X2, . . . , XN) para estimar θ ´e chamada de estimador; o valor ˆ
θ =g(x1, x2, . . . , xN) obtido para uma amostra poss´ıvel ´e chamado de estimativa.
Quando a m´edia que o estimador assume ´e diferente do verdadeiro valor do parˆametro, isto ´e, Eg(X1, X2, . . . , XN) 6= θ, diz-se que o estimador ´e enviesado, tendencioso ou viciado [24]. O desenvolvimento da equa¸c˜ao 4.1 traz o vi´es pela componente |θ−E(ˆθ)|, na desigualdade 4.3:
erro(θ) = |θ−θˆ| (4.1)
= |θ−E(ˆθ) +E(ˆθ)−θˆ| (4.2)
≤ |θ−E(ˆθ)|+|E(ˆθ)−θˆ| (4.3)
O estimador de probabilidade utilizado neste trabalho computa as frequˆencias rela-tivas dos valores que a vari´avel assume. No caso da distribui¸c˜ao conjunta das vari´aveis
X e Y, conta-se o n´umero de vezes que o par (X = x, Y = y) ocorreu e divide-se pelo
tamanho da amostra (ˆp(x, y) = n(x, y)/N). Este estimador ´e o de m´axima
verossimi-lhan¸ca para as probabilidades, o que significa que ele traz estimativas de probabilidade que maximizam a fun¸c˜ao de verossimilhan¸ca [25]. Quando a estima¸c˜ao de informa¸c˜ao ´e feita usando probabilidades obtidas segundo o estimador de m´axima verossimilhan¸ca,
chama-se o estimador de informa¸c˜ao deplug-in ounaive.
A Figura 4.1, adaptada da referˆencia [29], ilustra o caso em que dois neurˆonios hi-pot´eticos apresentam probabilidades de respostas (de um a dez) quando s˜ao apresentados a dois est´ımulos diferentes. O primeiro neurˆonio n˜ao apresenta nenhuma informa¸c˜ao so-bre qual est´ımulo foi apresentado, possuindo a mesma probabilidade de emitir qualquer uma das respostas, seja qual for o est´ımulo. O segundo neurˆonio apresenta probabilidades
diferentes para as respostas, condicionadas ao est´ımulo. As estimativas naive de
proba-bilidade s˜ao mostradas pelas linhas cinzas. Os histogramas mais `a direita mostram 5000
estimativas para informa¸c˜ao m´utua entre est´ımulos e respostas para os dois neurˆonios,
de informa¸c˜ao m´utua. Para ambos neurˆonios, o vi´es ´e reduzido quando a amostra ´e de maior tamanho (isto ´e, no caso do tamanho amostral 100).
Neurônio não informativo
Estímulo 1 Estímulo 2
Neurônio informativo
Estímulo 1 Estímulo 2
P
robabilidade
20 amostr
as
100 a
most
ras
0.1
0.1 0.3
0.3
Respostas Respostas 1 5 10 1 5 10
1 5 10
1 5 10 0 0.2 0.5
0 0.2 0.5 0.1
0.5 0.1
Informação Respostas Respostas
P
robabilidade
20 amostr
as
100 a
most
ras
0.1 0.1 0.3 0.3
1 5 10 1 5 10
1 5 10 1 5 10
Informação 0.1
0.2 0.1
0.5 0.66 1 0.5 0.66 1
Figura 4.1: Exemplo de emprego do estimador naive. Nos gr´aficos do lado
es-querdo, as probabilidades verdadeiras para cada uma das dez respostas `a apresenta¸c˜ao do est´ımulo 1 ou 2 s˜ao indicadas pela curva preta. Exemplos obtidos para as estima-tivas de probabilidades s˜ao mostrados nas curvas cinzas. No lado direito, frequˆencias de estimativas de informa¸c˜ao m´utua utilizando o estimador naive, para 5000 simula¸c˜oes.
O valor verdadeiro de informa¸c˜ao m´utua para os dois neurˆonios ´e indicado pela linha
tracejada. Para os dois neurˆonios, no painel superior as amostras possuem tamanho 20; no painel inferior, tamanho 100. Figura adaptada da referˆencia [29].
A estima¸c˜ao de entropia e de informa¸c˜ao m´utua usando o estimadornaive´e enviesada
e algumas aproxima¸c˜oes de vi´es foram desenvolvidas quando se est´a em regime assint´otico de amostragem [26]. O regime assint´otico pode ser descrito, de maneira informal, como um regime em que h´a muitas amostras em rela¸c˜ao `as respostas poss´ıveis que as amostras
podem resultar. De maneira mais formal, sejam r a vari´avel aleat´oria que associa um
valor `a resposta neurofisiol´ogica e s a que associa um valor ao est´ımulo apresentado,
temos que NsP(r|s)≫1. P(r|s) representa a probabilidade condicionada de registrar-se
a resposta r dado ter sido s o est´ımulo apresentado, Ns ´e o n´umero de amostras em
resposta `a apresenta¸c˜ao do est´ımulo s. As express˜oes aproximadas de vi´es para este caso s˜ao:
vies[H(R)] = −1
2Nln 2( ¯R−1)
vies[H(R|S)] = −1
2Nln 2
X
s
( ¯Rs−1)
vies[I(R;S)] = 1
2Nln 2
"
X
s
( ¯Rs−1)−( ¯R−1) #
(4.4)
Nessas express˜oes, N ´e o n´umero total de amostras, ¯R ´e o n´umero de respostas
que apresentam probabilidade n˜ao nula de ocorrˆencia e ¯Rs ´e o n´umero de respostas
condicionadas `a apresenta¸c˜ao de uma entrada s que apresentam probabilidade n˜ao nula
Apesar de n˜ao apresentarem valores exatos para o vi´es e serem derivadas para situa¸c˜oes quase ideais de regime assint´otico, as express˜oes em 4.4 permitem algum esclarecimento sobre o comportamento do vi´es conforme as amostras aumentam. Percebe-se, por
exem-plo, que quando o n´umero de amostras ´e constante, o vi´es da informa¸c˜ao m´utua aumenta
quando h´a mais valores poss´ıveis para as respostas neurofisiol´ogicas. Esta ´e uma situa¸c˜ao muito frequente em neurociˆencias, pois os dados neurofisiol´ogicos costumam formar um amplo espa¸co de respostas, em particular os referentes a c´odigos temporais. Para estes, o
universo de respostas poss´ıveis cresce exponencialmente com o n´umero debins, ou janelas
de tempo, que s˜ao registradas - se cada bin registrar 0 ou 1 spike, o n´umero de respostas poss´ıveis ser´a 2L (L o n´umero de bins).
