1 ARTHUR SÉRGIO CAVALCANTI DE FRANÇA
ALTERAÇÕES DO CICLO SONO-VIGÍLIA MODULADAS POR DOPAMINA E SUAS CONSEQUENCIAS MNEMÔNICAS
Dissertação apresentada à Universidade Federal do Rio Grande do Norte como requisito para a obtenção do título de
Mestre em Psicobiologia.
2 ARTHUR SÉRGIO CAVALCANTI DE FRANÇA
ALTERAÇÕES DO CICLO SONO-VIGÍLIA MODULADAS POR DOPAMINA E SUAS CONSEQUENCIAS MNEMÔNICAS
Dissertação apresentada à Universidade Federal do Rio Grande do Norte como requisito para a obtenção do título de
Mestre em Psicobiologia.
Orientador: Sidarta Ribeiro Co-orientador: Bruno Lobão Soares
3 Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
França, Arthur Sérgio Cavalcanti de.
Alterações do ciclo sono-vigília moduladas por dopamina e suas consequências mnemônicas / Arthur Sérgio Cavalcanti de França. - Natal, RN, 2012.
77 f. : Il.
Orientador: Prof. Sidarta Ribeiro. Coorientador: Prof. Bruno Lobão Soares.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Sono-vigília – Dissertação 2. Memória – Dissertação. 3. Cérebro – Dissertação. I. Ribeiro, Sidarta. II. Soares, Bruno Lobão. III. Universidade Federal do Rio Grande do Norte. IV. Título.
5
AGRADECIMENTOS
Antes de tudo gostaria de agradecer aos meus pais. Eles são minha fonte diária de inspiração profissional e pessoal.
Gostaria de agradecer as minhas irmãs, Ananda e Bárbara, por todo apoio, incentivo e amor ao longo todo meu mestrado.
Gostaria prestar meus agradecimentos ao meu avô Cícero Cavalcanti. Dentre as mais diversas lembranças, a de um verdadeiro patriarca e a de um incentivador permanente de meus estudos.
Ao meu avô Otacílio França, um dos meus maiores símbolos de mudança, a certeza que as pessoas podem melhorar a cada dia que passa.
As minhas avós, Diva e Francisca, símbolos de afeição, compaixão, fé e amor fraternal.
A toda minha família que de alguma forma contribuiu para meu crescimento moral e pessoal. Sou ausente, mas amo todos de coração.
A minha “tropa”. Meus queridos amigos, companheiros para todas as horas e os irmãos que eu escolhi.
Aos meus orientadores Sidarta Ribeiro e Bruno Lobão Soares por me tratarem sempre com respeito, por me fazer sentir o que é ser um verdadeiro cientista e pelas lições que levarei por toda a vida.
6 Ao meu Sifu e mestre José Balestrini por todas as conversas que se tornaram uma grande lição de vida. Por ser um grande exemplo de dedicação, disciplina e sabedoria. Por me ensinar a extrair e dar o melhor de mim em todas as oportunidades possíveis.
A toda equipe do Instituto do Cérebro por criarem um verdadeiro “paraíso” científico no ambiente de trabalho, por fazerem uma simples conversa no momento de tomar café verdadeiras discussões científicas das mais abrangentes e variadas temáticas possíveis.
Agradecimento especial aos colegas Hindiael, Fábio, Vitor, Enio e Aron pelos incontáveis momentos de ajuda e pelas discussões que me ajudam a amadurecer dentro da ciência e no meu trabalho.
Ao George Nascimento, sem ele as coletas dos dados de eletrofisiologia seriam praticamente inviáveis.
A Larissa Muratori, minha parceira de laboratório ao longo de toda minha pesquisa.
7
SUMÁRIO
1 – Resumo ... 9
Abstract ... 10
2 – Introdução ... 11
2.1 - Sono ... 11
2.2 – Dopamina ... 13
2.3 – Memória ... 20
3 – Metodologia ... 25
3.1 - Animais ... 25
3.2 – Procedimento cirúrgico ... 25
3.2.1 – Manufatura da matriz de multi-eletrodos ... 26
3.2.2 – Cirurgia de implante ... 28
3.2.3 – Histologia ... 29
3.3 – Aparato experimental ... 29
3.3.1 - Objetos ... 30
3.4 – Grupos experimentais ... 30
3.5 - Procedimentos Pré-teste ... 32
3.6 - Teste de reconhecimento de objetos novos ... 32
3.7 - Tratamento ... 33
3.8 – Procedimento experimental ... 33
3.8.1 – Ambiente Experimental ... 33
3.8.2 – Animais implantados ... 34
3.9 - Análise de dados ... 35
3.9.1 – Comportamento ... 35
3.9.2 – Eletrofisiologia ... 35
4 - Resultados ... 38
4.1 – Animais Selvagens ... 38
4.1.1 – Injeção de haloperidol 0,3 mg/Kg imediatamente após a exploração ... Error! Bookmark not defined. 4.1.2 – Dupla sessão de objetos novos ... 38
8 4.1.3 – Injeção 0,05 mg/Kg imediatamente após exploração ... Error! Bookmark not defined.
