O EFEITO “INIMIGO ÍNTIMO” É
APLICÁVEL A Dinoponera quadriceps
(HYMENOPTERA, FORMICIDAE)?
.
O EFEITO “INIMIGO ÍNTIMO” É APLICÁVEL A Dinoponera quadriceps (HYMENOPTERA, FORMICIDAE)?
Dissertação apresentada à Universidade Federal do Rio Grande do Norte para obtenção do título de mestre em Psicobiologia.
Orientador: Professor Arrilton Araújo
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Moreno, Isabelli de Carvalho.
O efeito “inimigo íntimo” é aplicável a Dinoponera quadríceps (Hymenoptera, Formicidae)? / Isabelli de Carvalho Moreno. – Natal, RN, 2010.
34 f. : Il.
Orientador: Prof. Arrilton Araújo
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Formicidae – Dissertação 2. Dinoponera quadriceps –
Dissertação. 3. Efeito inimigo íntimo – Dissertação. I. Araújo, Arrilton. II. Universidade Federal do Rio Grande do Norte. III. Título.
Autora: Isabelli de Carvalho Moreno
Data da defesa: 28 de maio de 2010
Banca Examinadora:
______________________________________ Profa. Ana Maria Matoso Bailez-Viana
Universidade Estadual do Norte Fluminense, RJ
______________________________________ Profa. Maria de Fátima Arruda
Universidade Federal do Rio Grande do Norte, RN
______________________________________ Prof. Márcio Zikán Cardoso
Universidade Federal do Rio Grande do Norte, RN
______________________________________ Prof. Arrilton Araújo
Agradeço a Deus pela realização deste trabalho e por todas as bênçãos alcançadas. Arrilton ter me aceito como aluna, por sua confiança de que eu iria realizar este projeto, pela paciência e orientação. Por está sempre presente para tirar dúvidas e sempre de bom humor!
A todos aqueles que acordaram cedo no fim de semana para ajudar na coleta das colônias: Anderson, Marcelo, Dina, Jeniffer, Priscila, Audra, Adriano, Cleine, aos
funcionários anônimos da UFRN que deram aquela “forcinha” e emprestaram seus
materiais. Sem vocês não haveria as colônias no laboratório!
Ao diretor presidente da EMPARN Dr. Henrique Eufrásio de Santana Júnior que autorizou a coleta de colônias nas dependências da Base Física Rommel Mesquita de Faria – EMPARN/JIQUI.
Mário Sérgio, funcionário da EMPARN/JIQUI que nos recebeu muito bem e nos deu livre acesso à reserva.
Marcos do Laboratório de Entomologia que nos apresentou ao Mário Sérgio, nos conduziu à EMPARN/JIQUI e mostrou as trilhas da reserva.
Agradeço a Ingrid, minha “câmera girl” que me ajudou nas coletas de dados, marcação das operárias e manutenção das colônias no laboratório.
As amigas Audra, Dina, Priscila e Kelly sempre presentes me ajudando nos momentos
de estresse, “coorientando” e pela companhia, festas, almoços, serviços de baby siter etc. Meninas, obrigada por tudo!!! Vocês foram muito importantes nessa trajetória.
Marcelo pela sua “consultoria” no Excel, estatística, formatação, caronas, enfim, por
está sempre disponível para ajudar.
Dina que me ajudou desde o piloto, me apresentando aos comportamentos agonísticos exibidos pelas operárias, me ensinou a marcação, ajudou na estatística, em tudo! Dina muito obrigada por tudo que você me ensinou!
Agradeço a minha família que me apoiou sempre, minha mãe que se dispôs a ficar com Linda para que fosse possível a realização deste trabalho. Meu pai e irmãos que ajudaram sempre que precisei.
Linda que apesar da pouca idade compreende a ausência da mamãe.
Agradeço à Fapern, CNPq e CAPES pelo apoio financeiro para compra de equipamentos e pela bolsa de pesquisa.
Ao Instituto Chico Mendes de Biodiversidade- ICMbio que autorizou a retiradas das colônias e sua manutenção em laboratório para esta pesquisa.
Aos professores da Pós Graduação em Psicobiologia e também aos professores da graduação que incentivaram meu interesse para a pesquisa.
A Dr. Deborah M. Gordon pela atenção e envio de seu artigo.
O acesso a fonte de alimentos, território ou parceiros reprodutivos pode gerar conflitos de interesses entre indivíduos em várias espécies de animais. Entretanto, vários animais respondem menos agressivamente à entrada de seus vizinhos do que a entrada de não-vizinhos em seus territórios, como forma de minimizar os custos de uma agressão contínua. Essa diferença na resposta agressiva é conhecida por Efeito Inimigo Íntimo, descrito em várias espécies de vertebrados e invertebrados. Para investigar se este fenômeno ocorre em Dinoponera quadriceps (Hymenoptera, Formicidae), três colônias, sendo duas vizinhas e uma distante, foram coletadas em seu ambiente natural e transferidas para o laboratório onde foram conduzidos confrontos experimentais intra e entre colônias, envolvendo um par de operárias. Comparamos a frequência comportamental exibida por cada operária envolvida nos confrontos, assim como a intensidade e a duração dos confrontos entre colônias vizinhas e distantes com referência do local de coleta. Nossos resultados revelam que o Efeito Inimigo Íntimo não é aplicável à D. quadriceps, pois resposta agressiva é mais intensa e duradoura entre operárias vizinhas do que entre distantes, provavelmente devido à competição intra-específica, fatores ecológicos e características da espécie.
Food access, territory or reproductive partner can generate conflicts between individuals in many species with occurrence of aggressive behaviors. However some species respond less aggressively to intrusion by neighbors than non-neighbors in its territory to minimize the costs of continuous fight. This difference in aggression is called Dear Enemy Effect described in various vertebrates and invertebrates. To investigate if this phenomenon occurs in Dinoponera quadriceps (Hymenopetra, Formicidae) three colonies, two neighbors and one non-neighbor, were captured in its natural environment then transfered to the laboratory where we did experimental confrontation intra and inter colonies involving one pair of workers. We compared the behavioral frequency exhibited by each worker, the intensity and duration of the confrontation between a neighbor and a non-neighbor referring the place where they were collected. Our results revealed that Dear Enemy Effect does not apply to D. quadriceps due aggressive response is more intense and longest toward neighbor than non-neighbor, probably due intra specific competition, ecological factors and characteristic of the species.
