Flávio Freitas Barbosa
Diferenças funcionais das regiões hipocampais na formação da memória do tipo “o quê", "quando" e "onde" em ratos
Tese apresentada à Universidade Federal do
Rio Grande do Norte, para obtenção do título
de Doutor em Psicobiologia.
Natal
Flávio Freitas Barbosa
Diferenças funcionais das regiões hipocampais na formação da memória do tipo “o quê", "quando" e "onde" em ratos
Tese apresentada à Universidade Federal do
Rio Grande do Norte, para obtenção do título
de Doutor em Psicobiologia.
Orientadora: Dra. Regina Helena da Silva
Natal
Agradecimentos
Gostaria de agradecer inicialmente aos agentes financiadores CNPq e
CAPES que permitiram a realização da pesquisa ao longo do meu doutorado.
Agradeço especialmente a minha orientadora a professora Dra. Regina
Helena da Silva pela paciência e ensinamentos nesses últimos quatro anos. Muito
obrigado!!!
Gostaria também de agradecer especialmente a professora Dra. Alessandra
Mussi Ribeiro e a Dra. Alicia Cabral pelos valiosos ensinamentos nas cirurgias
estereotáxicas, passo essencial na minha tese. Aos professores Dr. Sidarta Ribeiro
e Dra. Elaine Gavioli agradeço pelas ricas discussões que tivemos durante todo o
processo.
Para a realização desta pesquisa foi também de fundamental importância a
colaboração de todos os meus colegas de laboratório e em especial: Isabella;
Hermany; Luciana; Priscila. Muito obrigado pelo suporte.
Gostaria de agradecer especialmente a todos os meus amigos e familiares
que me apoiaram durante esses quatro anos. Principalmente aos meus pais que
sempre me incentivaram e ainda me incentivam. E também a minha esposa e amiga
Resumo
A memória episódica refere-se à capacidade de recordar quando e onde um
determinado evento ocorreu. O hipocampo é uma estrutura chave para esse sistema
de memória e diversos estudos teóricos têm sugerido que o giro denteado (GD) e
CA3 estão envolvidos na aquisição rápida da memória episódica, enquanto a
sub-região CA1 estaria envolvida na separação temporal de diferentes episódios.
Contudo, há poucos estudos em modelos animais com tarefas que acessem os
aspectos ―o que‖, ―quando‖ e ―onde‖ simultaneamente. Recentemente, uma tarefa de reconhecimento de objetos em roedores que avalia a memória similar a episódica foi
desenvolvida. A tarefa consiste em duas sessões de treino e uma de teste, cada
uma com 5 minutos de duração. Na primeira sessão de treino o rato é colocado em
uma arena familiar com quatro objetos idênticos (A), após uma hora o rato é
reexposto ao campo com outras cópias (B). O teste é realizado 1 h depois, e o
animal é apresentado a dois objetos da segunda e dois objetos da primeira
exposição, sendo que um dos objetos A está uma nova localização no campo
aberto. Espera-se que o objeto mais antigo e deslocado seja o mais explorado. O
objetivo deste estudo foi avaliar o papel das sub-regiões hipocampais na aquisição
da memória similar a episódica em ratos. Inicialmente, avaliamos a capacidade de
ratos Wistar evocarem essa tarefa após 24 h de retenção, uma vez que a tarefa foi
desenvolvida inicialmente com um intervalo de 1h. De fato, os animais conseguiram
discriminar a localização e a ordem de apresentação dos objetos após um intervalo
de 24h. Além disso, a administração de escopolamina (1 mg/kg, ip) imediatamente
após o treino prejudicou o desempenho dos animais, o que favoreceu a validação
o efeito da inativação temporária do giro denteado/CA3 e de CA1 na aquisição desta
tarefa. Muscimol, um agonista gabaérgico (0,250 µg/µl; volume = 0,5 µl), ou solução
salina no mesmo volume foram injetados nessas sub-regiões quinze minutos antes
da primeira sessão de treino. A inativação pré-treino do GD/CA3 prejudicou a
discriminação espacial dos objetos, enquanto que a inativação de CA1 levou a
exploração igual dos objetos, independentemente da localização ou ordem de
apresentação. Estes resultados corroboram os modelos teóricos, indicando um
papel importante de GD/CA3 na aquisição rápida de memórias episódicas, assim
como na separação de padrões espaciais, enquanto a sub-regiãoCA1 estaria
Abstract
Episodic memory refers to the recollection of what, where and when a specific
event occurred. Hippocampus is a key structure in this type of memory.
Computational models suggest that the dentate gyrus (DG) and the CA3
hippocampal subregions are involved in pattern separation and the rapid
acquisition of episodic memories, while CA1 is involved in memory
consolidation. However there are few studies with animal models that access
simultaneously the aspects ‗what-where-when‘. Recently, an object recognition episodic-like memory task in rodents was proposed. This task
consists of two sample trials and a test phase. In sample trial one, the rat is
exposed to four copies of an object. In sample trial two, one hour later, the rat
is exposed to four copies of a different object. In the test phase, 1 h later, two
copies of each of the objects previously used are presented. One copy of the
object used in sample trial one is located in a different place, and therefore it is
expected to be the most explored object.However, the short retention delay of
the task narrows its applications. This study verifies if this task can be evoked
after 24h and whether the pharmacological inactivation of the DG/CA3 and
CA1 subregions could differentially impair the acquisition of the task
described. Validation of the task with a longer interval (24h) was accomplished
(animals showed spatiotemporal object discrimination and scopolamine (1
mg/kg, ip) injected pos-training impaired performance). Afterwards, the GABA
agonist muscimol, (0,250 µg/µl; volume = 0,5 µl) or saline were injected in the
hippocampal subregions fifteen minutes before training. Pre-training
inactivation of the DG/CA3 subregions impaired the spatial discrimination of
preserved. Rats treated with muscimol in the CA1 subregion explored all the
objects equally well, irrespective of place or presentation time. Our results
corroborate the computational models that postulate a role for DG/CA3 in
spatial pattern separation, and a role for CA1 in the consolidation process of
Sumário
Introdução ... 1
Memória episódica em humanos ... 2
Memória similar à episódica em animais ... 3
Mecanismos neurais ... 5
Justificativa ... 8
Objetivo Geral ... 9
Objetivos específicos ... 9
Primeiro artigo: objetivo específico 1 ... 10
Extending possible applications of an episodic-like memory task in rats ... 10
Segundo artigo: objetivo específico 2 ... 12
Differential role of hippocampal regions for ―what‖, ―where‖ and ―when‖ memory in rats ... 12
Conclusão Geral ... 44
1 Introdução
A compreensão dos processos mentais e dos mecanismos neurais
subjacentes à memória é uma das principais áreas da neurociência moderna.
Entender como funcionam os diferentes processos mnemônicos e quais estruturas
encefálicas permitem essa função é um dos principais desafios do campo. Há muito
que a memória não é vista como um sistema unitário. A sua primeira classificação foi
quanto ao tempo de retenção, dividindo-se em memória de curto-prazo e de
longo-prazo. A primeira caracteriza-se por eventos que podem ser recordados por até
poucos minutos, enquanto a segunda se refere a uma memória de horas até toda a
vida (Squire e Zola, 1996; Eichenbaum, 2000; Tulving, 2001; 2002). Uma das
classificações mais utilizadas é a que divide a memória em dois grandes sistemas:
memória declarativa ou explícita; e memória não-declarativa ou implícita (Squire e
Zola, 1996).
A memória explícita refere-se ao sistema mnemônico em que a evocação de
um evento pode ser declarada de forma consciente pelo sujeito (Eichenbaum, 2000).
