INTRODUCTION
Glucokinase (GK) plays a key role in whole-body glucose homeostasis by catalyzing the phosphorylation of glucose in cells, such as pancreatic -cells and hepatocytes. Pancreatic GK acts as rate limiting enzyme for glucose utilization where it determines the rate of glucose-induced insulin secretion1 where as liver
GK determines the rate of glucose utilization and glycogen synthesis.2 These functions of GK
makes it a key metabolic regulator of blood glucose levels. In type 2 diabetes mellitus, hepatic glucose output is more than hepatic glucose utilization contributing to the elevated glucose levels in circulation.3
The mutations in GK results in its altered activities of glucose utilization causing abnormal glycemic control. This kind of loss and or alteration of the activity of GK by mutations is
linked up to maturity onset diabetes of the young 2 (MODY2), which is characterized by early onset and mild chronic fasting hyperglycaemia.4
MODY2 patients display an impaired glucose regulation with decreased accumulation of glycogen and increased hepatic glucose production.5 Mutations in GK causing MODY2
conditions results in decreased enzymatic activity due to reduction in its maximal velocity (Vmax) and or reduced affinity towards glucose
and adenosine triphosptate (ATP).6
A GK activator has the promise of potentially affecting both the -cells of the pancreas, by improving glucose sensitive insulin secretion, as well as the liver, by reducing uncontrolled glucose output and restoring post-prandial glucose uptake and storage as glycogen.7
Activation of GK results in a rise in insulin secretion from the pancreas and also stimulation of glucose uptake and glycogen synthesis in
Original Ariticle:
Molecular docking assessment of pyridone derivatives as
glucokinase activators
Y. Nanda Kumar,1 J.A. Pradeep Kiran,1 P. Santhosh Kumar,2 S.Yeswanth,2
K. Kalpana,1 V. Koteswara Rao,3 P.V.G.K. Sarma,2 M. Bhaskar1
1Division of Animal Biotechnology, Department of Zoology, Sri Venkateswara University, Tirupati and 2Department of Biotechnology, Sri Venkateswara Institute of Medical Sciences, Tirupati and 3Department of Pharmacology and Toxicology, University of Kansas, Lawrence, Kansas, USA
ABSTRACT
Background: Mutations in glucokinase (GK) gene results in maturity onset diabetes of the young 2 (MODY2). It has been observed that GK activators (GKAs) can activate GK structure and promote glucose phosphorylation and bring blood glucose levels to normal condition. The present study is aimed to identify the binding mode of pyridone deriva-tives (PDs) as GKAs through molecular docking study.
Methods: GK structure was retrieved from the Protein Data Bank (PDB), protonated and energy minimized. A database was constructed with 29 PDs and docked into the allosteric site specified with Y61, R63, S69 and Y215 residues using Molecular Operating Environment (MOE) software. Docking conformations were generated using triangle match algorithm and ranked by London dG scoring function. The binding orientations and strength of interactions were evaluated by ligand interaction module of MOE.
Results: Molecular docking of 29 PDs in allosteric site of GK gave reliable docking scores, interestingly arene cationic interactions were observed with the compounds PD1, PD12, PD20 and PD21. R63 residue of allosteric site played a predominant role in binding with PDs.
Conclusions: PDs can be potentially useful agents in future management strategies of type 2 diabetes mellitus.
Key Words:Glucokinase, Maturity onset diabetes of the young 2, Molecular docking, Glucokinase activators
Corresponding Author: Dr M. Bhaskar, Division of Animal Biotechnology, Department of Zoology, Sri Venkateswara
University, Tirupati 517 502, India. e-mail: [email protected]
Received: 7 June, 2012.
liver.8 These two functions lead to an increase
in glucose disposal and the return of blood sugar to basal levels. This provides a rational expectation that enhancement of GK activity would be a novel therapeutic strategy for type 2 diabetes mellitus. Consistent with this rationale, recently small molecules that are acting as allosteric modulators and can activate GK by binding to allosteric site on the GK have been discovered and its number has been enormously increasing.9,10 They have the ability
to enhance the GK activity and hence called as GK activators (GKAs). They can be the potential and promising molecules for an effective treatment of type 2 diabetes mellitus. To investigate the mechanism of action of GKAs, the mode of interaction between activator molecules and GK is a key aspect. Hence in the present study we describe the binding mode of a few recently published pyridone derivatives as GKAs11 by in silico
means to find out their efficacy to interact with the allosteric site of GK.