Felizmente, diante do exposto, h´a m´etodos para a corre¸c˜ao do vi´es na estima¸c˜ao
de entropia e informa¸c˜ao m´utua. Alguns dos mais utilizados e retratados a seguir s˜ao:
Shuffling, m´etodo de Panzeri-Treves e extrapola¸c˜ao quadr´atica (QE - Quadratic
Extra-polation).
4.1
M´
etodo
Shuffling
O m´etodoshuffling´e utilizado apenas para estima¸c˜ao de informa¸c˜ao m´utua e consiste em
parear as vari´aveis aleat´orias s de est´ımulo e r de resposta aleatoriamente, encontrando
um valor de informa¸c˜ao m´utua que seria supostamente nulo, uma vez que as poss´ıveis
correla¸c˜oes entre as duas vari´aveis estariam sendo destru´ıdas. Esse procedimento ´e repe-tido v´arias vezes e a m´edia dos valores obrepe-tidos,Ishuf f led(S;R), ´e subtra´ıda da estimativa
naive [27].
Itrue(S;R) =Inaive(S;R)−Ishuf f led(S;R) (4.5)
O m´etodo apresenta, no entanto, algumas restri¸c˜oes quanto a sua utiliza¸c˜ao em di-ferentes tipos de discretiza¸c˜ao e s´o pode ser usado quando as seguintes condi¸c˜oes s˜ao atendidas:
1. Distribui¸c˜ao de probabilidade estimada para vari´aveis aleat´orias discretas ou
sim-plesmente discretizadas (m´etodos que utilizem convolu¸c˜ao com fun¸c˜oes kernel n˜ao
devem usar este m´etodo para retirada de vi´es);
2. Nenhum par (s, r) pode apresentar probabilidade nula de ocorrˆencia (caso contr´ario haver´a superestima¸c˜ao de vi´es).
4.2
M´
etodo
Panzeri-Treves
(PT)
Panzeri e Treves encontraram que a m´edia da diferen¸ca entre a informa¸c˜ao m´utua
ver-dadeira e a estimativa naive, para todas as combina¸c˜oes de amostras de tamanho N, ´e
dada por uma s´erie infinita. Al´em disso, verificaram que se o primeiro termo dessa s´erie, C1, for suficientemente menor que um, ele ´e uma estimativa de vi´es poss´ıvel [27].
C1 =
1
2Nln 2(S−1)( ¯R−1) (4.6)
a frequˆencia relativa de uma resposta depende apenas da probabilidade subjacente de ocorrˆencia desta, independentemente da ocorrˆencia das demais.
A equa¸c˜ao 4.6 ´e a mesma em 4.4, s´o que considera que todas as respostasr poss´ıveis
ocorrem para cada um dos est´ımulos s ( ¯Rs = ¯R), como ´e desenvolvido abaixo.
1
2Nln 2
"
X
s
( ¯Rs−1)−( ¯R−1) #
= 1
2Nln 2
"
X
s
( ¯R−1)−( ¯R−1) #
= 1
2Nln 2[S( ¯R−1)−( ¯R−1)]
= 1
2Nln 2[(S−1)( ¯R−1)]
= C1
Essa suposi¸c˜ao, contudo, ´e uma superestima¸c˜ao do vi´es quando h´a de fato respostas que n˜ao ocorrem para um determinado est´ımulo. Por outro lado, a contagem simples de ocorrˆencia de respostas aos est´ımulos pode subestimar o verdadeiro valor do vi´es, haja visto que as amostras podem ter sido insuficientes para refletir toda a sua variabilidade. Dessa forma, o m´etodo PT consiste em empregar a teoria de Bayes, que utiliza pro-babilidades condicionadas, para estimar os valores de ¯R e ¯Rs, aplicando-os na equa¸c˜ao
4.4 para estimar o vi´es. Uma vez estimado, o vi´es ´e subtra´ıdo da estimativa naive de
informa¸c˜ao m´utua [28].