A consolidação desses eventos em tal sistema depende de estruturas do lobo
temporal medial, ou seja, do hipocampo e dos córtices adjacentes: entorrinal;
perirrinal e parahipocampal (O'reilly e Norman, 2002; Simons e Spiers, 2003;
Eichenbaum, 2004). A memória implícita é evocada inconscientemente, sendo
expressa através do desempenho em uma determinada tarefa. Consistem nos
nossos hábitos, habilidades e aprendizagem associativa e não associativa,
funcionando independentemente daquelas mediadas pelo lobo temporal medial, e
envolvendo outras estruturas neurais, como a amígdala, o estriado e o cerebelo
2 De acordo com Tulving (2001, 2002), a memória explícita pode ser dividida
em dois outros sistemas: memória semântica e episódica. A primeira caracteriza-se
pela evocação de fatos, como, por exemplo, a evocação de que a capital do Brasil é
Brasília. Na concepção de Tulving (2001, 2002), a evocação de um fato da memória
semântica não depende de uma ―viagem no tempo‖, enquanto a memória episódica
evoca eventos que podem ser localizados no tempo, sendo possível determinar
―onde‖ e ―quando‖ ele ocorreu. Ou seja, a memória episódica consiste na formação e
evocação de eventos específicos vivenciados pelo organismo.
Memória episódica em humanos
A memória episódica consiste na recuperação de um evento através de uma
ou várias associações feitas sobre aquele evento, ou seja, não apenas o ―quê‖ é resgatado, mas também o ―onde‖ e o ―quando‖. Além disso, é necessário fazer uma
viagem temporal mental até o evento. Para tanto é preciso que o organismo seja
capaz de estabelecer cognitivamente o tempo subjetivo, a consciência autonoética e
a autoconsciência (Tulving 2002).
O tempo subjetivo caracteriza-se pela capacidade do organismo em formar
conceitos sobre o que seja passado, presente e futuro. Sem essa capacidade não
há como realizar a viagem mental através do tempo. A consciência autonoética, por
sua vez, consiste na capacidade do indivíduo em se reconhecer como o mesmo
indivíduo, seja no passado ou no presente, ou ainda, capaz de se projetar no futuro,
ou seja, capaz de fazer planos sobre o que vai fazer em um futuro próximo ou
distante. E por último, para a realização da viagem é necessário um viajante, neste
caso o indivíduo precisa ter consciência de si mesmo como uma entidade diferente
3 Tulving (2002) sugeriu que apenas o ser humano seria capaz de realizar esta
viagem temporal mental.
Memória similar à episódica em animais
Existe um intenso debate na literatura sobre a possível capacidade dos
animais também realizarem uma viagem mental através do tempo, ou seja, se
apenas os seres humanos são capazes de realizar essa tarefa cognitiva (Dere,
Kart-Teke et al., 2006; Crystal, 2009). Parece haver alguns indícios de que algumas
espécies de primatas não-humanos e de aves (―scrub jays‖) são capazes de realizar planejamentos futuros (Clayton, Bussey et al., 2003; Crystal, 2009). Todavia, esse
dilema pode ser parcialmente evitado ao se separar os critérios fenomenológicos
dos comportamentais. Clayton e colaboradores (2003) sugeriram, então, denominar
esse tipo de memória de similar a episódica, em animais quando apenas os critérios
comportamentais (contexto espaço-temporal) são atendidos.
Há um esforço recente em se criar uma tarefa na qual os aspectos ―o quê‖, ―onde‖ e ―quando‖ sejam acessados (Ergorul e Eichenbaum, 2004; Dere, Huston et
al., 2005a; Dere, Huston et al., 2005b; Eichenbaum, Fortin et al., 2005; Babb e
Crystal, 2006). Alguns desses protocolos conseguem abordar alguns desses
aspectos ou até mesmo todos eles. Babb & Crystal (2006) conseguiram elaborar um
protocolo que avalia todos esses aspectos, no entanto a tarefa envolve um reforço e
necessitam muitas tentativas para a ocorrência da aprendizagem, o que poderia
levar à formação de uma memória semântica (O'reilly e Rudy, 2001; Dere, Kart-Teke
et al., 2006; Crystal, 2009). Recentemente, foi elaborado um protocolo que não
apresenta muitas séries e que envolve um comportamento natural dos roedores que
4 reconhecimento de objetos em camundongos, conseguindo assim acessar os
aspectos ―o quê‖, ―quando‖ e ―onde‖ da memória episódica. Esse protocolo consiste em duas fases de treino e a fase de teste. Na primeira fase, o camundongo é
colocado no centro de um campo aberto onde estão presentes quatro cópias de um
mesmo objeto (A), dispostos em uma configuração triangular. Após um intervalo de
50 min, o camundongo é colocado novamente no centro do campo aberto, onde
desta vez quatro cópias de um objeto diferente estão presentes (B) dispostos em
uma configuração quadrangular. Após um intervalo de retenção de 50 min ocorre o
teste. No teste duas cópias do objeto A e duas cópias do objeto B estão no campo
aberto, sendo que um dos objetos da primeira amostra está em uma localização
diferente, enquanto o outro objeto está na mesma localização. Os dois objetos da
segunda amostra são colocados nas mesmas posições da fase de treino. Nesta fase
os objetos também estão organizados em uma configuração quadrangular. Nesse
modelo, durante o teste o animal explora mais os objetos antigos (As) do que os
objetos recentes (Bs). Além disso, o objeto antigo deslocado é mais explorado que
os demais objetos, levando os autores a concluir que a tarefa conseguiu acessar a
memória dos camundongos para ―o quê‖, ―quando‖ e ―onde‖, ou seja, contemplando
os aspectos da memória similar a episódica (Dere et al. 2005).
Esse modelo foi recentemente adaptado para ratos Wistar (Kart-Teke, Silva et
al., 2006; Kart-Teke, Dere et al., 2007), mostrando a viabilidade dessa tarefa
também para esses animais. Esse paradigma é bastante promissor para o estudo
memória similar a episódica, podendo ser usado para o estudo dos mecanismos
5 Mecanismos neurais
Os estudos dos mecanismos neurais da memória explícita tanto em animais
quanto com seres humanos vem mostrando que o lobo temporal medial e os córtices
associativos são estruturas essenciais (Scoville e Milner, 1957; Squire e Zola, 1996;
Wiltgen, Brown et al., 2004). O lobo temporal medial é essencial na consolidação da
memória explícita, no entanto não é o seu armazém final. O neocórtex envolvido
com o processamento da informação parece desempenhar esse papel. Por exemplo,
as áreas envolvidas com o processamento de informação visual são o sítio final de
uma memória explícita de conteúdo puramente visual (Wiltgen, Brown et al., 2004;
Frankland e Bontempi, 2005). O modelo padrão que explica como estas estruturas
estão envolvidas na formação e consolidação do traço mnemônico relata que a
informação, após ser processada nos córtices associativos, é retransmitida ao
hipocampo, onde o traço é mantido. O hipocampo então retransmitiria a informação
ao neocortéx, até o momento em que esse se tornaria independente do hipocampo
para evocação da informação (Mcclelland, Mcnaughton et al., 1995; Wiltgen, Brown
et al., 2004; Frankland e Bontempi, 2005).
Já está bem estabelecido na literatura que algumas das células piramidais do
hipocampo apresentam uma maior taxa de disparo quando o rato encontra-se em
um determinado local; esse tipo de célula denomina-se célula de lugar (Okeefe e
Dostrovsky.J, 1971; Okeefe e Recce, 1993). Essas células de lugar têm sido
estudadas extensivamente há mais de quarenta anos, no entanto a maioria dos
estudos é com tarefas que não apresentam demandas cognitivas (Ferbinteanu,
6 papel especial dessa estrutura na codificação de informação espacial da memória
episódica.