Introdução 1
Objetivo 7
Objetivos Específicos 7
Hipóteses e Predições 7
Metodologia 8
Resultados 16
Discussão 22
Introdução
O acesso à fonte de alimentos, território ou parceiros reprodutivos pode gerar conflitos entre indivíduos em várias espécies, envolvendo comportamentos agressivos (Huntingford & Turner 1987). Essas situações ocorrem com mais frequência do que esperado diante dos custos que esses comportamentos provocam. Os animais podem minimizar estes custos iniciando confrontos com comportamentos estereotipados feitos a distância tais como vocalizações, exibição dos aparatos físicos, ou estabelecendo hierarquia de dominância (Matthews & Matthews 2010). Posteriormente os confrontos podem evolver contato físico com maior gasto energético sem causar danos físicos, como lutas de caudas em peixes (Huntingford & Chellappa 2006). Finalmente, pode atingir um estágio envolvendo mordidas ou ataques, injúrias e até mesmo a morte de um dos envolvidos (Huntingford & Turner 1987). O valor do recurso e a capacidade física do individuo é que vão determinar em que estágio o conflito será resolvido (Krebs & Davies 1996). Quando um encontro agonístico é resolvido sem haver luta, o confronto é
denominado “ritualizado” (Huntingford & Turner 1987).
pode estar à procura de um novo território (Temeles 1994). Outra hipótese baseia-se na redução dos custos energéticos de uma interação agressiva e na prevenção de lutas com vizinhos já que são frequentes os encontros durante o forrageio (Ydenberg et al 1988). A luta entre estranhos pode ser funcional para adquirir informações sobre condição e motivação do oponente ou simplesmente resultar de erros frequentes de avaliação estimando estas informações (Ydenberg et al. 1988, 1989; Getty 1989). Outra explicação para a variação da agressividade entre vizinhos é o fenômeno da habituação, descrito como rápida forma de aprendizagem que resulta na diminuição da resposta a um estímulo percebido repetidas vezes, como um odor persistente (Langen at al. 2000).
Todavia, as relações entre vizinhos dependem dos fatores ecológicos e ambientais. Se houver competição por recurso, o custo da interação com um vizinho é elevado e eles são tratados como ameaça, elevando a resposta agressiva. Neste caso, o fenômeno inimigo íntimo tende a não ocorrer (Gordon 1989).
As formigas, de modo geral, controlam e defendem seus territórios que fornecem recursos para alimentar o grande número de larvas, machos e fêmeas reprodutores (Newey et al 2010). Na maioria das espécies, encontros entre indivíduos de outras colônias podem gerar lutas ritualizadas. Em função do grau de competição, estes podem se transformar em lutas, com contatos físicos, resultando em mortes de vários indivíduos (Matthews & Matthews 2010).
de uma mesma colônia (Ratnieks et al. 2006). Isto ocorre provavelmente porque os insetos na realidade não são clones, mas famílias de indivíduos com elevado grau de parentesco apresentando diferentes interesses na maioria dos casos (Ratnieks et al. 2006). Fatores como número de rainhas e número de acasalamentos podem intensificar os conflitos entre os indivíduos devido a diluição do parentesco (Bourke & Franks 1995).
Para a manutenção da colônia, é necessário que haja mecanismos de reconhecimento que garanta a coesão social e previna a entrada de co-específicos
(Guerrieri & d‟Etorre 2007, Björkman-Chiswell et al. 2008). Nos insetos sociais o reconhecimento de indivíduos da mesma colônia se dá através dos hidrocarbonetos cuticulares (Gamboa et al. 1996, Lahav et al. 1999, Wagner et al. 2000, Lenoir et al. 2001) que podem ser geneticamente determinados (Beye et al. 1998; Suarez et al. 2002, Guerrieri & d‟Ettorre 2007, Thurin & Aron 2007) ou adquiridos através do ambiente, como odor da colônia (Carlin & Hölldobler 1983) e influência da dieta (Liang & Silverman 2000, Buczkowski & Silverman 2006, Sorvari et al. 2008) ou pela combinação dos dois componentes (Jutsum et al. 1979, Crosland 1989; Pirk et al. 2001, Suarez et al. 2002). Estes sinais são compartilhados pelos indivíduos da colônia através das interações sociais como trofilaxia, “grooming” e contato físico (Soroker et al. 1998; Lahav et al. 1999).
Herbers 2000, Boulay & Lenoir 2001, Suarez et al. 2002, Roulston et al. 2003, Buczkowski & Silverman 2005, Menzel et al. 2009).
Em relação às formigas o fenômeno inimigo íntimo varia. A redução da agressividade aos vizinhos é descrita em Acromyrmex octospinosus (Formicidae: Formicinae) (Jutsum et al. 1979), Leptothorax nylanderi (Formicidae: Myrmicinae) (Heinze et al. 1996), Pheidole tucsonica e Pheidole gilvescens (Formicidae: Myrmicinae) (Langen et al. 2000), Camponotus chilensis (Formicidae: Formicinae) (Velásquez et al. 2006) Formica xerofila e Formica intergroides (Formicidae: Formicinae) (Tanner & Adler 2009), Atta sexdens rubropilosa (Formicidae: Myrmicinae) (Araújo et al. 2009) e Acromyrmex lobicornis (Formicidae: Myrmicinae) (Dimarco et al. 2010).
Por outro lado, a elevação da resposta agressiva após encontros repetidos também pode ocorrer em Pogonomyrmex barbatus (Formicidae: Myrmicinae) (Gordon 1989) Cataglyphis fortis (Formicidae Formicinae) (Knaden & Wehner 2003), Pristomyrmex punctatus (anteriomente P.pungens) (Formicidae: Myrmicine) (Sanada-Morimura et al. 2003) e em Oecoplhilla smaragdina (Newey et al. 2010). Em Linepitherna humile (Formicidae: Dolichoderinae) variações nos níveis de agressividade ocorrem em função da experiência individual, defesa territorial e contextos sociais e/ou ecológicos (Suarez et al. 2002; Thomas et al. 2004, Buczkowski & Silverman 2005, 2006). Em Camponotus cruentatus não há diferença na agressividade em função das distâncias entre os ninhos (Boulay et al. 2007).
As formigas da subfamília Ponerinae se destacam por serem predadoras e por possuírem um ferrão funcional com o qual paralisam suas presas e utilizam para a defesa. As representantes do gênero Dinoponera são endêmicas da América do Sul e caracterizam-se pela ausência de diferenciação morfológica entre as operárias e a operária reprodutora. São consideradas primitivas por sua morfologia e comportamentos (Caetano et al. 2002).
