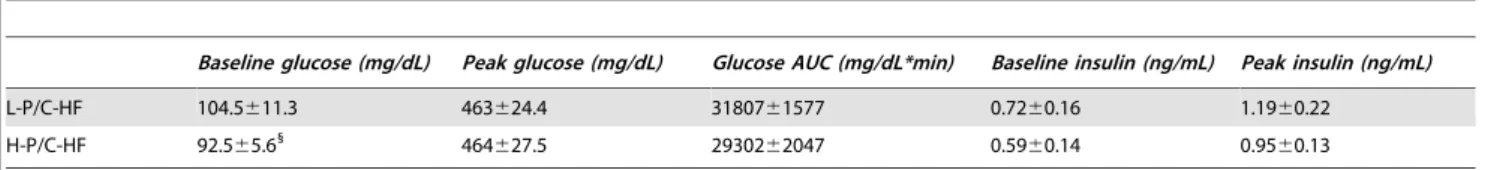
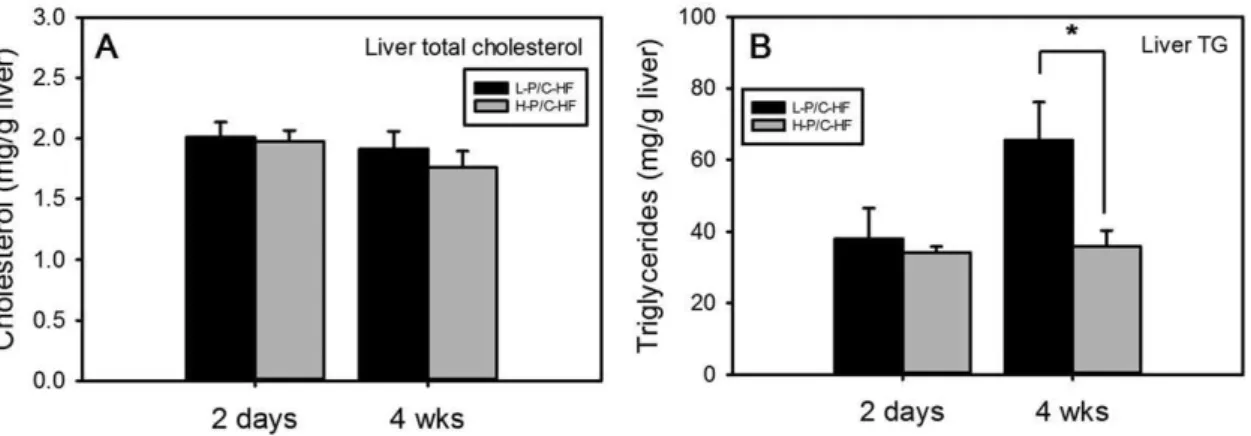
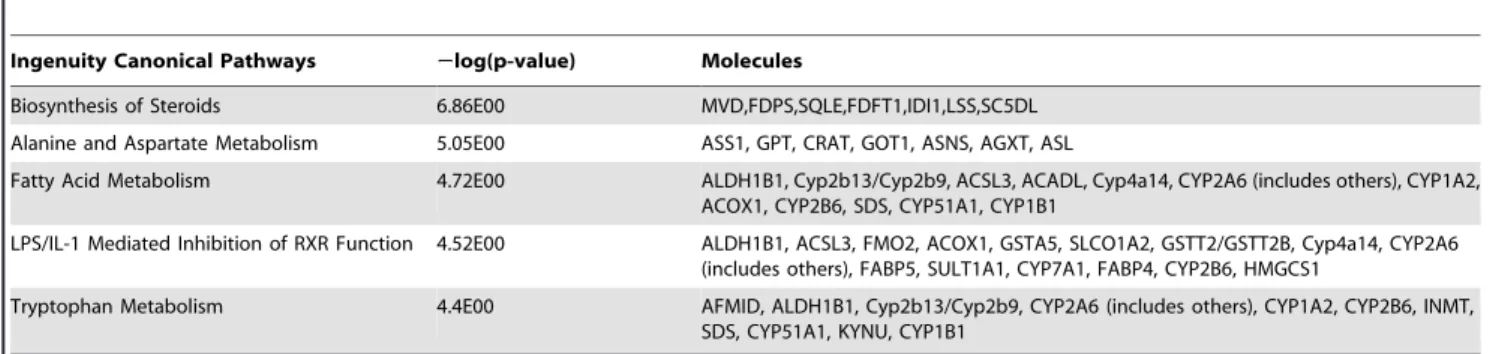
Consequences of exchanging carbohydrates for proteins in the cholesterol metabolism of mice fed a high-fat diet.
Texto
Imagem
Documentos relacionados
The aim of the present study was to assess the potential effects of flaxseed on atherogenesis in rabbits subjected to a high-cholesterol diet (containing 0.25% cholesterol) of
Observada a necessidade de melhoria no processo de obtenção e análise de dados fenotípicos em milho doce, os objetivos deste trabalho foram (i) comparar a eficiência de diferentes
(Nilsson et al ., 2012) The proteomic analysis of plasma from obese and thin rats fed with a high fat diet by Choi and Wang revealed that the change in the levels of twelve
Despite the high frequency of individuals with undesirable food habits, no association between lipoprotein alterations and high intake of total fat and cholesterol was observed..
se projetando numa hipotética pequena de Andrezinho 5. Nelson Rodrigues parece ter a intenção de tratar do homoerotismo logo no início do conto, quando Andrezinho, num dos
2: correlation between the number of Schistosoma mansoni eggs passed in faeces and the number of worm pairs recovered from mice fed high-fat diet (A) or standard diet
Effect of the ε 4 allele of apolipoprotein E on the levels of total cholesterol (TC), low-density lipo- protein cholesterol (LDLc), high-density lipoprotein cholesterol (HDLc),
In previous studies, we showed that the removal from the plasma of the free cholesterol (FC) label of a nano- emulsion that probes the low-density lipoprotein (LDL) metabolism