Um modelo computacional propôs o porquê dessas diferentes funções do
neocortéx e do hipocampo na aprendizagem e memória. McClelland et al. (1995)
sugerem que as funções complementares dessas estruturas decorrem da forma
como elas processam as informações. De acordo com o modelo teórico proposto por
eles, o neocortéx apresenta uma baixa taxa de aprendizagem, pois utiliza
representações distribuídas de forma sobrepostas, o que permite extrair aspectos
gerais do ambiente. Já o hipocampo, por utilizar representações distintas para
codificar os eventos, consegue aprender rapidamente sem sofrer interferência. Essa
capacidade de evitar interferência é fundamental para a memória episódica, que
requer a aprendizagem de eventos específicos do ambiente (Mcclelland,
Mcnaughton et al., 1995; Norman e O'reilly, 2003).
As sub-regiões do hipocampo responsáveis por conseguir codificar os
eventos em representações não sobrejacentes parecem ser o giro denteado e a
área CA3 (Eichenbaum 2004; O'reilly & Rudy 2001). É dito que essas regiões são
responsáveis por gerar a ―separação de padrões‖ (Leutgeb, Leutgeb et al., 2007), o que permite ao hipocampo evitar a interferência. Outra função importante do
hipocampo é conseguir evocar uma memória na presença de uma dica, mesmo que
essa dica não seja idêntica ao evento vivenciado anteriormente. Essa função é
chamada de ―completamento de padrões‖, sendo que nesse caso a área CA3 parece ser fundamental (O'reilly e Rudy, 2001; Guzowski, Knierim et al., 2004). Note
que essa função é conflitante com a anterior, já que, caso um determinado evento
seja julgado como antigo, isso levará a um completamento do padrão. Caso
7 em consideração os mais diversos dados da literatura, fica claro que o hipocampo é
uma estrutura fundamental na formação da memória episódica e que aparentemente
isso decorre de sua forma diferenciada de processar a informação, que permite a
8 Justificativa
Para estudar os mecanismos neurais da memória similar a episódica é
fundamental que se utilize um protocolo que realmente esteja acessando esse tipo de
memória. Apesar de se conhecer várias tarefas que dependam do hipocampo, poucas
parecem abordar o conceito de memória similar a episódica, ao menos nos aspectos de
―onde‖ e ―quando‖ um determinado evento (―o quê‖) ocorreu. Desse modo, o modelo
elaborado por Dere et al. (2005) parece englobar todos os pontos fundamentais que
caracterizam a memória similar a episódica, pois os animais tendem a explorar mais os
objetos da primeira sessão de treino em relação a segunda sessão, e mais o objeto
antigo deslocado que o antigo não-deslocado durante o teste. Uma vez que o tempo de
retenção é de aproximadamente 1 h, pode-se excluir a possibilidade da tarefa acessar a
memória operacional. Quanto ao conceito de memória episódica, parece impossível
que um único protocolo aborde todos os aspectos desse tipo de memória definida por
Tulving (2002), ou seja, ―o quê‖, ―quando‖ e ―onde‖, assim como também a consciência
autonoética, o tempo subjetivo e a autoconsciência. Sendo assim, esse protocolo
parece bastante interessante para estudar os mecanismos neurais da memória similar a
episódica, mais especificamente o papel do hipocampo na realização dessa tarefa e os
papéis do giro denteado e da área CA3. Tendo em vista que essas estruturas são
fundamentais para a capacidade do hipocampo em codificar eventos específicos do
9 Objetivo Geral
Avaliar a função das sub-regiões hipocampais em um protocolo de memória
similar a episódica baseada em reconhecimento de objetos em ratos.
Objetivos específicos
1) Verificar se a tarefa de memória similar a episódica proposta por Dere et al. (2005 a, b) pode ser evocada após um intervalo de retenção de 24 h;
2) Avaliar o desempenho dos animais no protocolo de memória episódica quando estes têm as sub-regiões hipocampais giro denteado/CA3 e CA1
10 Primeiro artigo: objetivo específico 1
Extending possible applications of an episodic-like memory task in rats
Flávio Freitas Barbosa, Isabella Maria de Oliveira Pontes, Alessandra Mussi Ribeiro,
Regina Helena Silva*
Memory Studies Laboratory, Physiology Department, Federal University of Rio
Grande do Norte, Natal, Brazil
* Corresponding author: Departamento de Fisiologia —Centro de Biociências — UFRN,
Av. Salgado Filho, s/n — Caixa Postal 1511, CEP 59078-970—Natal, RN, Brazil. Fax: +55 84 32119206.
E-mail address: [email protected] (R.H. Silva).
11 Resumo
Recentemente, uma tarefa similar a episódica baseada no reconhecimento de
objetos foi proposta para roedores. No entanto, o curto intervalo de retenção limita
as suas possíveis aplicações. Este estudo verificou se essa tarefa pode ser evocada
após 24h. Além disso, o efeito de uma agente amnésico clássico (escopolamina) foi
avaliado no processo de consolidação de memória similar a episódica. Os ratos
mostraram uma maior exploração dos objetos antigos em relação aos recentes,
assim como um maior tempo de exploração do objeto deslocado em relação ao
parado. Tanto a preferência temporal quanto espacial foi abolida pela administração
pós-treino de escopolamina (1 mg/kg ip), indicando prejuízo na evocação
espaço-temporal. Podemos concluir que a tarefa de reconhecimento de objetos que acessa
os componentes o quê, quando e onde da memória similar a episódica pode persistir
por 24h.
Palavras-chave: modelo animal; memória similar a episódica; reconhecimento de
12 Segundo artigo: objetivo específico 2
Differential role of hippocampal regions for “what”,“where” and “when” memory in rats
Flávio Freitas Barbosa1, Isabella Maria de Oliveira Pontes1, Sidarta Ribeiro2,3,
Alessandra Mussi Ribeiro1, Regina Helena Silva 1,2‘*
1. Physiology Department, Federal University of Rio Grande do Norte, Natal,
Brazil
2. Neuroscience Graduate Program, Federal University of Rio Grande do Norte
(UFRN), Natal, RN, Brazil
3. Edmond and Lily Safra International Institute of Neuroscience of Natal
(ELS-IINN), Rua Professor Francisco Luciano de Oliveira 2460, Bairro Candelária,
Natal, RN, Brazil
* Corresponding author: Departamento de Fisiologia —Centro de Biociências — UFRN,
Av. Salgado Filho, s/n — Caixa Postal 1511, CEP 59078-970—Natal, RN, Brazil. Fax: +55 84 32119206.
E-mail address: [email protected] (R.H. Silva).
13 Resumo
A memória episódica se refere à capacidade de recordar quando e onde um
determinado evento ocorreu. O hipocampo é uma estrutura chave para esse sistema
de memória e diversos estudos teóricos têm sugerido que o giro denteado (GD) e
CA3 estão envolvidos na aquisição rápida da memória episódica, enquanto a
sub-região CA1 estaria envolvida na separação temporal de diferentes episódios.