Na espécie Dinoponera quadriceps (Formicidae: Ponerinae), endêmica do Nordeste Brasileiro, não há a presença de uma rainha, mas de uma fêmea reprodutora dominante chamada “gamergate”. Todas as operárias tem capacidade reprodutiva, ou seja, são totipotentes. Os fatores que definem o status reprodutivo são a hierarquia linear, sinais químicos e comportamentos agonísticos da gamergate (Monnin & Peeters 1998). As colônias possuem, em média, 89 indivíduos, podendo variar de 30 a 240 operárias (Monnin & Peeters 1997, 1999). As operárias desta espécie medem aproximadamente 3 centímetros de comprimento (figura 1).
As fêmeas não apresentam vôo nupcial, mas os machos são alados e voam para outras colônias copulando com fêmeas na entrada ou no interior do ninho (Monnin & Peeters 1998). Estas características certamente influenciam o modo de fundação de
novas colônias, que podemos deduzir logicamente que por uma nova “gamergate”
(operária fecundada) que deixa a colônia natal acompanhada de uma pequena corte de operárias para fundar a sua própria colônia ou ocupar um ninho vazio. É pouco provável
que uma jovem “gamergate” possa sozinha realizar todas as tarefas de uma nova
fundada pela “gamergate” compartilha relações genéticas com a colônia mãe, de onde ela saiu.
Figura 1. Dinoponera quadriceps
As operárias apresentam forrageio solitário, sem recrutamento, e transportam principalmente restos de artrópodes para o ninho e alguns itens de origem vegetal como frutos e sementes (Araújo & Rodrigues 2006). As operárias apresentam fidelidade às áreas de caça, retornando aos mesmos locais ao longo do dia e podem se distanciar mais de 50 m do ninho durante o forrageio (Azevedo 2009). Nestes deslocamentos é comum o encontro entre operárias de colônias distintas. Nestes encontros podem ser vistos desde toques de antenas a confrontos que variam em duração e intensidade. No entanto, operárias apresentam a tendência de evitar a área de uma colônia vizinha, forrageando preferencialmente na direção oposta (Medeiros 2009). Todavia não há estudos que permitam apontar as causas da existência de confrontos em D. quadriceps, nem tampouco a variabilidade na intensidade dos mesmos.
Muitos autores tem investigado sobre as características ecológicas de Dinoponera quadriceps, sobretudo o forrageio (Araújo & Rodrigues 2006, Azevedo 2009) e características de sua estrutura social e reprodutiva assim como conflitos internos (Monnin & Peeter 1997, 1998, 1999, Monnin & Ratnieks 1999, Monnin et al. 2003). Dentro deste contexto, este trabalho procurou investigar os mecanismos relacionados
Objetivo
Este trabalho tem como objetivo verificar a ocorrência do efeito do “Inimigo Íntimo” em operárias de Dinoponera quadriceps em laboratório.
Objetivos específicos
Comparar a frequência dos comportamentos agressivos exibidos durante os confrontos entre operárias de mesma colônia, colônias vizinhas e distantes.
Investigar se há diferença comportamental entre as operárias residentes e intrusas. Analisar se existe relação entre a duração e a intensidade do confronto.
Hipótese e Predições
Hipótese: a ocorrência de comportamentos agressivos está relacionada com o não reconhecimento entre as operárias.
o Operárias de mesma colônia se reconhecem e não demonstram
comportamentos agressivos durante os encontros.
o Operárias discriminam colônias vizinhas e distantes, demonstrando baixa
Metodologia
Local de Coleta de Dados
Para este trabalho, foram coletadas três colônias de Dinoponera quadriceps em ambiente natural, com a autorização do ICMBio. Duas colônias foram coletadas dentro do Campus da Universidade Federal do Rio Grande do Norte (Colônia A - 5° 50‟ 13” S e 35° 12‟ O; Colônia B - 5° 50‟ 14”S e 35° 12‟ O), localizado no município de Natal-RN, contendo respectivamente, 113 e 83 operárias, 23 e 9 pupas, larvas, ovos e um macho em cada colônia. As entradas dos ninhos distavam 46 m. A terceira colônia foi coletada no município de Parnamirim – RN, dentro da reserva da Emparn, distante 7,6 Km da primeira área de coleta (55 47.5 SS e 35 11 18.3 W). No ninho havia 48 operárias, 9 larvas e 9 pupas.
A distância entre as duas áreas garante a ausência de encontros prévios entre as operárias. No ambiente do Campus Universitário, as colônias coletadas são consideradas vizinhas, pois em observações prévias verificamos sobreposição de áreas, havendo encontros entre as operárias durante o forrageio.
Cada operária foi marcada com etiqueta numerada colada no tórax (Corbara et al. 1986). As etiquetas apresentavam cores diferentes para cada uma das colônias a fim de facilitar a identificação (figura 3).
Figura 2. Estrutura de disposição dos ninhos e arenas de forrageamento em laboratório.
Figura 3. Operárias de colônias diferentes. Cada cor da etiqueta representa uma colônia.
A temperatura e a umidade do laboratório foram mantidas, respectivamente a 25 ± 2 º C e 65 ± 10% e o regime de luz 12:12 claro/escuro. A alimentação foi mantida de modo semelhante ao campo, sendo oferecidos itens de origem animal e vegetal
Foto: Dina Líllia
conforme descrito por Araújo & Rodrigues (2006). As três colônias receberam a mesma dieta e proporção de alimento a qual era disposta nas arenas de modo aleatório tanto espacial como temporalmente durante a fase clara pelo menos uma vez ao dia, e água disposta ad libitum.
Procedimento Experimental
1. A primeira etapa consistiu de encontros entre operárias de mesmo ninho (experimento controle).
Para cada colônia foram feitos 30 ensaios experimentais com duração de 15 minutos cada. Os encontros foram induzidos nas arenas de forrageamento. Duas operárias, presentes na arena, eram colocadas em uma mini arena de 15 cm de diâmetro, no centro da maior. A estrutura de plástico aumentava a probabilidade de uma operária perceber a presença da outra (figura 4). Após os primeiros contatos, a estrutura era removida.
Figura 4. Área delimitada para início dos encontros.
Após um minuto iniciamos o registro da frequência dos comportamentos
exibidos pelas operárias alvos. O registro da frequência comportamental foi realizado simultaneamente por dois observadores com grau de concordância superior a 95%, através da técnica focal instantâneo com registro a cada um minuto a partir do encontro até o término do experimento (Martin & Bateson 2007). Durante os experimentos, o acesso de outras operárias à arena foi impedido.