4.3
M´
etodo da extrapola¸c˜
ao quadr´
atica (QE)
O m´etodo da extrapola¸c˜ao quadr´atica, utilizado para estima¸c˜ao de entropia e informa¸c˜ao
m´utua, consiste em dividir o conjunto de amostras em parti¸c˜oes aleat´orias com 1/2 e
1/4 do tamanho populacional original. Para estes subgrupos, s˜ao estimadas a
entro-pia ou informa¸c˜ao m´utua, segundo o m´etodo naive, sendo feita em seguida a m´edia
dos dois e quatro valores estimados. Os pares (N, Inaive1(S;R)), (N/2, Inaive2(S;R)) e
(N/4, Inaive4(S;R)) s˜ao usados de forma a ajustar os parˆametros Itrue, a e b da curva descrita nas equa¸c˜oes 4.7, 4.8 e 4.9 abaixo:
Inaive1(S;R) = Itrue(S;R) + a
N +
b
N2 (4.7)
Inaive2(S;R) = Itrue(S;R) + a
(N/2) +
b
(N/2)2 (4.8)
Inaive4(S;R) = Itrue(S;R) + a
(N/4) +
b
(N/4)2 (4.9)
O valor encontrado para o parˆametro Itrue(S;R) ´e a aproxima¸c˜ao da informa¸c˜ao
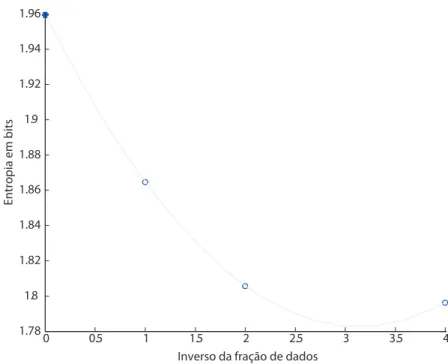
m´utua, corrigida. A Figura 4.2 ilustra um ajuste feito de estima¸c˜ao QE para entropia de
0 0.5 1 1.5 2 2.5 3 3.5 4 1.78
1.8 1.82 1.84 1.86 1.88 1.9 1.92 1.94 1.96
Inverso da fração de dados
Entr
opia em bits
Figura 4.2: Exemplo do m´etodo QE para estima¸c˜ao de entropia, para distribui¸c˜ao
homogˆenea e R = 4. As circunferˆencias azuis indicam estimativas de entropia
conside-rando a amostra inteiramente (abscissa 1) e consideconside-rando as m´edias de estimativas usando parti¸c˜oes da amostra (abscissas 2 e 4). O asterisco azul indica a estimativa QE. O valor te´orico de entropia ´e 2 bits.
4.4
Simula¸c˜
oes comparativas entre os m´
etodos
Diante dos diversos m´etodos explicados, surge a d´uvida sobre qual m´etodo de corre¸c˜ao
de vi´es ´e mais eficiente para os dados deste trabalho, no sentido de apresentar
estimati-vas mais confi´aveis e pr´oximas dos valores verdadeiros de entropia e informa¸c˜ao m´utua.
Para encontrar o m´etodo a ser utilizado com os dados experimentais, foram realizadas simula¸c˜oes e estimativas com pares de vari´aveis aleat´orias cujas verdadeiras informa¸c˜oes
m´utuas eram conhecidas. Foi utilizado o pacote para o software Matlab apresentado em
[30].
Para gerar conjuntos de dados, considerou-se o resultado obtido na referˆencia [31], segundo a qual a libera¸c˜ao de ves´ıculas de neurotransmissores em sinapses, quando os
PSPs s˜ao relativamente pequenos, seguem uma distribui¸c˜ao dePoisson. Esta distribui¸c˜ao
pode ser obtida por uma distribui¸c˜ao binomial, mantendo o produto de seus parˆametros
N p equivalente `a taxa λ da Poisson (al´em disso, N → ∞ e p → 0). Desta forma, em
nosso modelo hipot´etico geramos uma vari´avel aleat´oriaS, uniforme no conjunto discreto
{1,2, . . . ,32}, assim como ´e uniforme a distribui¸c˜ao dos est´ımulos sonoros apresentados
aos ratos cujos registros foram utilizados neste trabalho. A vari´avel S determinou o
parˆametro p de uma distribui¸c˜ao binomial da vari´avel R de parˆametros N = 3 e p(s)
(p(s)∈ {1/33,2/32, . . . ,32/33}). As estimativas obtidas de informa¸c˜ao m´utua entreS e
R pelos diferentes m´etodos apresentados nas se¸c˜oes acima, bem como estimativas naive,
0.4 0.5 0.6 0.7 0.8 0.9 0
5 10 15 20
Naive
0.4 0.5 0.6 0.7 0.8 0.9 0
5 10 15
20 QE
0.4 0.5 0.6 0.7 0.8 0.9 0
5 10 15
20 PT
0.4 0.5 0.6 0.7 0.8 0.9 0
5 10 15 20
Shuf
Bits Bits
Naive QE PT Shuf
0 0.05
0.1 0.15 0.2 0.25
RMSE
(A) (B)
(C) (D)
0 0.02 0.04 0.06 0.08 0.1
RMSE
Naive QE PT Shuf
0.55 0.6 0.65 0.7 0
5 10 15
Naive
0.55 0.6 0.65 0.7 0
5 10 15 20
QE
0.55 0.6 0.65 0.7 0
5 10 15
PT
0.55 0.6 0.65 0.7 0
5 10 15
Shuf
Bits Bits
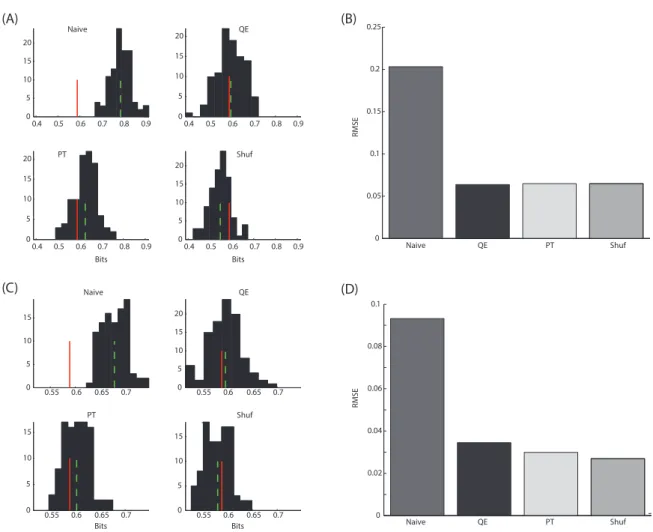
Figura 4.3: Compara¸c˜ao entre os m´etodos de corre¸c˜ao de vi´es.(A, C):
Histo-gramas de estimativas de informa¸c˜ao m´utua entre a vari´avel R, de distribui¸c˜ao
bino-mial, quando condicionada a uma realiza¸c˜ao da vari´avel S, de distribui¸c˜ao uniforme.