4.1.4 – Injeção de 0,05 mg/Kg 30 minutos antes da exploração ... Error! Bookmark not defined.
4.2 – Animais heterozigotos ... 46
4.2.1 – Injeção de haloperidol 0,3 mg/Kg imediatamente após a exploração ... Error! Bookmark not defined. 4.2.2 – Injeção 0,05 mg/Kg imediatamente após exploração ... Error! Bookmark not defined. 4.2.3 – Injeção 0,05 mg/Kg 30 minutos antes da exploração ... Error! Bookmark not defined. 4.3 – Comparação entre grupos (Selvagens vs Selvagens; Heterozigotos vs Heterozigotos; Heterozigotos vs Selvagens) ... Error! Bookmark not defined. 4.3.1 – Comparação entre grupo submetido a injeção imediatamente após e seis horas após exploração ... Error! Bookmark not defined. 4.2.3 – Comparação entre os animais selvagens com injeção pré e pós exploração ... Error! Bookmark not defined. 4.2.3 – Comparação entre os grupos de animais heterozigotos ... 46
4.3.3 – Comparação entre todos os grupos de animais selvagens e heterozigotos ... 48
4.4 – Eletrofisiologia ... 49
4.4.1 – Parâmetros comportamentais ... 49
4.4.2 – Grupo controle ... 50
4.4.3 – Grupo Exposição ... 51
4.4.4 – Comparação Exposição Vs Controle ... 53
4.4.5 – Comparação de dados normalizados ... 56
4.4.6 – Curso temporal do Sono MRO ... 58
5 – Discussão ... 62
5.1 – Animais Selvagens ... 62
5.2 – Animais Heterozigotos ... 64
5.3 – Eletrofisiologia ... 66
6 – Conclusões ... 72
9
1 – RESUMO
10
ABSTRACT
11
2 – INTRODUÇÃO
Os fenômenos e processos que permeiam o sistema nervoso central dos animais mais recentes na escala filogenética são, em grande maioria, de grande complexidade no que concerne a atividade biológica de um organismo. As diversas áreas da ciência que têm o sistema nervoso como foco de estudo obtiveram um enorme avanço nas ultimas décadas, deste modo, o cérebro vêm deixando de ser “uma caixa preta” e começou a ter seus processos fisiológicos elucidados. Apesar da crescente interdisciplinaridade dentro das neurociências acarretar em avanços rápidos nas ultimas décadas, vários processos ainda estão longe de serem esclarecidos. Fenômenos como o sono, a consolidação de memórias, a comunicação e organização das redes neurais, a consciência, entre outros, são particularmente interessantes e complexos, e faz-se necessário trilhar um longo caminho para que sejam completamente compreendidos.
2.1 - SONO
12 obteve-se grandes avanços no que concerne a compreensão de seus processos fisiológicos, bioquímicos e biofísicos.
Nas últimas décadas diversos trabalhos em várias áreas da ciência têm mostrado a importância do sono em diversos processos e fenômenos que têm relação direta com o sistema nervoso central, seja para a homeostase ou para a manutenção de funções ligadas as oscilações circadianas. Como ilustração, podemos citar vários trabalhos que mostram o sono possuindo papel nos processos de aprendizado, formação e extinção da memória (Izquierdo e Medina 1997, Ribeiro et al 2002, 2004, 2004, 2007, Kalia 2006, McCarley 2007,). O trabalho de revisão de Ribeiro e Nicolelis (2004) relata que foram verificadas reativações de padrões de disparo neuronal no sono de padrões verificados em experiências adquiridas durante a vigília em roedores, primatas não humanos, humanos e pássaros. No mesmo trabalho foi destacado que a reativação das informações durante o sono seriam ligadas a aquisição das memórias em roedores e humanos.
O sono possui vários estágios, sendo os principais o sono de ondas lentas (SOL) e sono de movimento rápido dos olhos (MRO). O sono SOL é geralmente subdividido em três ou quatro subtipos de acordo com as ondas de eletroencefalograma (EEG) dominadas por ritmos de baixa frequência (abaixo de 4 Hz). Em mamíferos o sono MRO está relacionado aos ritmos teta (4 a 7 Hz), alfa (8 a 13 Hz) e gama (>30 Hz), mesmos ritmos que são encontrados na fase de vigília. O ritmo teta, tanto na vigília como durante o sono está associado à transferência de informações entre o hipocampo e o córtex cerebral (Buzsáki 2002, Buzsáki e Draguhn 2004).
13 neurônios do locus ceruleus durante o ciclo sono-vigília. Nesse trabalho foi verificada a flutuação nas taxas de disparos espontâneos em cada fase do ciclo.