Contudo, há poucos estudos em modelos animais com tarefas que acessem os
aspectos ―o que‖, ―quando‖ e ―onde‖ simultaneamente. Recentemente, uma tarefa de
reconhecimento de objetos em roedores que avalia a memória similar a episódica foi
desenvolvida. A tarefa consiste em duas sessões de treino e uma de teste, cada
uma com 5 minutos de duração. Na primeira sessão de treino o rato é colocado em
uma arena familiar com quatro objetos idênticos, após uma hora o rato é reexposto
ao campo com outras cópias de um objeto diferente. O teste é realizado 24 h depois,
e o animal é apresentado a dois objetos da segunda e dois objetos da primeira
exposição, sendo que um dos objetos da primeira sessão está uma nova localização
no campo aberto. Espera-se que o objeto antigo e deslocado seja o mais explorado.
O objetivo deste estudo foi avaliar o papel das sub-regiões GD/CA3 e CA1 na
aquisição da memória similar a episódica em ratos. Muscimol, um agonista
gabaérgico (0,250 µg/µl; volume = 0,5 µl), ou solução salina no mesmo volume
foram injetados nessas sub-regiões quinze minutos antes do treino. A inativação
pré-treino do GD/CA3 prejudicou a discriminação espacial dos objetos, enquanto que a
inativação de CA1 levou a exploração igual dos objetos, independentemente da
localização ou ordem de apresentação. Estes resultados corroboram os modelos
14 memórias episódicas, assim como na separação de padrões espaciais, enquanto a
15 Abstract
Episodic memory refers to the recollection of what, where and when a specific event
occurred. Computational models suggest that the dentate gyrus (DG) and the CA3
hippocampal subregions are involved in pattern separation and the rapid acquisition
of episodic memories, while CA1 is involved in memory consolidation. Most of the
studies carried out to test this hypothesis failed to simultaneously address the ‗what‘, ‗where‘ and ‗when‘ aspects of episodic memory. Recently, an episodic-like memory
task based on object recognition was validated in rats. This task consists of two
sample trials and a test phase. In sample trial one, the rat is exposed to four copies
of an object. In sample trial two, one hour later, the rat is exposed to four copies of a
different object. In the test phase, 24 h later, two copies of each of the objects
previously used are presented. One copy of the object used in sample trial one is
located in a different place, and therefore it is expected to be the most explored
object. The goal of this study was to evaluate whether the pharmacological
inactivation of the DG/CA3 and CA1 subregions could differentially impair the
acquisition of the task described. Animals in the control group showed spatiotemporal
object discrimination. Inactivation of the DG/CA3 subregions impaired the spatial
discrimination of the objects (‗where‘), while the temporal discrimination (‗when‘) was preserved. Rats treated with muscimol in the CA1 subregion explored all the objects
equally well, irrespective of place or presentation time. To our knowledge, this is the
first study to evaluate the role of these hippocampal subregions in the acquisition of
an episodic-like memory task. Our results corroborate the computational models that
postulate a role for DG/CA3 in spatial pattern separation, and a role for CA1 in the
16 Introduction
Human episodic memory has been conceptualized as the recollection process
of ‗what‘, ‗where‘ and ‗when‘ a specific event occurred. Tulving pointed out that this
kind of memory is unique to humans, since it involves self-awareness, autonoetic
consciousness and a subjective sense of time (Tulving, 2001; Tulving, 2002).
However, in the past few years, a growing body of evidence has corroborated the
notion that non-human animals can retrieve the spatiotemporal content of an
episode. In studies with birds, Clayton and collaborators were able to distinguish
phenomenological and behavioral aspects (Clayton et al., 2003). The behavioral
criteria consist of the ‗what-where-when‘ content of a memory, and for this reason these authors designated this kind of memory as episodic-like memory. In rodents,
Dere and collaborators developed a new object recognition task that simultaneously
asks where and when a specific object was found previously in a familiar arena,
allowing the evaluation of the ‗what-where-when‘ aspects of episodic memory (Dere et al., 2005a; Dere et al., 2005b; Dere et al., 2006). This task has some advantages
when compared to other episodic-like memory tasks. Firstly, it accesses a natural
rodent behavior, i.e. the exploration of novel objects detected in a familiar
environment. Thus, the task does not involve extensive training. Secondly, different
mnemonic processes such as acquisition, consolidation and retrieval, can be easily
separated. The task developed by Dere and colleagues has an interval of just one
hour between the second sample trial and the test. Recently, we have shown in
Wistar rats that this task also works when a 24 h retention delay is employed
(Barbosa et al., 2010). This greatly extends the possible applications of the task,
including pharmacological manipulations, since most drugs can be active for a few
17 The role of the hippocampus in the formation of episodic memory has been
well established (Scoville and Milner, 1957; Tulving, 2002). Specifically, lesion
studies, both in mice and rats, have demonstrated a role for the hippocampus in the
object recognition episodic-like memory task mentioned above (DeVito and
Eichenbaum, 2010; Good et al., 2007; Li and Chao, 2008). Lesioned animals do not
show spatiotemporal discrimination of objects, which is in agreement with the
symptoms presented by patients after surgical hippocampal ablation (Manns and
Eichenbaum, 2006; Tulving, 2002). In addition to this general hippocampal role of
storing new episodes, some theoretical models suggest that the hippocampus is
crucial to avoid catastrophic interference between similar episodes i.e., this structure
can rapidly acquire new detailed events and store them separately (McClelland et al.,
1995; Norman and O'Reilly, 2003). In this respect, different hippocampal subregions
seem to have different functions in the processing of new information. It has been
proposed that the dentate gyrus (DG) and CA3 are essential for the separation of
similar episodes/patterns (Hunsaker and Kesner, 2008; O'Reilly and Norman, 2002;
Rolls, 2010; Rolls and Kesner, 2006). Interestingly, place cells in the DG have
smaller place fields than CA3 place cells (Jung and McNaughton, 1993), which could
explain the different roles of these subregions in the processing of spatial
information. Additionally, Hunsaker & Kesner (2008) have showed that rats with
bilateral DG lesions had a poor performance in a temporal order task for spatial
locations with high (but not low) spatial interference. This result corroborates the idea
that the DG is crucial for separating episodes with high spatial overlapping.
The dorsal CA1 field is the major output from the hippocampus to the
neocortex, and is the target of two different inputs from CA3 and entorhinal cortex.
18 CA3 and projects back to the neocortex. Therefore, CA1 has been suggested to play
an important role in the consolidation of different episodes. Additionally, CA1 lesion
studies have demonstrated a role in the temporal separation of sequential events,
such as odors and visual objects (Hoge and Kesner, 2007; Kesner and Hunsaker,
2010).
It is important to note that most of these studies did not use protocols that
accessed simultaneously the ‗what-where-when‘ aspects of an episode. Therefore, it is not yet clear what is the role of these subregions in the acquisition of an
episodic-like memory task. Furthermore, most of the studies regarding the functions of the
hippocampal subregions used permanent lesions, which preclude the investigation of
the temporary role of these structures in the formation of episodic-like memories.
Thus, the goal of the present study was to evaluate the effects of a temporary
inactivation of DG/CA3 or CA1 on the rapid encoding of spatiotemporal information
related to different objects. We found that the inactivation of DG/CA3 impaired the
spatial - but not the temporal - discrimination of objects. This result confirms previous
results that pointed to a role of these structures in fine spatial pattern separation. On
the other hand, the inactivation of CA1 impaired both spatial and temporal
discrimination, which is in agreement with the notion that CA1 is involved in the
long-term consolidation of episodic-like memories.
Materials and Methods
Animals
Thirty-nine 3-month old male Wistar rats (weighing 250-350 g) were used in
this study. Animals were housed under conditions of controlled temperature (25 1
19 libitum throughout the experiment. Rats were handled in accordance with the
guidelines of the Brazilian Society for Neuroscience & Behaviour for the use of
animals in research, and all procedures were approved by the local ethical
committee. All efforts were made to minimize animal pain, suffering or discomfort.