Parˆametros da binomial: N = 3 e p ∈ {1/33, 2/33, . . . ,32/33}. Valor verdadeiro
in-dicado pela linha vermelha. M´edia das estimativas na linha tracejada verde. (B, D):
Raiz do erro m´edio quadr´atico (RMSE) para os 4 m´etodos testados: naive; QE -
extra-pola¸c˜ao quadr´atica; PT - Panzeri Treves; Shuf - Shuffling, para as estimativas em (A,
C), respectivamente. Tamanhos amostrais em (A, B): 320; em (C, D): 720.
Observando os gr´aficos da Figuras 4.3, percebe-se que os desempenhos dos diferentes m´etodos nas condi¸c˜oes referidas foram semelhantes. A raiz do erro m´edio quadr´atico
(RMSE -root mean squared error) foi a medida usada para avalia¸c˜ao de qu˜ao distantes do
valor verdadeiro estavam as estimativas feitas. Observando mais atentamente os gr´aficos
das sub figuras (B, D), ´e poss´ıvel notar que nos casos de tamanho amostral N = 720 os
valores de RMSE s˜ao menores. Essa conclus˜ao j´a era esperada, uma vez que quando a amostra ´e maior, os valores estimados de probabilidades s˜ao mais pr´oximos dos reais e o
vi´es ´e diminu´ıdo (veja N no denominador das equa¸c˜oes 4.4). O m´etodo PT apresentou
Cap´ıtulo 5
An´
alise dos dados
Este cap´ıtulo apresenta a an´alise feita sobre os registros eletrofisiol´ogicos a fim de estimar a informa¸c˜ao m´utua entre eles e os est´ımulos sonoros. A an´alise seguiu os seguintes passos:
• Suaviza¸c˜ao das curvas;
• Retirada de amostras discrepantes;
• Verifica¸c˜ao de n´umero de amostras n˜ao discrepantes;
• Obten¸c˜ao de amplitudes e latˆencias.
5.1
Suaviza¸c˜
ao das curvas
−10 0 10 2000 −5 0 5 2214 −5 0 5 10 2451 0 5 10 2713 −10 −5 0 5 3003 −10 0 10 20 3325 −20 −10 0 10 3681 0 5 10 15 4074 −10 0 10 4510 0 5 10 4993 −5 0 5 10 15 5527 −5 0 5 10 6119 −5 0 5 10 15 6773 0 10 20 30 7498 0 10 20 8300 0 10 20 30 9188 −20 0 20 40 10172 −10 0 10 20 30 11260 −10 0 10 12465 −5 0 5 10 15 13799 −20 0 20 15275 −30 −20 −10 0 10 16909 −5 0 5 10 18719 0 10 20 20722 −5 0 5 10 15 22939 −5 0 5 10 25393 −10 0 10 20 30 28110 −10 0 10 20 31118 −5 0 5 10 15 34448 −20 −10 0 10 38134 −10 0 10 20 30 42214 −10 0 10 46731
0 150
tempo (ms) Hz P ot encial (a) PSPs −0.6 −0.4 −0.2 0 0.2 2000 −0.4 −0.2 0 0.2 2214 −0.4 −0.20 0.2 0.4 0.6 0.8 2451 −0.2 0 0.2 2713 −0.3 −0.2 −0.1 0 0.1 3003 −0.4 −0.2 0 0.2 3325 −0.6 −0.4 −0.2 0 0.2 3681 −0.2 0 0.2 4074 −0.2 0 0.2 0.4 4510 −0.1 0 0.1 0.2 4993 −0.6 −0.4 −0.2 0 0.2 5527 −0.3 −0.2 −0.1 0 0.1 6119 −0.2 0 0.2 0.4 0.6 6773 −1 −0.5 0 7498 −0.4 −0.2 0 0.2 0.4 8300 −0.6 −0.4 −0.2 0 0.2 9188 −1.5 −1 −0.5 0 0.5 10172 −1 −0.5 0 11260 −0.4 −0.2 0 0.2 12465 −1 −0.5 0 13799 −1 −0.5 0 0.5 15275 −0.6 −0.4 −0.2 0 0.2 16909 −0.1 0 0.1 18719 −0.4 −0.2 0 0.2 20722 −0.6 −0.4 −0.2 0 0.2 22939 −0.2 −0.1 0 0.1 25393 −0.8 −0.6 −0.4 −0.2 0 28110 −0.6 −0.4 −0.2 0 31118 −0.4 −0.2 0 34448 −0.5 0 0.5 38134 −0.6 −0.4 −0.2 0 0.2 42214 −1 −0.5 0 46731
0 150 tempo (ms) Hz P ot encial (b) LFPs
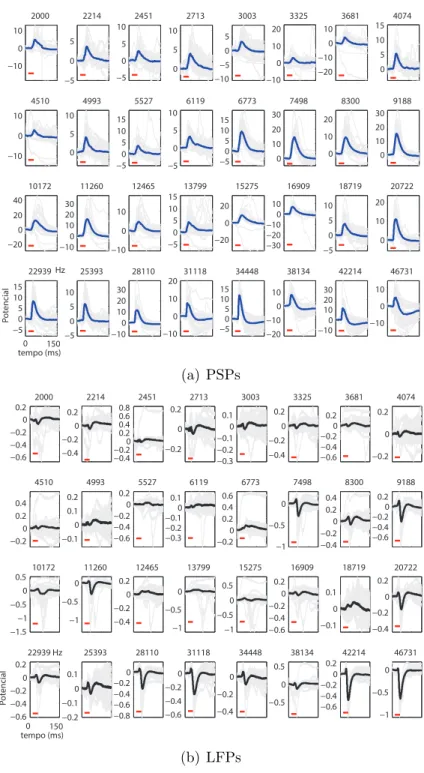
Figura 5.1: Exemplos de registros. Registros da c´elula 13 (030201md03b), separados
pela frequˆencia do est´ımulo sonoro. Tra¸cados m´edios de PSPs, nas curvas azuis, e de LFPs nas curvas pretas - feitos na vizinhan¸ca da c´elula (ver cap´ıtulo 2). As curvas em cinza mostram todos os registros para cada uma das 32 frequˆencias de est´ımulo (indicada no topo de cada minigr´afico). O potencial el´etrico ´e dado em rela¸c˜ao ao baseline (potencial m´edio nos 15 ms anteriores `a apresenta¸c˜ao do est´ımulo). O tra¸co vermelho indica o per´ıodo de apresenta¸c˜ao do est´ımulo.