A partir da década de 1970 vários trabalhos indicaram que a aprendizagem tem uma influência direta sob o ciclo de sono e vigília. O trabalho de Lucero (1970) com privação de alimento e exploração no labirinto em forma de estrela, revelou um aumento na duração do sono MRO nas primeiras duas horas após o teste em animais que aprenderam a localização do alimento. Nesse trabalho foram realizados registros de eletroencefalograma (EEG) e eletromiograma (EMG) até três horas após o procedimento de aprendizagem. O trabalho de Fishbein et al (1974) mostrou com dados de EEG e EMG que há um acréscimo no numero de episódios de sono MRO após uma tarefa de aprendizado de esquiva discriminativa. Resultado semelhante ao de Lucero foi encontrado pelo grupo de Smith (1982), nesse trabalho foi encontrado que os animais que passaram pela aprendizagem com esquiva inibitória teve um acréscimo de episódios de MRO nas primeiras quatro horas de registro. De modo geral, os trabalhos com tarefas com reforço negativo e positivo realizados em ratos levaram ao acréscimo de sono MRO, em número de episódios e a duração.
O trabalho de Schiffelholz e Aldenhoff (2002) mostrou resultado semelhante aos apresentados anteriormente, o seu diferencial foi mostrar que há o mesmo aumento específico nos episódios de MRO com uma tarefa que se baseia na tendência natural do animal de explorar novidade. Nesse trabalho foram coletados os dados de EEG com parafusos de aço inoxidável implantados em dez ratos Wistar. Apesar de a técnica não ser muito sofisticada e as análises serem pouco detalhadas, os resultados foram semelhantes aos dados prévios da literatura.
Trabalhos mais recentes vêm apontando, em complemento aos sistemas anteriormente comentados, a participação do sistema dopaminérgico na modulação do ciclo de sono e vigília, em particular a modulação das fases do sono (Lena et al 2005, Dzirasa et al 2006, Lu et al 2006, Monti e Monti 2007).
2.2 – DOPAMINA
14 grupamento hidroxila e origina a dopa. Através da enzima dopa descarboxilase a dopa perde um grupamento carboxila originando a dopamina, então pode ser utilizada tanto como neurotransmissor ou como precursor da noradrenalina e da adrenalina. Em uma típica sinapse dopaminérgica (figura 1) o neurotransmissor é liberado na fenda sináptica e age em receptores acoplados a proteína G, chamados de receptores metabotrópicos, tanto em neurônios pós-sinápticos como em neurônios pré-sinápticos. Esses receptores, quando ativados, abrem canais iônicos através de segundos mensageiros provocando a entrada de íons. No interior da membrana pré-sináptica a dopamina sofre a ação de uma enzima catabólica chamada de monoamina oxidase (MAO) que converte a dopamina em ácido dihidroxifenilacético (DOPAC).
Figura 1. Exemplo de uma sinapse envolvendo a dopamina como neurotransmissor. Nesta figura está ilustrada a síntese da dopamina a partir da tirosina, sua liberação, seu alvo de ação em receptores pós sinápticos, sua degradação por células da glia, a proteína DAT que realiza o transporte da dopamina para o neurônio pré sináptico e a ação da MAO dentro do neurônio.
15 finos (Colebrooke et al 2006) e sua porção ventral é associada com recompensa monetária e social (Tricomi et al 2010). Outro núcleo que possui neurônios dopaminérgicos em grande quantidade é o núcleo da área tegumentar ventral que projeta, principalmente, seus axônios para o sistema límbico e para o lobo frontal, em especial a região pré-frontal. Essa via está mais relacionada a processos emocionais e de tomada de decisão (Monti e Monti 2007, Cave e Baker 2009).
O sistema dopaminérgico possui atualmente 5 tipos de receptores descritos, divididos em duas famílias de receptores. A família D1 que inclui os receptores D1 e D5 e a família D2 que os inclui receptores D2, D3 e D4. Em geral, os receptores da família D1 são ativadores da adenilatociclase e os receptores da família D2 são inibidores da adenilatociclase, e ambos as famílias de receptores são expressas em todas as vias do sistema dopaminérgico, em quantidades diferentes, e possuem em geral efeito contrario na mesma via (Rang e Dale 2007, Stephen 2008 e Brunton et al 2005).
A dopamina está envolvida em diversas psicopatologias e neuropatologias. Um exemplo que pode ser citado é doença de Parkinson, síndrome neurodegenerativa que atinge principalmente os neurônios dopaminérgicos localizados na substância negra (Liou e Gallo 2009). Podemos falar também na esquizofrenia, doença que está relacionada com o desequilíbrio dos níveis dopaminérgicos da via mesolimbicacortical (Inta et al 2010).
16 provoca aumento da vigília e diminuição no estado de sono. Na ação farmacológica sobre o receptor do tipo D2 foi visto uma resposta bifásica, onde doses com pequenas concentrações de agonista D2 provocaram aumento de sono MRO e SOL e doses com concentração superiores resultaram em resposta contrária.
Os núcleos da rafe são regiões no tronco cerebral que possuem neurônios que produzem principalmente a serotonina. Há uma diminuição significativa na taxa de disparos dos neurônios dos núcleos da rafe durante o sono SOL e cessa durante o sono MRO. Por sua grande interconexão com sinapses dopaminérgicas Monti e Jantos (2008) propõem que a modulação dopaminérgica do sono se faz principalmente pela inervação aos núcleos da rafe (núcleos de neurônios serotonérgicos). Esse trabalho também apresenta mais dados farmacológicos, com resultados semelhantes ao artigo de Monti e Monti (2007), sobre o papel do receptor D2 sobre a modulação do aumento ou diminuição do sono MRO e SOL.