Os comportamentos registrados foram:
Ignorar – contato entre indivíduos em que nenhum demonstra interesse para o outro (Thomas et al. 2004);
Toque - quando uma operária toca as antenas ou partes do corpo da outra operária (Suarez et al. 1999);
Fuga – após toque, ou mesmo após uma agressão, a(s) operária(s) desloca(m)-se em direção oposta (Suarez et al. 1999);
Tentativa de escape – operária imobilizada abre sua mandíbula, movimenta a cabeça e o corpo em direção oposta à operária agressora para escapar da agressão;
Abertura de mandíbula: operária abre a mandíbula em direção à oponente ameaçando-a (Mercier et al. 1997) (figura 5A);
Tentativa de apreensão: operária abre e fecha as mandíbulas tentando prender a mandíbula, antena ou pernas de sua oponente;
Apreensão: operária fecha sua mandíbula imobilizando a mandíbula, antena ou perna da oponente (Mercier et al. 1997) (figura 5B);
Boxe antenal: a operária agressora bate repetidamente na cabeça da oponente com suas antenas (Monnin e Peeters 1999) (figura 5C);
Exibição do abdômen: a operária agressora encurva seu abdômen entre as pernas em direção a oponente (Monnin e Peeters 1999) (figura 5D);
Luta – inclui uma agressão recíproca e intensa, em que operárias esfregam o abdômen do corpo da outra, prendem suas mandíbulas mutuamente na cabeça, tórax, abdômen ou pernas e eliminação ácido fórmico perceptível através do cheiro no ambiente (Mercier et al. 1997). Neste estágio pode haver prejuízo para uma ou ambas, pois podem perder segmentos do corpo e até mesmo haver morte de uma das duas (figura 5E);
Perseguição: após a fuga, operária agressora desloca-se rapidamente na mesma direção da operária que fugiu.
Para avaliar a intensidade da agressão entre as operárias, os comportamentos exibidos receberam uma escala de pontuação de 0 a 6 a partir de Suarez et al. (1999). Sendo,
0: comportamento de ignorar; 1: toque;
2: comportamento de fuga;
3: operária exibe um único comportamento agressivo no momento do registro; 4: operária exibe simultaneamente dois comportamentos agressivos no momento do registro, por exemplo, apreensão e boxe antenal;
5: operária exibe simultaneamente três comportamentos agressivos no momento do registro, por exemplo, apreensão, boxe antenal e corpo elevado;
Figura 5. Comportamentos agonísticos. A: mandíbula aberta, B: apreensão, C: corpo elevado e boxe antenal, D: exibição do abdômen, E: luta.
Foto: Isabelli Moreno Foto: Isabelli Moreno
Foto: Isabelli Moreno
Foto: Dina Líllia Foto: Monnin
A
E D
B
Esta escala foi utilizada para verificar se havia diferença de intensidade agressiva entre operárias de mesmo ninho, vizinhas e distantes. Foi utilizada nas análises a pontuação máxima de agressão exibida durante os experimentos.
Foram utilizadas somente operárias que continham todos os apêndices e aquelas que estavam na arena de forrageamento. Registramos o número de operárias que morreram ou sofreram injúria em consequência da agressão.
2. Na segunda etapa realizamos encontros entre operárias de colônias diferentes, de acordo com as distâncias originais. Nesta etapa não interferimos na duração dos experimentos. As observações foram feitas até que as operárias alvos finalizassem o encontro.
2.1. Encontros experimentais entre operárias das colônias vizinhas. Foram realizados 60 encontros, sendo 30 na condição de residentes e 30 como intrusas. Os procedimentos adotados foram os mesmos descritos acima.
2.2. Encontros experimentais entre operárias provenientes de colônias distantes. Foram realizados 120 encontros entre as três colônias, adotando os mesmos procedimentos descritos acima na fase anterior.
Análise Estatística
entre operária de diferentes colônias tiveram duração variada, dividimos a frequência de cada comportamento pela duração e multiplicamos por 60. Desta forma obtivemos a frequência por hora de cada comportamento.
Resultados
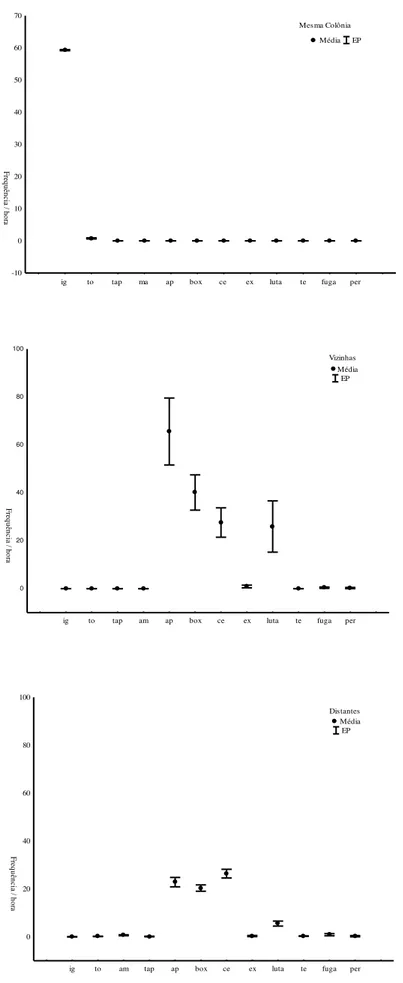
Nos encontros entre operárias de mesma colônia não ocorreram comportamentos agonísticos (H=3643,3 p=0,01). As operárias apresentaram o comportamento de ignorar (U=22,9 p = 0,01) em relação à sua companheira de ninho e eventuais toques de antena (U=5,1 p=0,01) predominando o nível 0 na escala de agressão (figura 6). Nesta situação as operárias permaneciam na arena forrageando, ingerindo água ou realizando limpeza de seu corpo.
Porém, quando uma operária encontrava outra que não pertencia à sua colônia, havia a exibição de comportamentos agonísticos (H=3643,3 p=0,01). Uma ou ambas tocava o corpo da outra utilizando as antenas, em seguida havia exibição agonística mútua, como abertura de mandíbula, elevação do corpo (U=14,4 p=0,01), tentativa de apreensão, geralmente de mandíbula e algumas vezes de pernas ou antenas (U=3,4 p=0,01). Posteriormente ocorria a apreensão de mandíbula (U=11,3 p=0,01), boxe antenal, (U=15,3 p=0,01), exibição do abdômen (U=2,8 p=0,01) e até mesmo luta (U=6,5 p=0,01) (figura 6). A apreensão da mandíbula ocorria do início ao final do confronto, sendo o comportamento mais frequente. Em alguns confrontos ocorria inversão dos papéis: a operária que sofria a apreensão, passava realizá-la. Em alguns confrontos, a operária imobilizada abria a mandíbula e realizava movimentos opostos à agressora para escapar da agressão (tentativa de escape U=2,1 p=0,01) e, obtendo sucesso, fugia (fuga: U=2,6 p=0,01) finalizando o confronto ou dava continuidade, iniciando a sequência comportamental descrita acima. Registramos 20% de luta em 180 confrontos entre colônias. Destes, 18,3% ocorreu fuga em seguida, finalizando. Geralmente precedendo a luta ocorria a exibição do abdômen.