Exemplos dos efeitos obtidos est˜ao ilustrados na Figura 5.2.
−0.5 −0.4 −0.3 −0.2 −0.1 0
−5 0 5 10
potencial (unidades normalizadas)
0.05 0.1 0.15
tempo (s)
0.05 0.1 0.15
tempo (s)
PSP LFP
Figura 5.2: Filtragem de registros. Sinais de PSP (`a esquerda) e LFP (`a direita), com
e sem a passagem por filtro passa-baixas a 300 Hz (curvas preta e azul, respectivamente). Exemplos da c´elula 13 (030201md03b) em resposta ao est´ımulo sonoro de 5527Hz. O potencial el´etrico, para cada sinal, ´e normalizado em rela¸c˜ao ao baseline (potencial m´edio nos primeiros 15 ms, anteriores `a apresenta¸c˜ao do est´ımulo, indicada pelo tra¸co vermelho).
5.2
Retirada de Amostras Discrepantes
O segundo passo na an´alise foi a exclus˜ao de registros eletrofisiol´ogicos discrepantes. O crit´erio utilizado foi o mesmo da referˆencia [17], observando-se o quanto o formato do tra¸cado se distancia do formato m´edio de tra¸cados em uma dada c´elula e para um mesmo est´ımulo, j´a filtrados. A an´alise ´e feita da seguinte forma: cada sinal ´e visto como um vetor e sua proje¸c˜ao ´e feita sobre o sinal m´edio:
proj~v~u=
~u.~v ~v.~v
~v. (5.1)
Na equa¸c˜ao 5.1, ~u ´e o vetor de uma resposta, e~v ´e o vetor da resposta m´edia. Os
vetores s˜ao considerados em apenas 100ms, come¸cando 15ms antes da apresenta¸c˜ao do
est´ımulo. Em seguida, a raiz do erro m´edio quadr´atico (rmse - root mean squared error)
entre a resposta m´edia e a proje¸c˜ao ´e calculada. Caso o escalar multiplicando o vetor~v
seja negativo ou o rmse calculado esteja acima de 0,35 vezes a distˆancia do m´ınimo ao pico da resposta m´edia para cada tom, o sinal ´e considerado discrepante e ´e exclu´ıdo das demais etapas de an´alise.
curva azul e exclu´ıdas, na curva vinho. Nota-se que os formatos das curvas azul e preta s˜ao similares, ao passo que a curva vinho mostra-se distante da m´edia.
0 0.02 0.04 0.06 0.08 0.1
−1 0 1 2 3 4 5
tempo (s)
potencial (unidades normalizadas)
PSPs
0 0.02 0.04 0.06 0.08 0.1
−0.12 −0.1 −0.08 −0.06 −0.04 −0.02 0 0.02 0.04 0.06
tempo (s) LFPs
Figura 5.3: Sinais discrepantes. M´edia dos sinais satisfat´orios, em azul, e discrepantes,
em vinho, segundo crit´erio sugerido na referˆencia [17]. Curvas em preto s˜ao a m´edia dos sinais de PSP (`a esquerda) e LFP (`a direita) j´a filtrados a 300 Hz. Exemplos da c´elula 13 (030201md03b) em resposta ao est´ımulo sonoro de 38134Hz. Apresenta¸c˜ao do est´ımulo indicada pelo tra¸co vermelho.
5.3
N´
umero de Amostras N˜
ao Discrepantes
O passo seguinte na an´alise foi a verifica¸c˜ao do n´umero de amostras n˜ao discrepantes, pois
um n´umero muito reduzido de amostras n˜ao permite estimativas coerentes de informa¸c˜ao
m´utua, mesmo aplicando uma corre¸c˜ao como PT, a escolhida no cap´ıtulo 4. As Figuras
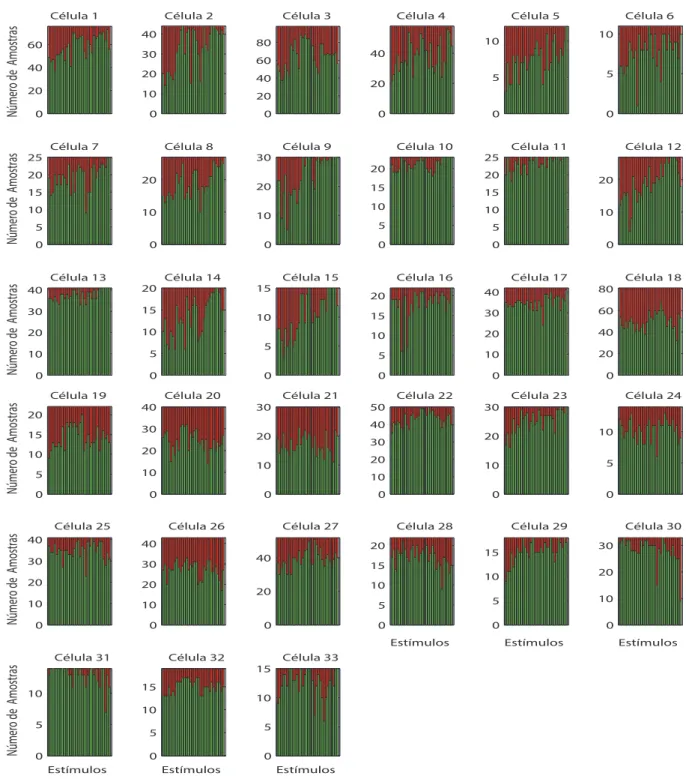
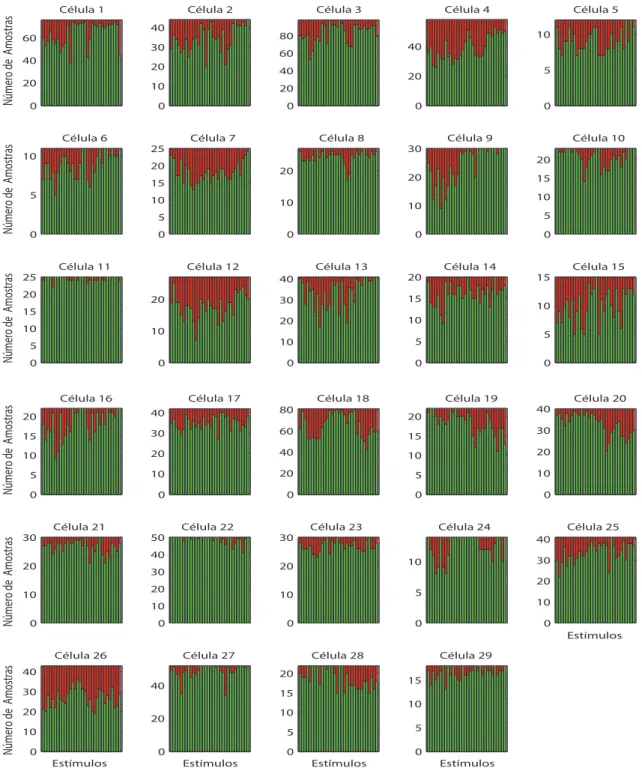
5.4 e 5.5 indicam para cada c´elula e est´ımulo o n´umero de amostras restantes. Na Figura