Surgery
Animals were randomly assigned to one of four groups: DG/CA3 (n = 13); CA1
(n = 11); DG/CA3 control (n = 7) or CA1 control (n = 8). Rats were anesthetized with
an intraperitoneal injection of ketamine (100 mg/kg) and xylazine (50 mg/kg). Next,
animals were positioned in a stereotaxic apparatus (Insight, Brazil) and the skull was
exposed. Stainless guide cannulas (25 gauge, 12 mm) were implanted bilaterally in
the dorsal DG/CA3 or in the dorsal CA1 hippocampus subregions. The stereotaxic
coordinates from bregma (Paxinos and Watson, 2009) for guide cannula placement
in the dorsal DG/CA3 were: anterior-posterior (AP) = -4.3 mm, medial-lateral (ML) = ±
2.0 mm, and dorsal-ventral (DV) = - 2.0 mm. The dorsal CA1 coordinates were:
anterior-posterior (AP) = -4.3 mm from bregma, medial-lateral (ML) = ± 2.5 mm, and
dorsal-ventral (DV) = - 1.5 mm. The guide cannula tips were placed 1 mm above the
injection site to minimize lesion in the area of interest. Guide cannulas were
anchored to the skull with small stainless steel screws and dental acrylic. At the end
of the surgery each cannula was temporarily sealed with a stainless steel wire to
prevent obstruction. After surgery, animals received an anti-inflammatory (diclofenac
sodium 75 mg/ml, i.m.) and an antibiotic (penicillin 60.000 UI/ml, i.m). Animals were
given 2 days of post-operative recovery prior to the start of the handling procedure.
The GABAA agonist muscimol (Sigma) was used to temporally inactivate the
hippocampal subregions GD/CA3 or CA1. Bilateral infusions of muscimol (0.25 µg/µl
20 microsyringe pump (Insight, Brazil) using 10 µl Hamilton syringes connected to
polyethylene tubing. Injection needles were left in the guide cannula for an additional
60 s following the infusions to allow for diffusion of the drug from the needle tip.
Behavioral tests were conducted 15 min after drug infusion.
Apparatus and objects
The behavioral tests were conducted in a circular open field (84 cm in
diameter, wall height 32 cm), made of wood and painted in black. External cues
(geometric forms) were placed in the room in order to facilitate self-location in the
open field. Three sets of objects, with four copies each, were used randomly among
experimental subjects. The objects used were a sugar bowl, a mug and a goblet. The
objects differed in height (9-12 cm), width (6-10 cm) and colour. All the objects were
made of plastic and filled with cement to ensure that animals could not displace
them. The apparatus was cleaned with a 5% alcohol solution after each behavioural
session. The sessions were recorded by a digital camera placed above the
apparatus and the behavioural parameters were registered by an animal tracking
software (Anymaze, Stoelting, USA). The behavioural sessions were monitored
through a computer screen placed in another room.
Episodic-like memory task
All rats were handled for 20 min/day for 5 days. Afterwards, animals were
submitted to a ten-minute habituation session in the open field (without the objects),
24h before the beginning of the object recognition episodic-like memory task. The
behavioral procedure was the same as described earlier (Barbosa et al. 2010).
Briefly, the task consisted of two sample trials and one test trial (figure 1). In the first
sample trial, each animal were placed in the open field with four identical copies of
21 in the arena with four objects B in quadrangular disposition (second sample trial).
Finally, test trial was performed 24h later. In this phase, two copies of the objects
from each trial were presented. One of the objects from the first trial was in a different
spatial location. The other objects were in the same positions of the sample trials. In
the test trial the rats are expected to spend more time exploring the two copies from
the first trial (A) compared to the two copies from the second trial (B). This preference
for the old over the more recent objects would account for the ―when‖ component of
episodic-like memory. In addition, the rats are also expected to spend more time
exploring the displaced object ―A‖ compared to the stationary object ―A‖, which would correspond to the ―where‖ component. DG/CA3 and CA1 animals and control group
received muscimol and saline, respectively, fifteen minutes before the first sample
trial.
Histological methods
Rats were deeply anesthetized with 1 ml of thiopental sodium (25 mg/ml) and
perfused intracardially with 0,9% saline, followed by 10% formol-saline solution.
Methylene blue (0,5 µl) was injected in the guide cannulas just before craniotomy.
The brains were removed and stored in 10% formol-saline for 24 h, and then stored
in 30% sucrose at 4 ºC for at least 72 h before being frozen and sectioned at 50 µm
with a cryostat. Sections were mounted on glass slides and then Nissl stained with
neutral red for verification of the exact placement, at the microscope, of the cannula
and the infusion needle tip.
Data collection and statistical analyses
Videos recorded by the animal tracking software were used to measure the
duration of object exploration. An experimenter, blind to group assignment, used
22 actively had physical contact with it. The dependent variable used was object
exploration ratio, which was calculated for each object separately (time exploring a
specific object / total time exploring all objects). ―Old familiar‖ object exploration ratio (time exploring both ―old familiar‖ objects/ total time exploring all objects) and ―recent familiar‖ object exploration ratio (time exploring both ―recent familiar‖ objects/ total
time exploring all objects) were also calculated. A two-way ANOVA with repeated
measures, with different groups as the between effect and objects exploration ratio
as the within effect, was carried out to the whole five minutes, as well to the first
minute of the test session. A previous study of our group has shown that control
animals spend more time exploring the old familiar-displaced object only in the first
minute of the test session (Barbosa et al. 2010). A priori planned paired two-tailed t
test was applied to compare ―old familiar‖ objects exploration ratio with ―recent familiar‖ object exploration ratio. Furthermore, a t test for dependent groups was carried out comparing ―old familiar‖ stationary object exploration ratio with recent
object exploration ratio mean to guarantee that successful recency discrimination in
these analyses could not be consequence of the displaced object exploration. The
―where‖ episodic-like memory aspect was assessed comparing the old displaced
object exploration ratio with the old stationary object. All these comparisons were run
separately for each group.
Two-way repeated ANOVA, with different groups as the between effect and
the sessions as the within effect, was run to analyze possible differences in the
locomotor activity. The total time exploring the four objects throughout the trials was
also compared by a two-way repeated ANOVA, with different groups as the between
23 post hoc was performed. In all statistical tests, rejection of the null hypothesis was
set at p < 0.05.
Results Histology
Only rats with correct cannula guide placement and infusion needle tip location
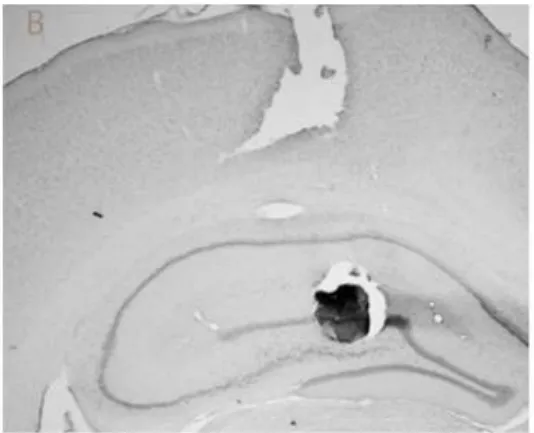
were behaviorally analyzed. Figure 2 shows two examples of microinjections located
in the dorsal CA1 and dorsal DG/CA3. As one can see, it is unlikely that muscimol
microinjected in the DG did not spread to the CA3 subregion. However, methylene
blue injected in dorsal CA1 did not reach DG/CA3, and vice-versa.