Figura 6. Frequência comportamental (média ± EP) exibida nos confrontos entre operárias de mesma colônia, colônias vizinhas e distantes. Ig = ignorar; to = toque; tap = tentativa de apreensão; ma = mandíbula aberta; ap = apreensão; box = box antenal; ce = corpo elevado; ex = exibição do abdômen; te = tentativa de escape; per = perseguição
ig to tap ma ap box ce ex luta te fuga per -10 0 10 20 30 40 50 60 Fre qu ên cia / h ora
Média EP
ig to tap am ap box ce ex luta te fuga per
0 20 40 60 80 100 Fre qu ên cia / h ora Média EP Vizinhas
Como foram registrados simultaneamente os comportamentos das duas operárias envolvidas nos experimentos, obtivemos dados comportamentais das residentes e intrusas. Mas todos os comportamentos agressivos observados em ambas as situações foram semelhantes, tanto defendendo seu território quanto no território pertencente à oponente, independente se esta era sua vizinha ou não (Mann-Whitney p > 0,05).
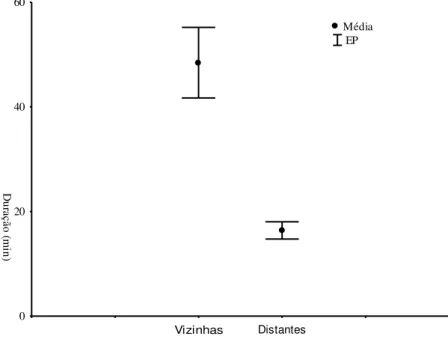
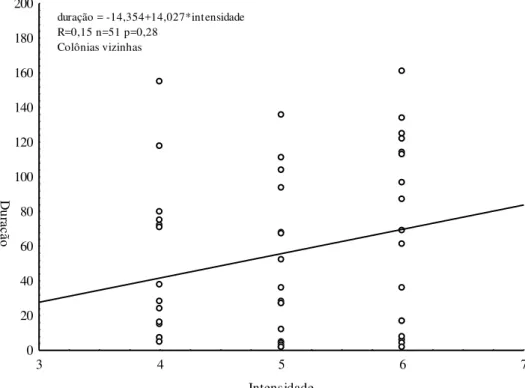
Em relação à duração dos experimentos, encontros entre colônias foi bastante diversificado, variando de 0,18 a 210 minutos. Os encontros entre operárias vizinhas foram mais duradouros comparando-se aos encontros entre colônias distantes do local de coleta (figura 7). Em relação à intensidade obtivemos uma concentração dos níveis 4, 5 e 6 de agressão tanto entre vizinhas quanto entre distantes. No entanto não houve diferença significativa quanto às distâncias das colônias (U=0,85, p=0,38). E não houve correlação entre a duração e intensidade dos confrontos entre colônias vizinhas. Por outro lado, houve correlação positiva entre as colônias distantes (figura 8).
Figura 7. Duração (em minutos) dos encontros experimentais entre colônias vizinhas e distantes com referencia ao local de coleta.
Vizinhas Distantes
0 20 40 60
D
ur
aç
ão
(m
in
)
3 4 5 6 7
Intensidade 0 20 40 60 80 100 120 140 160 180 200 D ur aç ão
duração = -14,354+14,027*intensidade R=0,15 n=51 p=0,28
Colônias vizinhas
3 4 5 6 7
Intensidade 0 20 40 60 80 100 120 140 160 180 200 D ur aç ão
duração = -3,378+4,405*intensidade R=0,24 n = 106 p=0,01
Colônias distantes
Discussão
O Efeito Inimigo Íntimo pode ser influenciado pelas diferentes histórias de vida, condições ecológicas ou modo de fundação de novas colônias (Boulay et al 2007). Habituação, sinais ambientais e genéticos, além do grau de ameaça que o intruso representa, tem sido apontados como causas próximas para o fenômeno (Langen et al. 200, Heinze et al. 1996, Temeles 1994). No entanto, as causas últimas permanecem questionáveis nos insetos sociais (Boulay et al. 2007).
Em nossos experimentos, não houve agressão entre operárias de mesmo ninho, mostrando evidências de sinais de reconhecimento entre elas. Por outro lado, observamos a ocorrência de comportamentos agressivos quando operárias oriundas de outras colônias foram transferidas para o centro da arena. Houve agressividade tanto em operárias de colônias originalmente próximas quanto entre operárias de colônias originalmente distantes. Porém, houve maior frequência de luta entre vizinhas, já que houve 33% de luta nestas, contra 13% em confrontos entre colônias distantes. Essa diferença na resposta agressiva aos vizinhos se dá provavelmente porque em Dinoponera quadriceps vizinhos representam forte ameaça o que pode está relacionado com o modo de forrageio solitário e distribuição dos recursos no ambiente, pois se alimentam de restos de animais mortos ou presas vivas (Araújo e Rodrigues 2006) aumentando seu deslocamento a procura de alimento (Azevedo 2009).
aos vizinhos é descrito em cupins Nasutitermes corniger (Dunn & Messier 1999), nas formigas Pogonomyrmex barbatus (Gordon 1989), Pristomyrmex punctatus (Sanada-Morimura et al. 2003), e em Linepithema humile (Thomas et al. 2004). Já em Diacamma sp (Formicidae: Ponerinae) não há diferença na resposta agressiva entre vizinhos ou indivíduos de colônias distantes, uma vez que as operárias apresentam forrageio solitário. Neste caso, o encontro com outra operária que não pertence à mesma colônia, independente da distância, pode representar uma ameaça (Suwabe et al. 2007).
Em nosso estudo, apesar da maior frequência de luta entre colônias vizinhas não houve diferença estatística de intensidade quando comparamos em função da localização dos ninhos. Mas observamos maior ocorrência de contextos ritualizados entre colônias distantes, fortalecendo a hipótese de que indivíduos estranhos representam pouca ameaça em relação aos vizinhos quando o território é utilizado para alimentação (Temeles 1989). Neste caso, luta entre indivíduos distantes pode resultar em perda de energia, enquanto lutas entre vizinhos podem prover acesso a novos recursos (Dunn & Messier 1999).