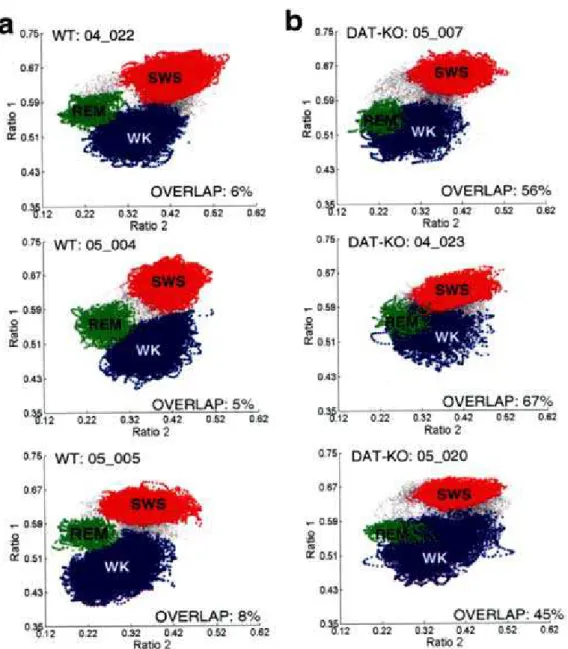
17 dos neurônios (Watanabe et al 2005), provocou uma redução drástica nos aglomerados de sono MRO em animais do tipo selvagem e acabou com os aglomerados de vigília, sono SOL e MRO nos animais DAT-Knockout (figura 3). O retorno específico do cluster de sono MRO só foi possível através de uma droga agonista inespecífica (L-dopa) ou de um agonista D2 (quipirole) (figura 4), revelando o papel importante da dopamina, particularmente os receptores da família D2, na modulação das fases do sono.
20 2.3 – MEMÓRIA
Como já mencionado, uma das possíveis funções do sono é a participação no processo de consolidação das memórias. O processo de aquisição, consolidação e evocação das memórias é um dos fenômenos mais importantes para os animais de modo geral, e é através desse fenômeno que os animais adquirem a capacidade de aprender novos modos de responder a estímulos ambientais, aprender novas atividades de cunho motor ou cognitivo tais como, reconhecer seus semelhantes, suas presas e evitarem seus predadores. Pode se dizer que, evolutivamente, o incremento crescente de capacidade de consolidação de memórias contribuiu para o aumento da plasticidade do comportamento dos animais que a possuem e com isso o aumento de sua adaptabilidade (Yamamoto e Volpato 2007).
O conceito de memória na literatura é bastante diverso, mas todas as definições permeiam a ideia de que se trata de um processo que envolve o armazenamento e aquisição de informação. O conceito de memória possui diversas classificações, como por exemplo, memória explicita ou declarativa que está relacionada a memórias de fatos ou eventos, memória implícita ou não declarativa que se relaciona aos condicionamentos bem como as memórias de procedimento. Outro sistema de classificação está relacionado ao tempo, essa classificação se diferencia em memórias de curto e longo prazo (Vianna et al 2000).
As memórias de curto e longo prazo são em geral hipocampo dependentes, ou seja, são necessários entradas neurais, processos eletrofisiológicos e moleculares do hipocampo para elas existirem. Podemos encontrar na literatura uma grande quantidade de teorias que envolvem o processo de consolidação de memórias. Desde de processos de transferência de informação por oscilações (Buzsáki 2002), processos moleculares de ativação de cascatas metabólicas (Izquierdo e Medina 1997), ou processos de fortalecimento ou enfraquecimento de sinapses (Whitlock et al 2006). Uma teoria não exclui a outra e é provável que todos os fenômenos participem de alguma forma na formação de memórias.
21 através de eletrodos na via perfurante, foi observado aumento dos potenciais pós sinápticos excitatórios das células hipocampais. Ao longo das quatro décadas seguintes vários avanços sobre o estudo do PLD e de sua inserção como modelo para memória e aprendizagem foram feitos, até que em 2006 o grupo de Mark Bear publicou um artigo sobre o achado do mecanismo de PLD, gerado pela aprendizagem, no hipocampo (Whitlock et al 2006).
O mecanismo de PLD, em síntese, pode ser descrito como um estado de otimização no processo de evocação de um potencial excitatório pós sináptico (PEPS), O PLD ocorre após sofrer um estímulo de alta frequência (estímulo tetânico). Ou seja, após a indução feita por um estímulo tetânico, a célula em estado de PLD é capaz de evocar um PEPS com maior facilidade do que antes do estado de PLD. A utilização dos conceitos de PLD e depressão de longa duração (DLD) para explicar a formação da memória segue os preceitos de Donald Hebb em que uma assembleia neural (formadoras de traços de memória) se mantém pelo fortalecimento e se desfazem pelo enfraquecimento das conexões entre os neurônios (Hebb 1949).
22 dopamina na manutenção de memórias condicionadas por medo através da modulação da produção de BDNF.