MATERIAL AND METHODS
All the computational studies were carried out using Molecular Operating Environment (MOE), 2011.10; Chemical Computing Group Inc. software.12
Protein processing and preparation
The X-ray Crystallographic structure of GK was retrieved from Protein Data Bank (PDB)13 (ID:
1V4S) with a resolution of 2.30 Å and loaded into MOE working environment ignoring water molecules and heteroatoms. The structure was protonated in generalized Born implicit solvated environment at a temperature of 300K, pH of 7 and a salt concentration of 0.1. Electrostatic potential was applied to a cut off value of 1.5 Å at a dielectric value of 1. A non-bonded cut off value of 8 Å was applied to the Leonard-Jones terms. After successful protonation, the structure was energy minimized in CHARM2714
force field applying a gradient cut-off value of 0.05. Kcal/mol/Å.
Molecular dynamics simulations15 were carried
out at a constant temperature of 300 K for a
heat time of 10 pico seconds and a total run time of 10 nano seconds. The temperature relaxation time was set to 0.2 pico seconds (ps) by the time step consideration of 0.001. The energy terms of GK were plotted as a graph to observe stability of the trajectories generated during simulations.
Construction and preparation of ligand database
A total of 29 pyridone derivatives (PDs) that can act as GKAs were obtained from literature survey11 and their three dimensional structure
were constructed in MOE working environment using MOE builder tool. A ligand database (.mdb file) was constructed using all the ligands and subjected to energy minimization in implicit solvated environment under AMBER9916 force
field. Related potential energy terms were enabled for all bonded and non bonded interactions at a gradient of 0.05 Kcal/mol/Å and force field partial charges were enabled during minimization process.
Allosteric site identification
GKAs can bind in the allosteric regulatory site situated near the catalytic site of GK and increase the rate of glucose phosphorylation. The allosteric site residues were identified from the previously reported glucose-GKA complex structures in PDB (IDs: 3VF6, 3ID8, 3H1V, 4DHY, 3IMX, 3F9M, 3S41, 3FRO, 3A0I and 3GOI). Their ligand plots were studied from PDBSum17 using their PDB IDs and the residues
that are in direct contact with GKAs were identified. The group of these residues was defined as binding site for the docking of ligands.
Molecular Docking
methodology and poses were generated by aligning the ligand triplets on the alpha sphere triplets of receptor. Thirty docking conformations were generated for each ligand and these conformations were ranked based on the free binding energies that were generated by London dG scoring function.18,19 The
conformations were refined and rescored in the same force filed to remove the duplicate conformations. From the final list of docked conformation the pose with least docking score was chosen for each ligand for the analysis.
Molecular dynamics of GK - PD24 complex
Among all the PDs, PD24 showed the best binding score with satisfactory hydrogen bond interactions. The strength of ligand to bind with GK and stability of this receptor-ligand complex was determined by molecular dynamics simulations. The glucokinase-pyridone derivative 24 (GK-PD24) docking confor-mation with lowest docking score was subjected to molecular dynamics simulation for a 10 ns of run time with the conditions specified above for GK simulations alone. The energy variations of the complex during simulations were plotted as graph to find out the stability of the complex through out the simulation period.