Além de uma maior frequência de luta, os encontros experimentais entre vizinhas tiveram maior variabilidade em sua duração e foram mais longos, comparando aos encontros entre colônias distantes os quais foram mais curtos e homogêneos e obtiveram uma correlação positiva com a intensidade.
da competição por alimento (Thomas et al. 2004). Em D. quadriceps a distribuição espacial dos ninhos é influenciada pela competição intra-específica, pois ninhos mais populosos mantém distâncias maiores entre seus vizinhos, mais do que os ninhos menos populosos (Vasconcelos et al. 2004).
O tamanho da colônia também pode influenciar na resposta agressiva, pois colônias grandes possuem maior probabilidade de encontros (Gordon 2010). No nosso estudo, a terceira colônia coletada era menor em relação às demais, o que pode gerar uma postura de submissão e evitar contextos agressivos, pois a perda de operárias implica na produtividade da colônia. Além disso, a presença de uma colônia pequena pode diminuir as chances de sobrevivência de uma colônia grande, pois necessitam de maior quantidade de alimento e consequentemente de uma maior área a ser explorada (Gordon 2010). As duas colônias vizinhas possuíam grande quantidade de operárias, e provavelmente ocorriam encontros periódicos durante o forrageio, aumentando a resposta agressiva em função da instabilidade do ambiente.
As colônias localizadas próximas antes de serem transferidas para o laboratório pertenciam ao mesmo micro-habitat, enquanto a terceira colônia coletada pertencia a uma área com distância superior a 7 km. Heinze et al. (1996) defendem que quando colônias compartilham o mesmo micro-habitat pode haver semelhança de odor produzido pelas colônias e como consequência operárias diminuem a resposta agressiva aos vizinhos. No nosso caso, houve maior frequência de luta entre as vizinhas, sugerindo que, neste caso, a possível semelhança não reduziu a resposta agressiva em virtude da competição entre as colônias.
Em D. quadriceps a similaridade genética não diminui a resposta agressiva entre colônias vizinhas. Medeiros (2009) descreve o processo de fissão em uma colônia de D. quadriceps e relata ocorrência de interação agonística entre a colônia recém formada e a colônia antiga como forma de estabelecimento territorial. Neste caso as interações agonísticas iniciaram após cinco meses da fissão, período provavelmente necessário para que a nova colônia atingisse um estágio mais maduro (Medeiros 2009) e consolidação de um novo odor. Isso demonstra que o odor da colônia é dinâmico e está em permanente mudança (Lenoir et al. 2001). Em Aphaenogaster senilis (Formicidae: Myrmecinae) o isolamento de operárias por 10 dias é capaz de provocar mudanças no perfil dos hidrocarbonetos e desencadear resposta agressiva quando expostas às companheiras de ninho (Lenoir et al. 2001).
Alguns autores também sugerem que a dieta exerce influencia na formação do odor e argumentam que dieta semelhante diminui a resposta agressiva, assim como uma colônia que é separada artificialmente e é fornecida diferente dieta a cada uma, operárias respondem de forma agressiva quando expostas às antigas companheiras de ninho (Jutsum et al. 1979, Liang & Silverman 2000, Sorvari et al. 2008). Em nosso estudo, as três colônias receberam os mesmos itens alimentares (larva de tenébrio, grilos, mariposas e frutas) e mesmo assim demonstraram agressão durante os encontros experimentais, principalmente as colônias que eram vizinhas.
Durante os encontros experimentais e após a finalização, as operárias realizavam auto grooming como forma de manter seu próprio cheiro. Além disso, nos episódios de luta intensa onde havia um maior contato físico entre as operárias rivais e eliminação de ácido fórmico, ao retornarem à arena (no caso de intrusas) havia rejeição pelas companheiras de ninho através de comportamentos agonísticos como apreensão e exibição do abdômen. Isso demonstra que o corpo da operária fica impregnado com o odor da adversária e para manter sua identidade é necessária uma produção contínua dos sinais de reconhecimento, acompanhados de um mecanismo de distribuição na superfície corporal do individuo, assim como a troca entre membros da colônia (Soroker et al. 1998).
Quando avaliamos se operárias residentes respondiam de forma mais agressiva à entrada de intrusos, constatamos uma simetria de comportamentos entre residentes e intrusas. Enquanto uma atacava, a oponente contra-atacava, não assumia submissão. Buczkowski & Silverman (2005) relatam que em Linepithema humile operárias são mais agressivas quando defendem o território. Se as operárias estão longe da sua área de uso, apresentam pouca ou nenhuma agressão a co-específicos. Em Pheidole pallidula intrusas adotam a estratégia burguês, não disputando o território já estabelecido por um proprietário (Cammaerts & Cammaerts 1998). No ambiente natural quando operárias de D. quadriceps estão longe da entrada do ninho, ao encontrar outra elas evitam o contexto agonístico, já quando estão próximas ao ninho, ou em área de sobreposição, ao encontrar uma intrusa, há resposta agressiva com mais frequência, provavelmente como forma de manter a fronteira territorial (Dina Azevedo, comunicação pessoal, 14 de abril de 2010).
foram em encontros entre vizinhas. E dez operárias tiveram perda total ou parcial de pernas ou antenas. Para o total de confrontos realizados, este é um número pequeno de morte, já que houve uma frequência de registro de luta em 20% dos experimentos. No entanto Mercier et al. (1997) afirmam que quando há pelo menos uma morte em 100% dos confrontos significa que há competição entre as duas sociedades. Houve também 23,8% de pequenos episódios de luta que duraram alguns segundos e logo em seguida o confronto era finalizado, onde cada operária seguia direção oposta. Nestas situações as lutas não foram registradas devido à rápida duração e por ocorrer após o último registro. Em 57,9% os confrontos terminaram no estágio de ritualização. Com isso podemos inferir que há um balanço entre o custo da interação agonística e o beneficio (recurso) resultante da competição e as operárias parecem evitar uma agressão mais intensa onde há um maior custo energético. Ao invés disso as operárias realizam ameaça através de exibições de mandíbula e abdômen, elevam o corpo ou realizam boxe antenal e/ou apreensão de partes do corpo da oponente.