5.6 est˜ao apresentados o n´umero de registros simultˆaneos n˜ao discrepantes de PSPs e
LFPs. No anexo C, as tabelas C.1, C.2 e C.3 mostram detalhadamente, para cada
c´elula e est´ımulo, o n´umero de amostras restantes expressas nas Figuras 5.4, 5.5 e 5.6,
0 20 40 60
Célula 1
0 10 20 30 40
Célula 2
0 20 40 60 80
Célula 3
0 20 40
Célula 4
0 5 10
Célula 5
0 5 10
Célula 6
0 5 10 15 20 25
Célula 7
0 10 20
Célula 8
0 10 20 30
Célula 9
0 5 10 15 20
Célula 10
0 5 10 15 20 25
Célula 11
0 10 20
Célula 12
0 10 20 30 40
Número
d
e
Amostra
s Célula 13
0 5 10 15 20
Célula 14
0 5 10 15
Célula 15
0 5 10 15 20
Célula 16
0 10 20 30 40
Célula 17
0 20 40 60 80
Célula 18
0 5 10 15 20
0 10 20 30 40
0 10 20 30
0 10 20 30 40 50
0 10 20 30
0 5 10
0 10 20 30 40
0 10 20 30 40
0 20 40
0 5 10 15 20
0 5 10 15
0 10 20 30
0 5 10
Estímulos
0 5 10 15
0 5 10 15
Número
d
e
Amostra
s
Número
d
e
Amostra
s
Númer
o d
e
Amostra
s
Númer
o d
e
Amostra
s
Númer
o d
e
Amostra
s Célula 19 Célula 20
Célula 25 Célula 26 Célula 27 Célula 28 Célula 29 Célula 30 Célula 21 Célula 22 Célula 23 Célula 24
Célula 31 Célula 32 Célula 33
Estímulos Estímulos
Estímulos Estímulos Estímulos
Figura 5.4: N´umero de amostras de PSPs. Os gr´aficos mostram o n´umero de
0 20 40 60
Célula 1
0 10 20 30 40
Célula 2
0 20 40 60 80
Célula 3
0 20 40
Célula 4
0 5 10
Célula 5
0 5 10
Célula 6
0 5 10 15 20 25
Célula 7
0 10 20
Célula 8
0 10 20 30
Célula 9
0 5 10 15 20
Célula 10
0 5 10 15 20 25
Célula 11
0 10 20
Célula 12
0 10 20 30 40
Célula 13
0 5 10 15 20
Célula 14
0 5 10 15
Célula 15
0 5 10 15 20
Célula 16
0 10 20 30 40
Célula 17
0 20 40 60 80
Célula 18
0 5 10 15 20
Célula 19
0 10 20 30 40
Célula 20
0 10 20 30
Célula 21
0 10 20 30 40 50
Célula 22
0 10 20 30
Célula 23
0 5 10
Célula 24
0 10 20 30 40
Célula 25
0 10 20 30 40
Célula 26
0 20 40
Célula 27
0 5 10 15 20
Célula 28
0 5 10 15
Célula 29
Número
d
e
Amostras
Número
d
e
Amostras
Número
d
e
Amostras
Númer
o
d
e
Amostra
s
Númer
o
d
e
Amostra
s
Númer
o
d
e
Amostra
s
Estímulos
Estímulos
Estímulos Estímulos
Estímulos
Figura 5.5: N´umeros de amostras de LFPs. Os gr´aficos mostram o n´umero de
0 20 40
Célula 16
0 20 40
Célula 17
0 20 40
Célula 18
0 20 40
Célula 19
0 20 40
Célula 20
0 20 40
Célula 21
0 20 40
Célula 22
0 20 40
Célula 23
0 20 40
Célula 24
0 20 40
Célula 25
0 20 40
Nú
mero d
e Amostras
Célula 26
0 20 40
Célula 27
0 20 40
Célula 28
0 20 40
Célula 29
Nú
mero d
e Amostras
Nú
mero d
e Amostras
Estímulos Estímulos Estímulos Estímulos
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
0 20 40
Célula 11
0 20 40
Célula 12
0 20 40
Célula 13
0 20 40
Célula 14
0 20 40
Célula 15
Número de Amostras
Número de Amostras
Número de Amostras
Célula 1 Célula 2 Célula 3 Célula 4 Célula 5
Célula 6 Célula 7 Célula 8 Célula 9 Célula 10
Estímulos
Figura 5.6: N´umeros de amostras de PSPs e LFPs. O n´umero de amostras n˜ao
discrepantes, para cada um dos 32 est´ımulos (frequˆencias) apresentados, para as 29 c´elulas com registros simultˆaneos de PSPs e de LFPs.