Como já evidenciado, o sono possui um papel essencial nos processos tanto de entrada de informação como na expressão de genes para plasticidade neural envolvidos na consolidação da memória. No trabalho de revisão de Ribeiro e Nicolelis (2004) sobre a relação das diferentes fases do sono com as etapas do processo de consolidação da memória, foi evidenciado que o sono SOL possui o papel de trocar e reverberar informações de diferentes áreas do cérebro adquiridas durante a fase de vigília. Ainda no mesmo trabalho foi discutido o papel do sono MRO no processo de consolidação da memória, é na fase do sono MRO que ocorreria a maior parte dos processos bioquímicos e fisiológicos para ativação da neuroplasticidade necessários para consolidação das memórias. No sono MRO, assim como na vigília, ocorre à ativação do gene imediato zif-268, esse gene é responsável por promover a ativação de uma cascata metabólica que inicia a produção de proteínas componentes de novos botões sinápticos para aumento do número de sinapses e fortalecimento das redes neurais envolvidas com as memórias adquiridas.
O trabalho de Eckel-Mahan et al (2008) mostra que, em ratos, as cascatas que envolvem CREB e AMPc no processo de consolidação da memória ocorre somente no período claro (período de maior descanso nos ratos), dando mais um indício de que o sono possa ter um importante papel na formação e consolidação de memórias de longo prazo.
A privação de sono, principalmente o estágio de sono MRO, está ligada diretamente ao prejuízo na aprendizagem ou formação de memórias. Trabalhos como os de Pearlman (1969) , Fishbein (1971), Smith e Kelly (1988) e Rolls et al (2011), usando desde técnicas simples, como a atonia encontrada no sono MRO para o animal acordar, até o uso de técnicas complexas, como a optogenética, levaram a resultados semelhantes em que a privação do sono MRO leva a um déficit na aprendizagem de tarefas como a esquiva inibitória. O trabalho de Palchykova et al (2006) mostra que a privação total de sono afeta a tarefa de reconhecimento em objetos novos.
23 et al (2007) discute a relação entre hiperdopaminergia, diminuição nos níveis de DLD hipocampais (utilizando eletrofisiologia in vitro) e perda na capacidade cognitiva (memória espacial no labirinto aquático), concordando com a dopamina e sua relação com o processo de aquisição e consolidação de memórias.
24
3 – OBJETIVOS
25
4 – METODOLOGIA
4.1 - ANIMAIS
Os animais utilizados no presente estudo são camundongos machos da espécie
Mus musculus tipo Black (figura 5). Foram utilizados 81 camundongos da cepa
selvagem na faixa etária de 3 a 6 meses. Foram utilizados 36 animais heterozigotos para o gene que transcreve a proteína transportadora de dopamina, com idade de 3 a 6 meses. Esses animais são hiperdopaminérgicos por não apresentarem um alelo codificante para o transportador que faz a retirada do neurotransmissor da fenda sináptica (Gainetdinov e Caron 2003). Foram utilizados animais do biotério de camundongos do Instituto Internacional de Neurociências de Natal Edmond e Lily Safra (IINN-ELS – Natal, RN) e do biotério de camundongos do Instituto do Cérebro (ICe-UFRN), ambos mantidos em um ciclo claro-escuro de 12 horas, sem restrição alimentar e de água. A temperatura do biotério foi mantida a 22°C ± 1°.
Figura 5. Camundongo da espécie Mus musculus tipo Black.
4.2 – PROCEDIMENTO CIRÚRGICO
26 4.2.1 – Manufatura da matriz de multi-eletrodos
A matriz de multi-eletrodos (figura 6) foi confeccionada manualmente com componentes pré-fabricados: conector da marca omnetics, placa PCB (Printed Circuit
Board-PCB) e eletrodos de tungstênio (diâmetro 50 micrômetros, impedância < 1
MOhm). O alinhamento de eletrodos foi realizado com gabarito de espaçamento de 300 micrômetros sob planejamento prévio através de atlas estereotáxico (Paxinos e Franklin 2001) (figura 7). O conector foi fixado na placa PCB através de solda de estanho e os eletrodos de tungstênio foram conectados com a placa através de tinta de prata. O revestimento utilizado para a matriz foi cola do tipo epóxi.
Figura 6. Matriz de multi-eletrodos. Podemos visualizar na figura o conector omnetcs (branco com o numero de identificação 1033), dois grupos de eletrodos e a placa PCB envolvida por epóxi.
28 4.2.2 – Cirurgia de implante
Durante o procedimento cirúrgico os animais foram mantidos anestesiados com isoflorano por via inalatória. Os animais foram posicionados em um aparelho estereotáxico (David Koppf). A craniotomia foi realizada com a referencia ao bregma na posição entre o córtex motor, o córtex somatosensorial e hipocampo, com as coordenadas 0,00 latero-lateral(LL), 0,75 Rostro-caldal (RC); 0,00 LL, 1,65 RC; -2,10 LL, 0,75 RC; -2,10 LL, 1,65 RC, em milímetros. Para fixar e aterrar o circuito da matriz, três parafusos foram aterrados com fio de prata e foram fixados em diferentes posições do crânio do animal. Dois eletrodos foram selecionados para registro de Eletromiograma (EMG), implantados no músculo trapézio do animal. Por fim, para cobrir a matriz foi feito um capacete no animal com acrílico. O head stage, aparelho que é acoplado ao conector na cabeça do animal, foi associado a um acelerômetro que informou por meio do sistema coletor de dados a movimentação do animal em três dimensões.