RESULTS
The three dimensional X-ray crystallographic structure of GK was obtained from PDB and the structure was optimized by energy minimization (Supplementary Figure 1). The molecular dynamics simulations of this structure have generated a total of 20000 conformations for a period of 10 ns. Their energy plot of the dynamics simulations showed that the energy fluctuated up to 2 ns, afterwards found to be stabilized and generated stable trajectories. This stability was found through out simulations after 2 ns (Supplementary Figure 2). The stabilized conformation was obtained at the end of the simulations is used for the docking purpose. The binding site residues were identified as Y61, R63, S69 and Y215 from the ligand plots of all GK entries with GKAs (Supplementary Figures 3-12).
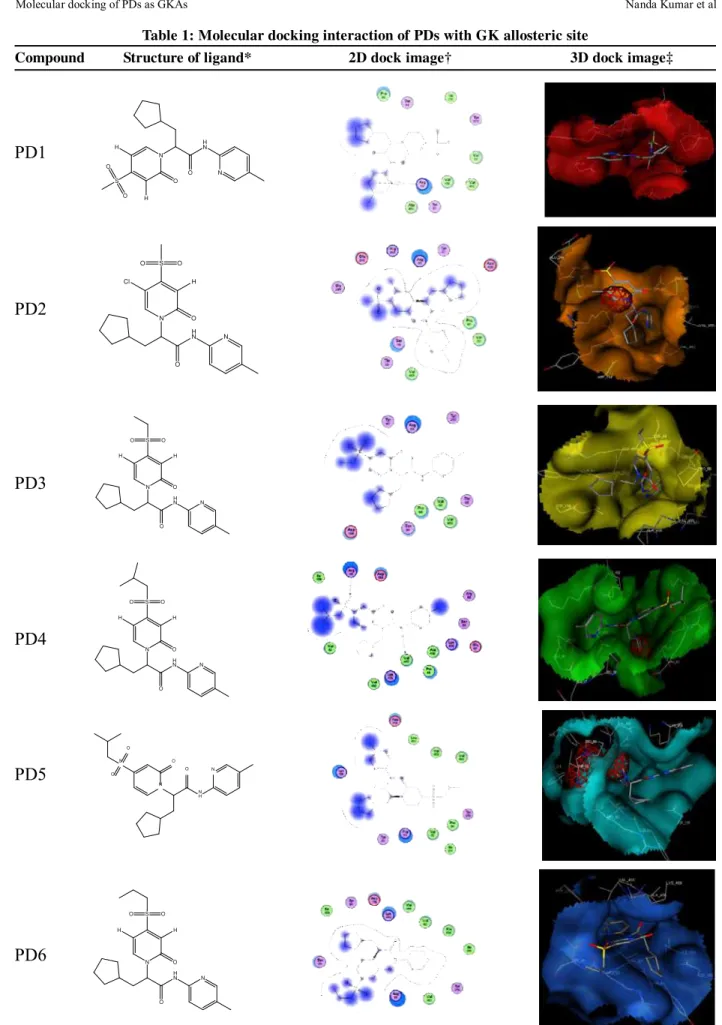
Table 1: Molecular docking interaction of PDs with GK allosteric site
Compound Structure of ligand* 2D dock image† 3D dock image‡
N H N
O N
S O
O H H
O
N H N
O N S
O O
H Cl
O
N H N
O N S
O O
H H
O
N H N
O N S
O O
H H
O
N
H N
O N
S
O O
O
N H N
O N S
O O
H H
O PD1
PD2
PD3
PD4
PD5
PD7
PD8
PD9
PD10
PD11
PD12
N H N
O N S
O O
H H
O
N H N
O N S
O O
H
O
N H N
O N S
O O
H Cl
O
N H N O
N
S
O O
H O
N H N
O N S
O O
Cl H
O
N
H N
O N
S
O O
PD13
PD14
PD15
PD16
PD17
PD18
N H N
O N
S O
O O
N H N
O N
S O
O O
N H N
O N
S O
O O H
F F
N H N
O N
S O
O O O
N H N
O N
S O
O O
N H N O N S
O
O
O H
PD19
PD20
PD21
PD22
PD23
PD24
N H N O N S
O
O O H
N H N
O N
S O
O O H
F F F
N H N
O N
N S
O
O
O H
H
N H N O
N
N S
O
O
O H
H
N H N
O N
N S
O
O O Cl
N H N
O N
N S
O
O
PD25
PD26
PD27
PD28
PD29
N H N
O S
O
O O H
N N
H
N H N
O S
O
O O H
N N
N H N
O S
O
O O H
N N
N H N
O S
O
