Leaving the past behind.
Texto
Imagem
Documentos relacionados
NÓS E OS OUTROS EM/COM QUEM HABITAMOS Na atual sociedade do conhecimento, pautada por encontros inesperados com as mudanças da era digital e cimentada por múltiplas incertezas,
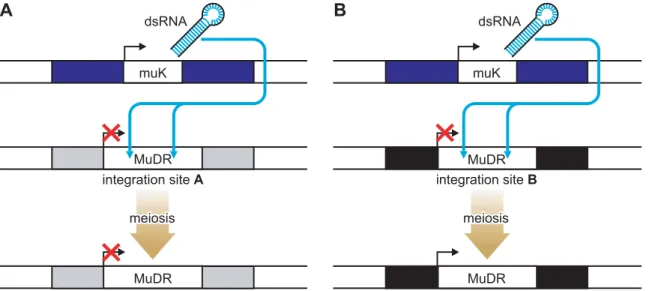
It is likely that the former process occurs in the G 1 phase of the cell cycle while the latter process must occur no later than during the G 2 phase (Golubovskaya et al.,
The main factors we identified are organised into four groups of variables: civil society (f)actors such as the Portuguese print media and key opinion makers; state actors, such as
[r]
Face ao exposto, esta revisão bibliográfica narrativa tem como objetivo aprofundar o estudo da reabilitação protética com implantes dentários, nomeadamente ao nível
As a storage tissue, the mature endosperm accumulates compounds that will serve as nutrient reserves that are mobilized by the embryo during the germinative
O objetivo desta tese será mostrar a utilidade das técnicas de Data Mining (DM) para conse- guir prever Efeitos Adversos de Medicamentos (EAM) que advém da interação entre
During early development of the tick embryo, glycogen degradation occurs while during the late stages of embryogenesis glycogen reserves are resynthesized and protein