Behavioral Results
Locomotor activity and motivation
Saline DG/CA3 and saline CA1 animals did not show significant differences in
any of the dependent variables analyzed (data not shown). For this reason, these
groups were pooled together in the analyses. Two-way ANOVA with repeated
measures, with different groups as the between-subject factor and the sessions as
the within-subject factor, revealed a main effect of distance travelled across the trials
[F(2,36) = 4.63; p = 0.015]. Bonferroni post test showed that rats travelled a longer
distance in the first sample trial when compared to the second trial (p = 0.004, fig. 3).
We found no main effect of treatment [F(2,36) = 1.30; p = 0.28], and no interaction
treatment x distance travelled [F(4,36) = 0.32; p = 0.86]. A two-way ANOVA with
different groups as the between-subject factor and objects exploration time as the
within-subject factor showed no main effect of treatment [F(2,36) = 1.22; p = 0.30] or
24 group x object exploration interaction was found [F(4,36) = 3.63; p = 0.01]. To further
investigate this interaction, separate one-way ANOVAs with repeated measures were
applied to each treatment group, but no significant effect was found (Fs > 0.13; p >
0.05).
Episodic-like memory task
First, a two-way ANOVA with repeated measures, with different groups as the
between-subject factor and objects exploration ratio as the within subject factor, was
applied in the analysis of the first minute of the test session. There was no significant
group effect [F(2,36) = 0.72; p = 0.49], but we detected a significant objects
exploration ratio effect [F(3,36) = 8.00; p < 0.001], as well as a significant group x
objects exploration ratio interaction [F(6,36) = 2.75; p = 0.01; fig 4]. A priori planned
paired two-tailed t tests were applied to each group. Saline rats spent more time
exploring ―old‖ when compared to ―recent‖ objects (t (14) = 3.09; p = 0.008).
Additionally, no difference was found between old-familiar stationary object
exploration and the means of both ―recent familiar‖ objects exploration ratios. As expected, saline rats spent more time exploring the old-familiar displaced object that
the old-familiar stationary object (t (14) = 3.32; p = 0.005). Finally, saline rats did not
discriminate between the two ―recent‖ objects (p > 0.6). When the same comparison
was applied to CA1 rats, we found that rats explored all the objects at similar rates (p
> 0.05). However, GD/CA3 rats were able to temporally discriminate the objects,
since they spent more time exploring ―old‖ objects than ―recent‖ objects (t (12) = 5.56;
p < 0.001). They also explored more the old-familiar stationary object than the means
of both ―recent familiar‖ objects. On the other hand, they explored old-familiar
25 Additionally, GD/CA3 rats did not discriminate between the two ―recent‖ objects (p >
0.05).
A two-way repeated ANOVA, with different groups as the between effect and
objects exploration ratio as the within effect, was also applied to the whole test
session. There was no significant group effect [F(2,36) = 2.38; p = 0.10; fig 5] or
group x objects exploration ratio interaction [F(6,36) = 0.54; p = 0.77], but a
significant objects exploration ratio main effect was found [F(3,36) = 5.67; p = 0.001].
A priori planned paired two-tailed t tests were also applied to each group. Saline rats
explored more ―old‖ objectsthan ―recent‖ objects (t(14) = 4.68; p < 0.001), and spent
more time exploring old-familiar stationary than the means of both ―recent familiar‖ object exploration ratios (t(14) = 4.66; p < 0.001). This group did not discriminate
between old-familiar displaced object and old-familiar stationary object (t(14) = 1.39;
p = 0.18), nor between the two ―recent‖ objects (p > 0.05). When the same
comparison was applied to CA1 and DG/CA3 rats, we found that the rats explored all
the objects at similar rates (p > 0.05).
Discussion
Our results showed that pre-training administration of muscimol in the DG/CA3
impaired spatial discrimination of the objects, although temporal discrimination of the
episodes was not affected. On the other hand, rats treated with muscimol in the CA1
subregion before the first sample trial did not show spatial or temporal discrimination.
These results confirm previous findings showing that dorsal DG/CA3 is crucial for
spatial pattern separation, and dorsal CA1 is involved in temporal pattern separation
26 Dorsal Dentate Gyrus and CA3
The present study confirms and extends previous findings showing that both
dorsal dentate gyrus and CA3 are involved in detecting fine metric changing in the
environment, therefore both structures have a crucial role in rapid encoding of
episodic-like memory. Hunsaker and colleagues (2008b) have demonstrated that rats
with DG lesions could not detect changes in environmental geometry, or fine metric
changes in object location. Other studies from Kesner and colleagues have indicated
a role for the dentate gyrus in fine spatial pattern separation processes (Gilbert et al.,
2001; Hunsaker and Kesner, 2008; Kesner and Hunsaker, 2010). Our results extend
these findings since the dentate gyrus seems to play an important role in
episodic-like memory formation. In addition, these previous studies did not access memory for
‗what-where-when‘. It is also important to note that, as opposed to permanent lesion
approaches, in the present study the hippocampal subregions were temporally
inactivated before training, so that the target regions were inactive during acquisition
and consolidation processes, but not at the test phase. This is in agreement with the
idea that the DG is involved in the rapid formation of episodic memory (Daumas et
al., 2009; Lassalle et al., 2000; Stupien et al., 2003), but not in the retrieval. Lassale
et al. (2000) showed that reversible inactivation of the mossy fibers synapses impairs
learning in the Morris maze, but neither consolidation nor memory recall. Stupien and
colleagues (2003) demonstrated that similar temporary inactivation of the mossy
fibers can impair acquisition and consolidation of object spatial location, but not
memory retrieval. It is interesting that similar results were also found when
hippocampal subregions were inactivated in a contextual fear conditioning task
27 and early consolidation, but not for the recall process. However, further studies are
necessary to test this hypothesis in the episodic-like memory task.
Concerning a methodological issue, it is very unlikely that muscimol injected in
the DG failed to spread to the CA3 subregion. For this reason, we included both
subregions in a single group. In this respect, while the CA3 subregion is treated in
most theoretical models as a homogeneous autoassociative network, recent
empirical studies do not support this notion. Indeed, CA3c sends backprojections
indirectly to DG, and theoretical models have suggested that both regions work
together to generate orthogonal encoding of similar episodes (Myers and Scharfman,
2010). CA3c neurons probably inhibit granule cells in the dentate gyrus indirectly,
possibly contributing to the sparse activity in this region (Myers and Scharfman,
2010). To corroborate this idea, Hunsaker et al. have found that CA3c, but not CA3
a,b, are important to detect fine metric changes in object location (Hunsaker et al.,
2008). Thus, it is possible that in the present study CA3c inactivation was specifically
involved in impaired pattern separation. However, more studies are needed to
evaluate possible differential roles of CA3c and CA3a,b in the episodic-like memory
acquisition.
Concerning a methodological issue, at first glance, it seems that the object
recognition episodic-like memory task employed here did not involve fine pattern
separation, since the old-familiar object was subjected to a large displacement
(around 60 cm). However, it is important to note that the old-familiar displaced object
occupied a place previously occupied by another object in the second sample trial
(fig 1), so it is possible that the rats had to elaborate fine metric relations between
objects in the open field. According to our results, this cognitive process seems to be
28 Dorsal CA1
Rats treated with muscimol in the dorsal CA1 explored all four objects equally
during the test phase, indicating that both the spatial and the temporal discrimination
of the objects were impaired. It has been suggested that dorsal CA1 plays an
important role in temporal pattern separation (Kesner and Hunsaker, 2010; Manns et
al., 2007) . Specifically, this region would be involved in separating different episodes
and creating a temporal context. Hoge & Kesner (2007) have tested the role of CA1
and CA3 on the temporal processing of memories for objects. They found that
CA3a,b lesions did not impair temporal order preference, which means that animals
spent more time exploring old objects than recent objects. Nevertheless, CA1 lesion
rats displayed an opposite pattern, exploring more the recent than the old objects.