O O H
N N
N H N
O S
O
O
O H
S N
*Structure of PDs, †two dimensional linear representation of docking conformations of respective
PDs with receptor. Arrow symbol indicates the hydrogen bond interactions and blue shaded region indicates the solvent contacts made by ligands. ‡Three dimensional graphical representations of docking conformations of respective PDs with receptor. The positional variations in the binding mode of PDs within the allosteric cavity of GK are depicted
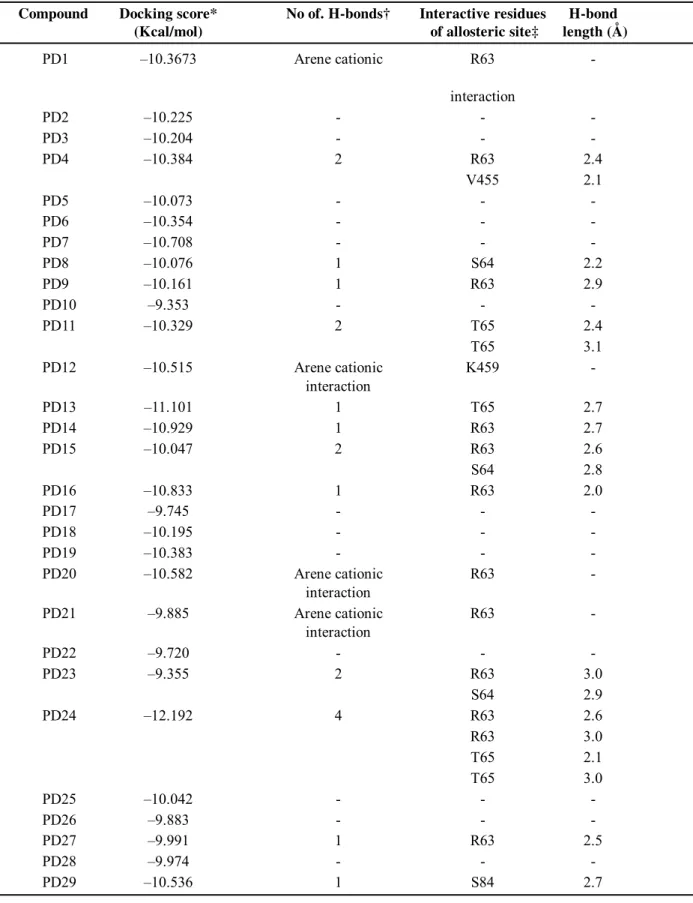
Table 2: Molecular docking information of PDs with GK allosteric site Compound Docking score* No of. H-bonds† Interactive residues H-bond
(Kcal/mol) of allosteric site‡ length (Å)
PD1 –10.3673 Arene cationic R63
-interaction
PD2 –10.225 - -
-PD3 –10.204 - -
-PD4 –10.384 2 R63 2.4
V455 2.1
PD5 –10.073 - -
-PD6 –10.354 - -
-PD7 –10.708 - -
-PD8 –10.076 1 S64 2.2
PD9 –10.161 1 R63 2.9
PD10 –9.353 - -
-PD11 –10.329 2 T65 2.4
T65 3.1
PD12 –10.515 Arene cationic K459
-interaction
PD13 –11.101 1 T65 2.7
PD14 –10.929 1 R63 2.7
PD15 –10.047 2 R63 2.6
S64 2.8
PD16 –10.833 1 R63 2.0
PD17 –9.745 - -
-PD18 –10.195 - -
-PD19 –10.383 - -
-PD20 –10.582 Arene cationic R63
-interaction
PD21 –9.885 Arene cationic R63
-interaction
PD22 –9.720 - -
-PD23 –9.355 2 R63 3.0
S64 2.9
PD24 –12.192 4 R63 2.6
R63 3.0
T65 2.1
T65 3.0
PD25 –10.042 - -
-PD26 –9.883 - -
-PD27 –9.991 1 R63 2.5
PD28 –9.974 - -
-PD29 –10.536 1 S84 2.7
*Docking scores of respective PDs and GK docking complexes. †The total number of hydrogen bonds formed be-tween the corresponding PDs and GK allosteric site residues. ‡The amino acid residues of allosteric site that are involved in the interaction with PDs.