29 4.2.3 – Histologia
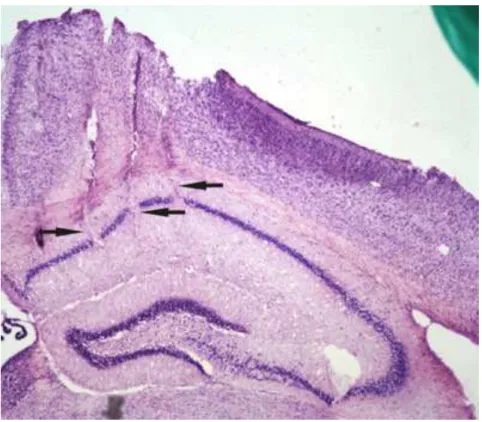
Para a verificação histológica do posicionamento dos eletrodos os cérebros foram removidos e armazenados a temperatura de -80ºC. A secção coronal do cérebro foi realizada a 30µm em um criostato (Zeiss). As secções foram dispostas em lâminas de vidro, seguindo uma distribuição em série, e posteriormente foram coradas com Níssil.
Figura 9. Corte histológico do hipocampo. A figura mostra três trilhas de eletrodos implantados no hipocampo (apontadas por setas).
4.3 – APARATO EXPERIMENTAL
30 4.3.1 - Objetos
No teste de reconhecimento de objetos novos foram utilizados seis itens para cada sessão de teste comportamental (doze itens foram utilizados para os animais que passaram por duas sessões). Os objetos utilizados no teste dos grupos comportamentais foram uma esponja de lavar louça foi colada em forma de V, uma pirâmide de acrílico azul, um cano de metal, a ponta cortada de uma escova, um rolo cilíndrico oco com fios de plástico em sua superfície externa (bob) e dois pedaços de borracha colados entre si em com forma de paralelepípedo oco.
Os animais que passaram pelo procedimento cirúrgico foram submetidos a duas sessões de exploração de objetos novos. Os objetos citados acima foram utilizados na primeira sessão de exploração de objetos. Na segunda sessão foram utilizados mais seis objetos, um rosca de plástico, uma pirâmide de borracha, um dado de pelúcia, uma clava de plástico, um tubo plástico com uma das pontas obstruída e uma bola de borracha.
Todos os objetos foram de comprimento e altura semelhantes, foram dispostos equidistantes entre si e a 5 cm das bordas para possibilitar a livre passagem do animal. Ambas as sessões possuíram dois objetos fixos e dois objetos móveis, um objeto de cada tipo foi destinado aos familiares e os outros dois para os não-familiares (Hughers 2007; Heyser e Chemero 2011).
4.4 – GRUPOS EXPERIMENTAIS
O presente trabalho possui vários grupos, do ponto de vista prático podemos dividir todos os animais utilizados em três grandes grupos experimentais. O primeiro com animais selvagens, segundo com animais heterozigotos para DAT-nocaute e o terceiro de animais implantados. Todos os grupos podem ser visualizados na tabela abaixo.
31 realizada imediatamente após o teste. O segundo procedimento foi realizado com uma sessão de exploração de objetos, também subdivididos em grupo tratado (halo 0,3 mg/Kg) e grupo controle (veículo) e foram submetidos a injeção seis horas após a exploração. O terceiro procedimento possuiu duas sessões de exploração de objetos, cada animal foi o controle dele mesmo. Na primeira sessão de exploração foi injetado veiculo e na segunda sessão foi injetado haloperidol (0,3 mg/Kg). O ultimo procedimento foi realizado com dois grupos. Os grupos tratado (haloperidol 0,05 mg/Kg) e controle (veículo) foram submetidos a injeção trinta minutos antes da exploração dos objetos.
O grupo de animais heterozigotos possuiu 36 animais em sua totalidade, divididos em dois procedimentos diferentes. No primeiro procedimento vinte e dois animais foram submetidos a uma sessão de exploração de objetos com injeção imediatamente após a exploração, os vinte e dois animais foram subdivididos nos grupos halo 0,3 mg/Kg, halo 0,05 mg/Kg e veículo. No segundo procedimento foram utilizados quatorze animais subdivididos nos grupos tratado halo 0,05 mg/Kg e grupo veiculo. A injeção foi realizada 30 minutos antes da exploração de objetos.
32 Tabela 1. Tabela esquemática dos grupos comportamentais. A tabela mostra que dose no procedimento de exploração de objetos novos os animais selvagens, heterozigotos e implantados foram submetidos.
3.5 - PROCEDIMENTOS PRÉ-TESTE
Os animais foram submetidos a 10 sessões de manipulação de cinco minutos, em dias consecutivos. Durante os experimentos não houve, com exceção da manipulação, nenhuma quebra na rotina dos animais, incluindo exposição a ambientes novos ou diferenciação no ciclo claro e escuro. As habituações ocorreram em sala fechada, iluminada e com temperatura de 23° Celsius.