These results are at variance with our results, probably due to two different reasons.
First, we applied a longer retention interval (24 h instead of 3 min), evaluating
long-term and not short-long-term episodic-like memory. Second, we interfered with CA1
functioning through a different approach, i.e. the temporary inactivation instead of a
permanent lesion. In the present study, DG/CA3 rats were able to temporally
discriminate the sequence of the objects, probably due to a normal functioning of the
dorsal CA1 subregion in these animals. It is possible that the temporary inactivation
of CA1 impairs the consolidation processes that follow the two sample trials, leading
to a failure of episodic-like memory formation in both training sessions. CA1 is the
major output from the hippocampus and is probably involved in memory
consolidation processes both at the synaptic and the systems levels (Rolls and
Kesner, 2006). Daumas et al. (2005) showed that post-training infusion of CA1 with
29 control animals exhibited a temporal effect, which means an increase in the
percentage of freezing around the second minute of the test session; however CA1
mice did not show this effect. The present results support a key role of CA1 in the
overall consolidation of episodic-like memories, since rats treated with muscimol in
the dorsal CA1 explored the four objects at the same rate.
Conclusions
We found that the DG/CA3 complex is crucial for the formation of episodic-like
memories, being specifically involved in the process of spatial pattern separation that
permits rats detect spatial novelty in the environment. On the other hand, CA1 seems
to be important for temporal pattern separation and the consolidation of the different
episodes. To our knowledge, this is the first study to evaluate the role of these
hippocampal subregions in the acquisition of an episodic-like memory task, i.e.,
addressing what-where-when of a certain episode. These results corroborate
computational models suggesting an important role of DG/CA3 in spatial pattern
separation and of CA1 in the acquisition/consolidation process of the different
episodes. Further studies are needed to unravel how exactly the dentate gyrus and
the CA3 field work together to permit episodic-like memory formation, and to
determine whether these structures are also important for retrieval during the test
30 Acknowledgments
The authors would like to thank Ana Paula Lima and Aline Dierschnabel for technical
assistance and Nelson Lemos for image plotting. We would also like to thank Alicia
Cabral for helping in the behavioral sessions.
Figures:
Fig 1: Schematic drawing of the experimental design.
34 References
Barbosa FF, Pontes IMD, Ribeiro AM, Silva RH. 2010. Extending possible
applications of an episodic-like memory task in rats. Behavioural Brain
Research 215(2):326-331.
Clayton NS, Bussey TJ, Dickinson A. 2003. Can animals recall the past and plan for
the future? Nature Reviews Neuroscience 4(8):685-691.
Daumas S, Ceccom J, Halley H, Frances B, Lassalle JM. 2009. Activation of
metabotropic glutamate receptor type 2/3 supports the involvement of the
hippocampal mossy fiber pathway on contextual fear memory consolidation.
Learning & Memory 16(8):504-507.
Dere E, Huston JP, Silva M. 2005a. Integrated memory for objects, places, and
temporal order: Evidence for episodic-like memory in mice. Neurobiology of
Learning and Memory 84(3):214-221.
Dere E, Huston JP, Silva MAS. 2005b. Episodic-like memory in mice: Simultaneous
assessment of object, place and temporal order memory. Brain Research
Protocols 16(1-3):10-19.
Dere E, Kart-Teke E, Huston JP, Silva MAD. 2006. The case for episodic memory in
animals. Neuroscience and Biobehavioral Reviews 30(8):1206-1224.
DeVito LM, Eichenbaum H. 2010. Distinct contributions of the hippocampus and
medial prefrontal cortex to the "what-where-when" components of episodic-like
memory in mice. Behavioural Brain Research 215(2):318-325.
Gilbert PE, Kesner RP, Lee I. 2001. Dissociating hippocampal subregions: A double
35 Good MA, Barnes P, Stual V, McGregor A, Honey RC. 2007. Context- but not
familiarity-dependent forms of object recognition are impaired following
excitotoxic hippocampal lesions in rats. Behavioral Neuroscience
121(1):218-223.
Hoge J, Kesner RP. 2007. Role of CA3 and CA1 subregions of the dorsal
hippocampus on temporal processing of objects. Neurobiology of Learning
and Memory 88(2):225-231.
Hunsaker MR, Kesner RP. 2008. Evaluating the differential roles of the dorsal
dentate gyrus, dorsal CA3, and dorsal CA1 during a temporal ordering for
spatial locations task. Hippocampus 18(9):955-964.
Hunsaker MR, Rosenberg JS, Kesner RP. 2008. The Role of the Dentate Gyrus,
CA3a,b, and CA3c for Detecting Spatial and Environmental Novelty.
Hippocampus 18(10):1064-1073.
Jung MW, McNaughton BL. 1993. Spatial selectivity of unit-activity in the
hippocampal granular layer. Hippocampus 3(2):165-182.
Kesner RP, Hunsaker MR. 2010. The temporal attributes of episodic memory.
Behavioural Brain Research 215(2):299-309.
Lassalle JM, Bataille T, Halley H. 2000. Reversible inactivation of the hippocampal
mossy fiber synapses in mice impairs spatial learning, but neither
consolidation nor memory retrieval, in the Morris navigation task. Neurobiology
of Learning and Memory 73(3):243-257.
Li JS, Chao YS. 2008. Electrolytic lesions of dorsal CA3 impair episodic-like memory
in rats. Neurobiology of Learning and Memory 89(2):192-198.
Manns JR, Eichenbaum H. 2006. Evolution of declarative memory. Hippocampus
36 Manns JR, Howard MW, Eichenbaum H. 2007. Gradual changes in hippocampal
activity support remembering the order of events. Neuron 56(3):530-540.
McClelland JL, McNaughton BL, Oreilly RC. 1995. Why there are complementary
learning-systems in the hippocampus and neocortex - insights from the
successes and failures of connectionist models of learning and memory.
Psychological Review 102(3):419-457.
Myers CE, Scharfman HE. 2010. Pattern separation in the dentate gyrus: A role for
the CA3 backprojection. Hippocampus:n/a-n/a.
Norman KA, O'Reilly RC. 2003. Modeling hippocampal and neocortical contributions
to recognition memory: A complementary-learning-systems approach.
Psychological Review 110(4):611-646.
O'Reilly RC, Norman KA. 2002. Hippocampal and neocortical contributions to
memory: advances in the complementary learning systems framework. Trends
in Cognitive Sciences 6(12):505-510.
Paxinos G, Watson C. 2009. The Rat Brain in stereotaxic coordinates: Elsevier. 400
p.
Rolls ET. 2010. A computational theory of episodic memory formation in the
hippocampus. Behavioural Brain Research 215(2):180-196.
Rolls ET, Kesner RP. 2006. A computational theory of hippocampal function, and
empirical tests of the theory. Progress in Neurobiology 79(1):1-48.
Scoville WB, Milner B. 1957. Loss of recent memory after bilateral hippocampal
lesions. Journal of Neurology Neurosurgery and Psychiatry 20(1):11-21.
Stupien G, Florian C, Roullet P. 2003. Involvement of the hippocampal CA3-region in
acquisition and in memory consolidation of spatial but not in object information
37 Tulving E. 2001. Episodic memory and common sense: how far apart? Philosophical
Transactions of the Royal Society of London Series B-Biological Sciences
356(1413):1505-1515.