were found to be present inside the cavity of the allosteric site indicating their role in the catalytic mechanism. The arene cationic interactions were found between the pyridine rings of PDs and the R-group of basic amino acid residues lysine (K459) and arginine (R63). Both of them are charged amino acid residues and these arene cationic interactions were found with the pyridine rings especially making solvent contacts. This might be a key factor for the induction of charge in the allosteric sight which in turn promotes the activation of GK and results in the phosphorylation of glucose in the catalytic site situated very near to the allosteric site. Finally it is clear that this docking study helped to predict the binding mode and their role in the activation of GK where their efficacy of activation was already proved9 but not the
binding mode and catalytic mechanism.
DISCUSSION
Blood glucose levels are dynamically regulated in the body via multiple mechanisms among which GK plays a major role in all the metabolically responsive organs like liver, pancreas, brain and gut.20 The X-ray crystal
structure of GK along with activators shows a palm shape topology where two unequal size of domains are observed that can be separated by a deep cleft. This cleft forms the active site for glucose phosphorylation and is situated about 20 Å away from the hydrophobic allosteric site. GKAs can bind to this allosteric site and activates GK.21 The docking studies of
present PDs as GKAs revealed the involvement of R63 as predominant residue in hydrogen bond formation with almost all ligands. Few of the ligands showed arene cationic interactions which contributed to the hydrophobic interaction with allosteric site. Ligands having such interactions with the residues like lysine and arginine shows stronger binding affinities and contribute to the potentiality of ligands. All the PDs in the present study have a pyridone core moiety in which the amide nitrogen shows high reactivity with allosteric site residues
forming hydrogen bond interactions and helps for the GK activation. Hence, it may be an ideal choice focusing the enzyme like GK having high control strength to maintain basal glucose levels and can also be a plausible rationale for narrowing the options to advance the diabetic therapy by means of GKAs. On the basis of the binding efficiency of present explained GKAs we can anticipate that they have the ability to better interact with the allosteric site there by enhancing the GK activity so as to maintain the glucose homeostasis. These results support the concept that GKAs represent a new class of compounds that increase both insulin secretion and hepatic glucose use by means of GK activation and in turn prove to be effective agents for the control and management of blood glucose levels in type 2 diabetes mellitus. The present study gave a better understanding and insight into the interaction of PDs as GKAs and also pointed out the contribution of arene cationic interactions and hydrophobic interactions by the aromatic pyridone structures to the docking score. Exploration of these ligand interactions expelled the key residues of allosteric site to activate GK. Finally it can be concluded that the present ligands could be the potential agents for activation of GK which is likely to be the future of management of type2 diabetes mellitus.
ACKNOWLEDGEMENTS
This work is supported by Department of Science and Technology (DST) INSPIRE Fellowship grant for supporting doctoral studies, INSPIRE Division, Department of Science and Technology, Government of India, New Delhi.
REFERENCES
1. Matschinsky FM. Banting Lecture 1995. A lesson
in metabolic regulation inspired by the glucokinase glucose sensor paradigm. Diabetes 1996;45:223-41.
2. Ferre T, Riu E, Bosch F, Valera A. Evidence from
3. Pal M. Medicinal chemistry approaches for glucokinase activation to treat type 2 diabetes. Curr Med Chem 2009;16:3858-74.