A fim de investigar o n´umero de amostras necess´arias para estimativas pr´oximas
do valor verdadeiro de informa¸c˜ao, realizaram-se simula¸c˜oes as quais consideraram as distribui¸c˜oes emp´ıricas (naive) de probabilidade de amplitudes de PSPs da c´elula 13. As amplitudess foram obtidas ap´os a realiza¸c˜ao do processo da se¸c˜ao 5.2 e foram discretizadas
em classes igualmente populosas (R = 4 ou R = 16 classes). Pares de est´ımulos e
respostas foram gerados, em que os est´ımulos foram repetidos aproximadamente o mesmo
10 20 30 40 50 60 0
2 4 6 8 10 12 14 16 18 20 22
Ns
Erro percentual
(a) R= 4
10 20 30 40 50 60
0 2 4 6 8 10 12 14 16 18 20 22
Ns
Erro percentual
(b) R= 16
Figura 5.7: Erro percentual m´edio e n´umero de amostras. Erro percentual m´edio
obtido nas 100 estimativas feitas com simula¸c˜oes em fun¸c˜ao do n´umero de amostras por
est´ımulo (Ns), e c´alculo dos desvios padr˜oes dos erros, indicadas nas linhas vermelhas. Si-mula¸c˜oes feitas considerando como modelo a distribui¸c˜ao emp´ırica (naive) de amplitudes de PSPs da c´elula 13, condicionadas aos est´ımulos.
geradas segundo a distribui¸c˜ao naive de probabilidade condicionada das respostas aos
est´ımulos. Observando o n´umero Ns m´edio de amostras por est´ımulo de cada c´elula,
variou-se o tamanho amostral nas simula¸c˜oes, indo de Ns m´ınimo at´e Ns m´aximo entre
as c´elulas com registros de PSP. As simula¸c˜oes foram feitas 100 vezes para cada valor de Ns, computando-se estimativas de informa¸c˜ao m´utua, com a corre¸c˜ao PT, e calculando-se o erro percentual m´edio para as 100 estimativas em seguida. Os resultados obtidos est˜ao expressos na Figura 5.7.
Um fator interessante e observ´avel na Figura 5.7 foi a diminui¸c˜ao do erro percentual
m´edio e do desvio padr˜ao do erro conforme Ns cresce. Em particular, observa-se que
para o caso de poucas amostras, tanto o erro quanto seu desvio padr˜ao s˜ao menores
quando a resposta ´e discretizada em R = 4. Teoricamente, quando h´a muitas amostras,
a discretiza¸c˜ao em mais classes ´e mais interessante, retratando melhor a resposta e, por
conseguinte, o est´ımulo, pois o limite imposto pela entropia da vari´avel R ´e aumentado.
Na pr´atica, o observado nas simula¸c˜oes foi que quando h´a mais amostras, nem o erro percentual m´edio nem o desvio padr˜ao dos erros percentuais diminu´ıram muito.
Por-tanto, sabendo da necessidade de escolher um ´unico n´umero para representar as repostas
eletrofisiol´ogicas e comparar estimativas, optou-se por manterR = 4, haja visto que para
as c´elulas menos amostradas este n´umero causa menores erros e desvios e, para as mais
amostradas, apesar do n´umero ser conservador, o desempenho foi similar ao casoR = 16.
Al´em disso, para o caso de estimativas de informa¸c˜ao conjunta, a discretiza¸c˜ao de cada
resposta em 16 n´ıveis traria uma resposta conjunta que poderia atingir 16×16 = 256
valo-res, enquanto a informa¸c˜ao conjunta da resposta discretizada em 4 n´ıveis atinge o m´aximo de apenas 16 valores poss´ıveis, causando menor vi´es (veja o numerador na equa¸c˜ao de aproxima¸c˜ao de vi´es, equa¸c˜ao 4.4, cap´ıtulo 4).
Ainda observando-se a Figura 5.7, agora para o caso R= 4 em particular, percebe-se
que o erro percentual m´edio ´e menor que 10% para Ns≥8, que constitui um valor baixo
de erro. Dessa forma, optou-se por excluir da an´alise as c´elulas que apresentaram Ns
respectivamente). A c´elula 9 (022301md01) tamb´em foi exclu´ıda da an´alise, por n˜ao apresentar quaiquer amostras para um dos est´ımulos, de maneira que das 29 c´elulas com registros simultˆaneos de PSPs e LFPs, 25 foram mantidas para c´alculo de informa¸c˜ao considerando os dois tipos de registros. Todas as c´elulas s˜ao utilizadas, no entanto, quando se trata de estimar informa¸c˜ao utilizando apenas PSPs ou apenas LFPs.
5.4
Obten¸c˜
ao de Amplitudes e Latˆ
encias
O passo seguinte na an´alise foi a retirada de aspectos dos registros a fim de computar
a informa¸c˜ao m´utua entre est´ımulos e respostas eletrofisiol´ogicas. Esta etapa reduz os
valores poss´ıveis de respostas, reduzindo um sinal no tempo para apenas um ou dois escalares. Os escalares escolhidos foram a amplitude, computada como a diferen¸ca entre os valores m´aximo e m´ınimo do sinal, e a latˆencia, computada como a dura¸c˜ao temporal da apresenta¸c˜ao do est´ımulo ao m´aximo do sinal, no caso de PSPs, ou ao m´ınimo do sinal, no caso de LFPs. A Figura 5.8 ilustra esses parˆametros, o intervalo para sua obten¸c˜ao foi de 85ms (nos registros, desde a apresenta¸c˜ao do est´ımulo em 15ms at´e 100ms).