Tulving E. 2002. Episodic memory: From mind to brain. Annual Review of
38 Conclusão Geral
Há muito é estudado o papel do hipocampo na formação da memória
declarativa, mas apenas mais recentemente é que teve início o desenvolvimento de
tarefas comportamentais para estudar memória do tipo episódica em animais. A
memória episódica é o primeiro sistema de memória afetado na Doença de
Alzheimer, logo é fundamental termos uma melhor compreensão dos mecanismos
neurais básicos desse tipo de memória. Apenas recentemente foi desenvolvida uma
tarefa de reconhecimento de objetos que acessa memória episódica em roedores, e
aqui demonstramos que esse tipo de tarefa também funciona com um intervalo de
retenção de 24 h. Além do mais, quando injetamos uma droga amnésica clássica,
escopolamina, os animais tiveram prejuízo na discriminação espaço-temporal dos
objetos. A validação desse protocolo com este intervalo entre treino e teste permite
um uso mais amplo da tarefa para estudar os mecanismos neurais subjacentes a
memória similar a episódica, como, por exemplo, manipulações farmacológicas nas
diferentes fases de formação da memória. Isso nos permitiu estudar o papel de
diferentes sub-regiões hipocampais na aquisição desse tipo de memória. Diversos
estudos teóricos têm sugerido que o giro denteado (GD) e CA3 estão envolvidos na
aquisição rápida da memória episódica, enquanto a sub-região CA1 estaria
envolvida na separação temporal de diferentes episódios. A inativação pré-treino do
GD/CA3 prejudicou a discriminação espacial dos objetos, enquanto que a inativação
de CA1 levou a exploração igual dos objetos. Estes resultados corroboram os
modelos teóricos, indicando um papel importante de GD/CA3 na aquisição rápida de
memórias episódicas, assim como na separação de padrões espaciais. Já CA1 deve
39 são necessários no futuro para separar o papel especificamente do GD e de CA3,
assim como separar a função das regiões CA3 a,b e CA3 c.
Referências
BABB, S. J.; CRYSTAL, J. D. Episodic-like memory in the rat. Current Biology [S.I.], v. 16, n. 13, p. 1317-1321, Jul 2006.
CLAYTON, N. S. et al. Can animals recall the past and plan for the future? Nature Reviews Neuroscience [S.I.], v. 4, n. 8, p. 685-691, Aug 2003.
CRYSTAL, J. D. Elements of episodic-like memory in animal models. Behavioural Processes [S.I.], v. 80, n. 3, p. 269-277, Mar 2009.
DERE, E. et al. Integrated memory for objects, places, and temporal order: Evidence for episodic-like memory in mice. Neurobiology of Learning and Memory [S.I.], v. 84, n. 3, p. 214-221, Nov 2005a.
DERE, E. et al. Episodic-like memory in mice: Simultaneous assessment of object, place and temporal order memory. Brain Research Protocols [S.I.], v. 16, n. 1-3, p. 10-19, Dec 2005b.
DERE, E. et al. The case for episodic memory in animals. Neuroscience and Biobehavioral Reviews [S.I.], v. 30, n. 8, p. 1206-1224, 2006.
EICHENBAUM, H. A cortical-hippocampal system for declarative memory. Nature Reviews Neuroscience [S.I.], v. 1, n. 1, p. 41-50, Oct 2000.
EICHENBAUM, H.. Hippocampus: Cognitive processes and neural representations that underlie declarative memory. Neuron [S.I.], v. 44, n. 1, p. 109-120, Sep 2004.
EICHENBAUM, H. et al. Episodic recollection in animals: "If it walks like a duck and quacks like a duck...". Learning and Motivation [S.I.], v. 36, n. 2, p. 190-207, May 2005.
ERGORUL, C.; EICHENBAUM, H. The Hippocampus and Memory for "What," "Where,"" and "When". Learning & Memory [S.I.], v. 11, n. 4, p. 397-405, Jul-Aug 2004.
FERBINTEANU, J. et al. Episodic memory - From brain to mind. Hippocampus [S.I.], v. 16, n. 9, p. 691-703, 2006.
40 GUZOWSKI, J. F. et al. Ensemble dynamics of hippocampal regions CA3 and CA1. Neuron [S.I.], v. 44, n. 4, p. 581-584, Nov 2004.
KART-TEKE, E. et al. Reinstatement of episodic-like memory in rats by neurokinin-1 receptor antagonism. Neurobiology of Learning and Memory [S.I.], v. 87, n. 3, p. 324-331, Mar 2007.
KART-TEKE, E. et al. Wistar rats show episodic-like memory for unique experiences. Neurobiology of Learning and Memory [S.I.], v. 85, n. 2, p. 173-182, Mar 2006.
LEUTGEB, J. K. et al. Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science [S.I.], v. 315, n. 5814, p. 961-966, Feb 2007.
MCCLELLAND, J. L. et al. WHY THERE ARE COMPLEMENTARY LEARNING-SYSTEMS IN THE HIPPOCAMPUS AND NEOCORTEX - INSIGHTS FROM THE SUCCESSES AND FAILURES OF CONNECTIONIST MODELS OF LEARNING AND MEMORY. Psychological Review [S.I.], v. 102, n. 3, p. 419-457, Jul 1995.
MOSER, E. I. et al. Place cells, grid cells, and the brain's spatial representation system. Annual Review of Neuroscience [S.I.], v. 31, p. 69-89, 2008.
NORMAN, K. A.; O'REILLY, R. C. Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach. Psychological Review [S.I.], v. 110, n. 4, p. 611-646, Oct 2003.
O'REILLY, R. C.; NORMAN, K. A. Hippocampal and neocortical contributions to memory: advances in the complementary learning systems framework. Trends in Cognitive Sciences [S.I.], v. 6, n. 12, p. 505-510, Dec 2002.
O'REILLY, R. C.; RUDY, J. W. Conjunctive representations in learning and memory: Principles of cortical and hippocampal function. Psychological Review [S.I.], v. 108, n. 2, p. 311-345, Apr 2001.
OKEEFE, J.; DOSTROVS.J. HIPPOCAMPUS AS A SPATIAL MAP - PRELIMINARY EVIDENCE FROM UNIT ACTIVITY IN FREELY-MOVING RAT. Brain Research [S.I.], v. 34, n. 1, p. 171-&, 1971.
OKEEFE, J.; RECCE, M. L. PHASE RELATIONSHIP BETWEEN HIPPOCAMPAL PLACE UNITS AND THE EEG THETA-RHYTHM. Hippocampus [S.I.], v. 3, n. 3, p. 317-330, Jul 1993.
SCOVILLE, W. B.; MILNER, B. LOSS OF RECENT MEMORY AFTER BILATERAL HIPPOCAMPAL LESIONS. Journal of Neurology Neurosurgery and Psychiatry [S.I.], v. 20, n. 1, p. 11-21, 1957.
41 SQUIRE, L. R.; ZOLA, S. M. Structure and function of declarative and nondeclarative memory systems. Proceedings of the National Academy of Sciences of the United States of America [S.I.], v. 93, n. 24, p. 13515-13522, Nov 1996.
TULVING, E. Episodic memory and common sense: how far apart? Philosophical Transactions of the Royal Society of London Series B-Biological Sciences [S.I.], v. 356, n. 1413, p. 1505-1515, Sep 2001.
TULVING, E. Episodic memory: From mind to brain. Annual Review of Psychology [S.I.], v. 53, p. 1-25, 2002.