3.6 - TESTE DE RECONHECIMENTO DE OBJETOS NOVOS
O teste de reconhecimento de objetos foi realizado no aparato supracitado (item 3.3). Este teste consiste na exposição a quatro objetos diferentes ao animal
Horário da injeção/ Dose e sessão
30 min. antes da exploração
Imediatamente após exploração
Seis horas após exploração
Halo 0,3 mg/Kg
Selvagens Heterozigotos
Implantados
Selvagens
Halo 0,05 mg/Kg Heterozigotos Selvagens
Heterozigotos Selvagens
Dupla sessão Selvagens
33 durante dez minutos; os objetos são colocados na mesma posição e equidistantes entre si para manter o padrão e evitar variações entre os animais. A injeção de veículo ou haloperidol foi aplicada intraperitoneal (IP)em três diferentes tempos: 1) logo após o a exposição dos objetos. 2) trinta minutos antes da exposição. 3) seis horas após a exposição dos objetos. 24 horas após a exposição são apresentados dois objetos novos e dois objetos anteriormente expostos. No caso dos grupos que são submetidos a duas sessões de exploração a segunda sessão é realizada sete dias após a primeira, repete-se o mesmo procedimento da primeira com todos os objetos diferentes da primeira sessão.
O teste se baseia na característica natural dos roedores de exploração de ambientes novos, portanto ao apresentar objetos novos os animais tendem a explorar significativamente mais os objetos novos em relação aos previamente expostos (Hughers 2007, Heyser e Chamero 2011). O teste utilizado nesse experimento foi adaptado de Ribeiro et al. (2004) para camundongos; o período de exposição foi adaptado para a realização dos testes durante o dia e o tempo de reexposição para 24 horas após a primeira exposição.
3.7 - TRATAMENTO
A dosagem da droga (haloperidol) usada nesse experimento foi baseada no trabalho de Dzirasa et al (2006). A concentração utilizada no trabalho foi de 0,3 mg/Kg e 0,05 mg/Kg, a droga foi diluída em água destilada.
3.8 – PROCEDIMENTO EXPERIMENTAL 3.8.1 – Ambiente Experimental
34 atenção do animal durante o teste. Após 10 minutos de exploração livre, os animais foram retirados do aparato. Após cada exploração o campo aberto e os objetos foram limpos com álcool 30%.
Os animais foram contidos sem auxilio de anestesia e foram submetidos a uma injeção intraperitoneal de veículo ou haloperidol. Transcorridas 24 horas após a exploração os animais foram reexpostos ao mesmo paradigma de objetos novos, com a presença de dois objetos não-familiares e dois familiares.
3.8.2 – Animais implantados
Os animais implantados são divididos em dois grupos. O primeiro grupo, chamado de controle, foi constituído por quatro animais que foram registrados em sua própria caixa durante 8 horas ciclo sono e vigília. Esses animais não passaram por nenhum procedimento comportamental. Foram submetidos a um registro após injeção de veículo e outro registro, sete dias após o primeiro, com injeção de haloperidol 0,3 mg/Kg.
35 3.9 - ANÁLISE DE DADOS
3.9.1 – Comportamento
Os dados foram coletados através de vídeo captado por uma câmera Panasonic acoplada ao sistema Plexon com o software da própria câmera. Os dados considerados nesse experimento foram a razão entre o tempo de exploração dos objetos familiares/não-familiares, o tempo total de exploração (familiares + não-familiares) e a atividade locomotora, todos coletados durante a reexposição.
Consideramos exploração quando o animal mantém contato com o objeto através de vibrissas e ou com as duas patas dianteiras. Qualquer outra parte do animal em contato com o objeto não é considerado no tempo de exploração (corpo, pata traseira ou cauda, por exemplo).
3.9.2 – Eletrofisiologia
36 Figura 10. Representação do mapa de estado do ciclo de sono e vigília. Os pontos pretos representam o cluster de vigília, os pontos verdes representam o aglomerado de sono MRO e os pontos vermelhos representam o aglomerado de sono SOL. Os pontos azuis representam a as faixas de transição do ciclo.
37 Figura 11. Histogramas associados a formação do mapa de estado. O eixo x e y de todos os quadros são equivalentes a razão 2 e 1, respectivamente, da figura 10. O quadro Mapa de Estado é um histograma da distribuição e intensidade dos pontos do mapa de estado. O quadro acelerômetro é uma distribuição de força da variação da aceleração no posicionamento do animal ao longo do registro. O quadro Teta e Delta representam uma distribuição de força das faixas de oscilação ao longo do registro.
Foram analisadas as durações de cada fase dos ciclos sono e vigília em quatro, seis horas de registro. Foi verificado também o curso temporal da fase de sono MRO em janelas de dez minutos durante as quatro primeiras horas de registros.