0 0.02 0.04 0.06 0.08 0.1
−5 0 5 10 15 20 25 30 35
Potencial (unidades normalizadas)
Tempo (s)
PSP
0 0.02 0.04 0.06 0.08 0.1
−
−0.25
−
−0.15 −0.1 −0.05
0 0.05
0.3 0.2
Tempo (s)
LFP
Figura 5.8: Obten¸c˜ao de amplitudes e latˆencias retiradas de registros da c´elula 12
(030201md01b) durante o est´ımulo de 3003Hz (latˆencia em azul e amplitude em verme-lho).
´
Cap´ıtulo 6
Resultados
Neste cap´ıtulo computamos a informa¸c˜ao dos dados j´a tratados com os m´etodos dos cap´ıtulos 4 e 5. Para cada c´elula, optou-se por realizar uma discretiza¸c˜ao simples das amplitudes ou latˆencias das respostas de PSP/LFP em 4 classes equipovoadas. A ideia foi comparar as estimativas obtidas de registros de natureza distinta, em particular PSPs e LFPs, em seus diferentes aspectos, i.e., amplitudes e latˆencias, e ver quais s˜ao mais in-formativas sobre os est´ımulos sonoros. Ainda verificaram-se poss´ıveis rela¸c˜oes de sinergia ou redundˆancia.
6.1
Caracteriza¸c˜
ao das amplitudes e latˆ
encias de PSPs
e LFPs
Antes de estimar a informa¸c˜ao m´utua em si, realizamos um estudo de caracteriza¸c˜ao
8 9 10 0
2 4 6 8 10 12 14 16 18 20 PSP
Frequência (ln Hz)
Amplitude Média
8 9 10
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Frequência (ln Hz) LFP
A B
Figura 6.1: Exemplos de curvas de caracteriza¸c˜ao de amplitudes, c´elula 13. Barras
indicam amplitudes m´edias, de PSPs (A) ou LFPs (B), encontradas para cada frequˆencia de est´ımulo (cujo logaritmo natural est´a indicado no eixo horizontal).
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
17 16
11 21
24
1 2 3
4 19
22
18
23 27
28
8 5 6
7
9 10 13
14 15
C
élula
A
mplitude M
édia Nor
malizada por C
élula
26 20 31
33
25
12
29
32 30
2000 H z
3003 H z
4993 H z
8300 H z
13799 H
z 22939 H
z 38134 H
z
PSP
A
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
17 20 26 29 16 25 11 21 24 1 2 3
4 19 12 22 18 23 27 28
8 5 6 7 9 10 13 14 15
C
élula
A
mplitude M
édia Nor
malizada por C
élula
2000 H
z 3003 H
z 4993 H
z 8300 H
z 13799 H
z 22939 H
z 38134 H
z Frequência
B
LFP
Figura 6.2: Curvas normalizadas para todas as c´elulas (amplitudes). Para cada
c´elula, a amplitude m´edia da resposta de PSP/LFP foi normalizada pela amplitude m´axima. A ordem das c´elulas foi rearranjada de acordo com o est´ımulo de maior resposta.
De posse dos resultados de para quais est´ımulos, na m´edia, as c´elulas apresentam
mai-ores amplitudes, foi poss´ıvel construir histogramas que contam o n´umero c´elulas segundo
sua frequˆencia de amplitude m´axima (Figura 6.3). ´E interessante ressaltar, no entanto,
20000 10000 46731 1
2 3
20000 10000 46731
1 2 3 4 5 6 7 8
Frequência
Número de células
Frequência PSP
A
LFP
B
Figura 6.3: Distribui¸c˜ao de est´ımulos de maior amplitude. Os gr´aficos mostram
histogramas indicando o n´umero de c´elulas cujo PSP/LFP teve maior amplitude para a
frequˆencia indicada no eixo horizontal (em escala logar´ıtmica).
8 9 10 0
0.005 0.01 0.015 0.02 0.025 0.03 0.035
Frequência (ln Hz)
Latência Média (s)
8 9 10
0 0.01 0.02 0.03 0.04 0.05 0.06
Frequência (ln Hz)
PSP LFP
A B
Figura 6.4: Exemplos de curvas de latˆencias, c´elula 13. Barras indicam latˆencias
2000 H z
300 3 H
z 499
3 H z
830 0 H
z 13799
Hz
22939 H z
38134 H
z Frequência
17
16
11 21
24
1 2 3
4 19
22 18 23
27
28 8
5 6
7 9
10
13 14 15 26
20 25
12 29
C
élula
B LFP
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
L
a
tência M
édia Nor
malizada por C
élula
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
L
a
tência M
édia Nor
malizada por C
élula
PSP A
2000 H z
300 3 H
z 499
3 H z
830 0 H
z 13799
Hz
22939 H z
38134 H
z 17
16 11
21 24
1 2 3
4 19 22
18 23
27 28
8 5 6
7 9
10
13 14 15
C
élula
26
20 31
33 25 12
29
32 30
Figura 6.5: Curvas normalizadas para todas as c´elulas (latˆencias). Para cada
c´elula, a latˆencia m´edia da resposta de PSP/LFP foi normalizada pela latˆencia m´axima. A ordem das c´elulas foi rearranjada de acordo com o est´ımulo de menor latˆencia.
Tamb´em para o caso de latˆencias foi poss´ıvel construir histogramas que contam o
n´umero c´elulas segundo sua frequˆencia de latˆencia m´ınima (Figura 6.6). Observa-se que
![Figura 2.1: Exemplo de aparelho de registro simultˆaneo de PSP por patch clamp e de LFP por eletrodo, in vivo (imagem retirada da referˆencia [23]).](https://thumb-eu.123doks.com/thumbv2/123dok_br/15623668.108197/19.892.195.698.107.557/figura-exemplo-aparelho-registro-simultˆaneo-eletrodo-retirada-referˆencia.webp)