Nosso estudo apresenta evidências demonstrando que em Dinoponera quadriceps operárias respondem de forma mais agressiva às suas vizinhas do que às operárias de colônias distantes, prevalecendo o efeito “vizinho indesejável” (nasty neighbor) (Newey et al. 2010). Estes resultados contradizem a nossa hipótese, que, baseada no Efeito Inimigo Íntimo esperaríamos uma menor resposta agressiva entre operárias vizinhas (Yndenberg 1988, Temeles 1994). Temeles (1994) afirma que a ocorrência deste fenômeno depende do grau de ameaça que o vizinho representa, comparando a um estranho que por sua vez depende do tipo de território defendido.
território, ele representa ameaça e seus vizinhos respondem de forma mais agressiva (Booksmythe et al. 2010). Neste caso, a intensidade de agressão pode ser explicada pelo nível de ameaça representada pelo intruso em cada tipo de confronto que difere em função da motivação e valor do recurso (Booksmythe et al. 2010). Na formiga Pogonomyrmex barbatus Gordon (1989) descreve a ausência do fenômeno devido à maior competição alimentar entre vizinhas. Segundo Gordon (1989), indivíduos de colônias distantes podem ser forrageadores perdidos e não representam risco de invasão territorial.
Referência Bibliográfica
Araújo, A. & Rodrigues, Z. 2006. Foraging behavior of the queenless ant Dinoponera quadriceps Santschi (Hymenoptera: Formicidae). Neotropical Entomology, 35, 159-164.
Araújo, G. D. F. T., Malaquias, K. S., Bailez, O. E. & Viana-Bailez, A M. 2009. Ocorrência do fenômeno “Inimigo Próximo” em Atta sexdens rubropilosa (Hymenoptera, Formicidae). In: XIX Simpósio de Mirmecologia. Resumos. (Ed por Sociedade Brasileira de Mirmecologia). Ouro Preto: Editora UFOP.
Azevedo, D. L. O. 2009. O papel das rotas e obtenção de informações sobre a eficiência no forrageio de Dinoponera quadriceps em ambiente natural. Dissertação de Mestrado, Universidade Federal do Rio Grande do Norte.
Beye, M., Neumann, P., Chapuisat, M., Pamilo, P. & Moritz, R. F. A. 1998. Nestmate recognition and the genetic relatedness of nests in the ant Formica pratensis. Behavioral Ecology and Sociobiology, 43, 67-72.
Björkman-Chiswell, B. T., van Wilgenburg, E., Thomas, M. L., Swearer, S. E. &
Elgar, M. A. 2008. Absence of aggression but not nestmate recognition in an Australian population of the ant Linepithema humile. Insectes Sociaux, 55, 207-212.
Booksmythe, I., Jennions, M. D., Backwell, P. R. Y. 2010. Investigating the „dear enemy‟ phenomenon in the territory defense of the fiddler crab, Uca mjoebergi. Animal Behaviour, 79, 419-423.
Boulay, R. & Lenoir, A. 2001. Social isolation of mature workers affects nestmate recognition in the ant Camponotus fellah. Behavioral Processes, 55, 67-73.
effect? Animal Behaviour, 74, 985-993.
Bourke A. F. B. & Franks N. R. 1995. Social evolution in ants. 1a ed. New Jersey: Princeton University Press.
Brandstaetter, A. S., Endler, A. & Kleineidam. 2008. Nestmate recognition in ants is possible without tactile interaction. Naturwissenschaften, 95, 601-608.
Buczkowski, G. & Silverman, J. 2005. Context-dependent nestmate discrimination and the effect of action thresholds on exogenous cue recognition in the Argentine ant. Animal Behaviour,69, 741-749.
Buczkowski, G. & Silverman, J. 2006. Geographical variation in Argentine ant aggression behavior mediated by environmentally derived nestmate recognition cues. Animal Behaviour, 71, 327-335.
Caetano F.H., Jaffe K. & Zara F.J. 2002. Formigas: biologia e anatomia. Araras-SP: Gráfica e Editora Topázio.
Cammaerts, M. C., & Cammaerts, R. 1998. Marking of nest entrance vicinity in the ant Pheidole pallidula (Formicidae, Myrmicinae). Behavioral Processes, 42, 19-31.
Carlin, N. F. & Hölldobler, B. 1983. Nestmate and kin recognition in interspecific mixed colonies of ants. Science, 222, 1027-1029.
Corbara, B., Fresneau, D., Lachaud, J-P., Leclerc, Y. & Goodall, G. 1986. An automated photographic technique for behavioural investigations of social insects. Behavioral Processes, 13, 237-249.
Dimarco, R. D., Farji-Brener, A. G. & Premoli, A. C. 2010. Dear enemy phenomenon in the leaf-cutting ant Acromyrmex lobicornis: behavioral and genetic evidence. Behavioral Ecology, 21, 304-310.
Dunn, R. & Messier, S. 1999. Evidence for the opposite of the dear enemy phenomenon in termites. Journal of insect behavior, 12, 461-464.
Gamboa, G. F., Grudzien, T. A., Espelie, K. E. & Bura, E. A. 1996. Kin recognition pheromones in social wasps: combining chemical and behavioral evidence. Animal Behaviour, 51, 625-629.
Getty, T. 1989. Are dear enemy in a war of attrition? Animal Behaviour, 37, 337-338. Gordon, D. M. 1989. Ants distinguish neighbor from strangers. Oecologia, 81, 198-200.
Gordon, D. M. 2010. Ants encounters. Interaction, networs and colony behavior. Princenton: Princenton University Press.
Guerrieri, F. J. & D’Etorre, P. 2007. The mandible opening response: quantifying aggression elicited by chemical cues in ants. The Journal of Experimental Biology, 211, 1109-1113.
Heinze, J., Foitzik, S., Hippert, A. & Hölldobler, B. 1996. Apparent dear-enemy phenomenon and environment-based recognition cues in the ant Leptothorax nylanderi. Ethology, 102, 510-522.
Huntingford, F. A. & Chellappa, S. 2006. Agressão. In: Comportamento Animal. (Org. por M. E. Yamamoto & G. L. Volpato), pp. 157-174. Natal: Edufrn.
Huntingford, F. A. & Turner, A. K. 1987. Animal Conflict.1 ed. New York: Chapman and Hall Ltd.
Jutsum, A. R., Saunders, T. S. & Cherrett, J. M. 1979. Intraspecific aggression in the leaf-cutting ant Acromyrmex octospinosus. Animal Behaviour, 27, 839-844.
Knaden, M & Wehner, R. 2003. Nest defense and conspecific enemy recognition in the desert ant Cataglyfis fortis. Journal of Insect Behavior, 16, 717-729.
Krebs, J.R. & Davies, N. B. 1996. Introdução à Ecologia Comportamental. 3ª Ed. São Paulo: Atheneu.
Lahav, S., Soroker, V., Hefetz, A. & Meer, R. K. V. 1999. Direct behavioral evidence for hydrocarbons as ant recognition discriminators. Naturwissenschaften, 86, 246-249.