4. Vionnet N, Stoffel M, Takeda J, Yasuda K, Bell
GI, Zouali H, et al. Nonsense mutation in the glucokinase gene causes early-onset non-insulin-dependent diabetes mellitus. Nature 1992;356:721-2.
5. Clement K, Pueyo ME, Vaxillaire M,
Rakotoambinina B, Thuillier F, Passa P, et al. Assessment of insulin sensitivity in glucokinase-deficient subjects. Diabetologia 1996;39:82-90.
6. Davis EA, Cuesta-Muñoz A, Raoul M, Buettger
C, Sweet I, Moates M, et al. Mutants of glucokinase cause hypoglycaemia- and hyperglycaemia syndromes and their analysis illuminates fundamental quantitative concepts of glucose homeostasis. Diabetologia 1999;42:1175-86.
7. Bebernitz GR, Beaulieu V, Dale BA, Deacon R,
Duttaroy A, Gao J, et al. Investigation of functionally liver selective glucokinase activators for the treatment of type 2 diabetes. J Med Chem 2009;52:6142-52.
8. Liu S, Ammirati MJ, Song X, Knafels JD, Zhang
J, Greasley SE, et al. Insights into mechanism of glucokinase activation: observation of multiple distinct protein conformations. J Biol Chem 2012;287:13598-610.
9. Grimsby J, Sarabu R, Corbett WL, Haynes NE,
Bizzarro FT, Coffey JW, et al. Allosteric activators of glucokinase: potential role in diabetes therapy. Science 2003;301:370-3.
10. Matschinsky FM. Assessing the potential of glucokinase activators in diabetes therapy. Nat Rev Drug Discov 2009;8:399-416.
11. Pfefferkorn JA, Lou J, Minich ML, Filipski KJ, He M, Zhou R, et al. Pyridones as glucokinase activators: identification of a unique metabolic liability of the 4-sulfonyl-2-pyridone heterocycle. Bioorg Med Chem Lett 2009;19:3247-52. 12. Vilar S, Cozza G, Moro S. Medicinal chemistry
and the molecular operating environment (MOE): application of QSAR and molecular docking to
drug discovery. Curr Top Med Chem 2008;8:1555-72.
13. Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, Shindyalov IN, Bourne PE. The protein Data Bank. Nucleic Acids Res 2000;28:235-42.
14. Brooks BR, Bruccoleri RE, Olafson BD, States DJ, Swaminathan S, Karplus M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J Comput Chem 1983;4:187-217.
15. Zhang J, Li C, Chen K, Zhu W, Shen X, Jiang H. Conformational transition pathway in the allosteric process of human glucokinase. Proc Natl Acad Sci USA 2006;103:13368-73.
16. Hans JT, David J. Wiedenfeld, Jung GL, Richard AF. AMBER force field parameters for the trimethylanilinium cation. Bioorganic Med Chem Lett 1992;4:291-94.
17. Laskowski, RA. PDBsum: summaries and analyses of PDB structures. Nucleic Acids Res 2001;29:221-2.
18. Magdziarz T, Mazur P, Polanski J. Receptor independent and receptor dependent CoMSA modeling with IVE-PLS: application to CBG benchmark steroids and reductase activators. J Mol Model 2009;15:41-51.
19. Colotta V, Capelli F, Lenzi O, Catarzi D, Varano F, Poli D, et al. Novel potent and highly selective human A(3) adenosine receptor antagonists belonging to the 4-amido-2-arylpyrazolo[3,4-c]quinoline series: molecular docking analysis and pharmacological studies. Bioorg Med Chem 2009;17:401-10.
20. Schuit FC, Huypens P, Heimberg H, Pipeleers DG. Glucose sensing in pancreatic beta-cells: a model for the study of other glucose-regulated cells in gut, pancreas, and hypothalamus. Diabetes 2001;50:1-11.