Langen, T. A., Tripet, F. & Nonacs, P. 2000. The red and black: habituation and the dear-enemy phenomenon in two desert Pheidole ants. Behavioral Ecology and Sociobiology, 48, 285-292.
Lenoir, A., Cuisset, D. & Hefetz, A. 2001. Effects of social isolation on hydrocarbon pattern and nestmate recognition in the ant Aphaenogaster senilis (Hymenoptra, Formicidae). Insectes Sociaux, 48, 101-109.
Liang, D. & Silverman, J. 2000. “You are what you eat”: Diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepithema humile. Naturwissenschaften, 897, 412-416.
Lumsden, C. J. & Hölldobler, B. 1983. Ritualized combat and intercolony communication in ants. Journal of Theorical Biology, 100, 81-98.
Martin, P. & Bateson, P. 2007. Measuring behaviour: an introductory guide. Cambridge: Cambridge University Press.
Matthews, R. W. & Matthews, J. R. 2010. Insect Behaviour. 2a ed. London: Springer. Medeiros, J. C. 2009. Influência do processo de fissão de colônia sobre comportamento
Trabalho de Monografia. Universidade Federal do Rio Grande do Norte.
Menzel, F., Schmitt, T. & Blüthgen, N. 2009. Intraspecific nestmate recognition in two parabiotic ant species: acquired recognition cues and low inter-colony discrimination. Insectes Sociaux, 56, 251-260.
Mercier, J. L., Lenoir, A. & Djean, A. 1997. Ritualised versus aggressive behaviours displayed by Polyrhachis laboriosa (F. Smith) during intraspecific competition. Behavioral Processes, 41, 39-50.
Monnin, T. & Peeters, C. 1997. Canibalism on subordinates‟ eggs in the monogynous queenless ant Dinoponera quadriceps. Short communications. Naturwissenschaften, 84, 499-502.
Monnin, T. & Peeters, C. 1998. Monogyny and regulation of worker mating in the queenless ant Dinoponera quadriceps. Animal Behaviour, 55, 299-306.
Monnin, T. & Peeters, C. 1999. Dominance hierarchy and reproductive conflicts among subordinates in a monogynous queenless ant. Behavioral Ecology, 10, 323-332.
Monnin, T. & Ratnieks, F. L. W. 1999. Reproduction versus work in queenless ants: when to join a hierarchy of hopeful reproductives? Behavioral Ecology and Sociobiology, 46, 413-422.
Monnin, T., Ratnieks, F. L. W. & Brandão, C. R. F. 2003. Reproductive conflict in animal societies: hierarchy length increases with colony size in queenless ponerine ants. Behavioral Ecology and Sociobiology, 54, 71-79.
Newey, P. S., Robson, KS. K. A. & Crozier, R. H. 2010. Weaver ants Oecoplylla smaragdina encounter nasty neighbors rather than dear enemies. Ecology. 9, 2366-2372
characters, and the importance of queen-worker dimorphism. In: The evolution of social behavior in insects and arachnids. (Org. por J. C. Choe & B. J. Cropi) Cambridge: Cambridge University Press.
Pirk, C. W. W., Neumann, P., Moritz, R. F. A. & Pamilo, P. 2001. Intranest ratedness and nestmate recognition in the meadow ant Formica pratensis (R.). Behavioral Ecology and Sociobiology, 49, 366-374.
Ratnieks, F. L. W., Foster, K. R., & Wenseleers, T. 2006. Conflict resolution in insect societies. Annual Review Entomoogy. 51, 581-608.
Roulston, T. H., Buczkowski, G., & Silverman, J. 2003. Nestmate discrimination in ants: effect of bioassay on aggressive behavior. Insectes Sociaux, 50, 151-159. Sanada-Morimura, S., Minai, M., Yokoyama, M., Hirota, T. Satoh, T. & Obara, Y.
2003. Encounter-induced hostility to neighbors in the ant Pristomyrmex pungens. Behavioral Ecology, 14, 713-718.
Soroker, V., Fresneau, D. & Hefetz, A. 1998. Formation of colony odor in Ponerie ant Pachycondyla apicalis. Journal of Chemical Ecology, 24, 1077-1090.
Sorvari, J., Theodora, P., Turillazzi, S., Hakkarainen, H. & Sundsteöm, L. 2008.Food resources, chemical signaling, and nest mate recognition in the ant Formica aquilonia. Behavioral Ecology, 19, 441-447.
Stuart, R. J. & Herbers, J. M. 2000. Nest mate recognition in ants with complex colonies: within- and between population variation. Behavioral Ecology, 11, 676-685.
Suarez, A. V., Tsuitsui, N. D. Holway, D. A., & Case, T. J. 1999. Behavioral and genetic differentiation between native and introduced populations of the Argentine ant. Biological Invasions, 1, 43-53.
Spatiotemporal patterns of intraspecific aggression in the invasive Argentine ant. Animal Behaviour, 64, 697-708.
Suwabe, M., Ohnishi, H., Kikuchi, T. & Tsuji, K. 2007. Nestmate discrimination in the queenless ponerine ant Diacamma sp from Japan. Entomological Science, 10, 7-10.
Tanner, C. J. & Adler, F. R. 2009. To fight or not fight: context-dependen interspecific aggression in competing ants. Animal Behaviour, 77, 297-305.
Temeles, E. J. 1994. The role of neighbours in territorial systems: when are they „dear enemies‟? Animal Behaviour, 47, 339-350.
Thomas, M. L., Tsutsui, N. D. & Holway, D. A. 2004. Intraspecific competition influences the symmetry and intensity of aggression in the Argentine ant. Behavioral Ecology, 16, 472-481.
Vasconcelos, A., Santana, G. G., Souza, A. K. 2004. Nest spacing and architecture, and swaring of males of Dinoponera quadriceps (Hymenoptera, Formicidae) in a remnant of the Atlantic Forest in Northeast Brazil. Braz. J. Biol,. 64, 357-362. Velásquez, N., Gómez, M., González, J. & Vásquez, R. A. 2006. Nest-mate
recognition and the effect of distance from the nest on the aggressive behavior of Camponotus chilensis (Hymenoptera: Formicidae). Behaviour, 143, 811-824. Wagner, D., Tissot, M., Cuevas, W. & Gordon, D. M. 2000. Harvester ants utilize
cuticular hydrocarbons in nestmate recognition. Journal of Chemical Ecology, 26, 2245-2257.
Ydenberg, R. C., Giraldeau, L. A. & Falls, J. B. 1988. Neighbours, strangers, and asymmetric war of attrition. Animal Behaviour, 36, 343-347.