38
5 - RESULTADOS
5.1 – ANIMAIS SELVAGENS
5.1.1 – Parâmetros comportamentais na exploração de objetos realizada em duas sessões distintas
39
Veículo Halo 0,3 mg/Kg
0.0 0.5 1.0 1.5 2.0 2.5
**
A Nã o -f a m il ia r/ F a m il ia rVeículo Halo 0,3 mg/Kg
0 50 100 150 200
*
B F a m il ia re s + Nã o f a m il ia re sVeículo Halo 0,3 mg/Kg
0 10 20 30 C Di st â n c ia t o ta l pe rc o rr ida e m m e tr o s
Figura 12. A dose de 0,3 mg/Kg afeta a memória de reconhecimento de objetos. Primeira sessão de exploração de objetos com animais submetidos a injeção de veículo, a segunda sessão os animais foram submetidos a injeção de 0,3 mg/Kg haloperidol. A – Média da razão da duração de exploração entre objetos familiares e objetos não-familiares entre as duas sessões. B – Tempo total de exploração em segundos dos objetos familiares e não familiares. C – Atividade locomotora medida pela média da distância total percorrida no campo aberto. * p < 0,05, ** p < 0,01.
5.1.2 – Parâmetros comportamentais na exploração de objetos em animais selvagens submetidos a injeção trinta minutos e imediatamente após exploração
41
A
42
C
Figura 13. A dose baixa de haloperidol possui efeito contrário da dose alta no parâmetro de memória de reconhecimento de objetos. A – Comparação entre as médias da razões ente objetos não familiares e familiares. B – Comparação do tempo total de exploração de objetos. C – Comparação da distância total percorrida durante a exploração de objetos. * p < 0,05; ** p < 0,01; *** p < 0,001.
5.1.2 – Parâmetros comportamentais em animais submetidos a injeção seis horas após a exploração de objetos
43 da distância total percorrida dos grupos veículo (32.03 ± 2.96) e halo 0,3 mg/Kg (31.36 ± 2.81) revelou [t (0,16), p = 0,87].
Veículo Halol 0,3mg/Kg
0.0 0.5 1.0 1.5 2.0 2.5 A Nã o -f a m il ia r/ F a m il ia r
Veículo Halo 0,3mg/Kg
0 50 100 150 200 250 B F a m il ia re s+N ã o f a m il ia re s
Veículo Halo 0,3mg/Kg
0 10 20 30 40 C Di st â n c ia t o ta l p e rc o rr id a em m et ros
44 5.2.1 – Comparação de parâmetros comportamentais na exploração de objetos novos em animais submetidos a injeção imediatamente após e seis após a exploração
45 Pós-teste Seis horas
0.0 0.5 1.0 1.5 2.0 2.5 * A Nã o -f a m il ia r/ F a m il ia r
Pós-teste Seis horas
0 50 100 150 200 250 ** B F a m ili a re s + Nã o f a m ili a re s
Pós-teste Seis horas
0 10 20 30 40 Veículo Halo 0,3mg/Kg *** C Di st â n ci a t o ta l pe rc o rr ida e m m et ro s
46 4.2 – ANIMAIS HETEROZIGOTOS
4.2.3 – Parâmetros comportamentais na exploração de objetos em animais heterozigotos submetidos a injeção trinta minutos e imediatamente após exploração
47
A
48
C
Figura 16. A dose de 0,05 mg/Kg de haloperidol é capaz de modificar o padrão de reconhecimento de objetos novos em animais heterozigotos. A – Comparação entre as médias da razões ente objetos não familiares e familiares. B – Comparação do tempo total de exploração de objetos. C – Comparação da distância total percorrida durante a exploração de objetos. * p < 0,05, ** p < 0,01, ***p <0,001.
4.3.3 – O haloperidol difere na ação na comparação entre animais selvagens e animais heterozigotos
49 = 36,96, p < 0,0001] que indicou que a cepa possui efeito significativo sobre os dados.
Figura 17. O haloperidol pode bloquear, aumentar ou restabelecer a memória de reconhecimento de objetos novos. Comparação entre as médias da razão de exploração de objetos em animais selvagens comparados com animais heterozigotos. *** p <0,001.
4.4 – ELETROFISIOLOGIA
4.4.1 – Parâmetros comportamentais na exploração de objetos novos em animais implantados
50 menor que a primeira. No parâmetro de tempo total de exploração a comparação entre a sessão com veículo (118,5 ± 13,41) e a sessão com halo 0,3 mg/Kg (108,5 ± 28,61) revelou (p = 0,7128, t = 0,4047).
Veículo Halo 0,3mg/Kg
0.0 0.5 1.0 1.5 2.0 2.5 A
*
N ã o -f a m il ia r/ Fa m il ia rVeículo Halo 0,3mg/Kg
0 50 100 150 B Fa m il ia re s + N ã o f a m il ia re s
Figura 18. A dose de 0,3 mg/Kg de haloperidol afeta o reconhecimento de objetos novos em animais implantados. Primeira sessão de objetos submetidos a injeção de veículo, na segunda sessão foram tratados com haloperidol. A – Média da razão da duração de exploração entre objetos familiares e objetos não-familiares entre as duas sessões. B – Tempo total de exploração em segundos dos objetos familiares e não familiares. * p < 0,05.
4.4.2 – Análise da duração do ciclo sono e vigília no grupo controle
51 Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 50 100 150 200 250 A
6 horas 4 horas
Du ra ç ã o e m M in u to s
Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 10 20 30 40 B
6 horas 4 horas
Du ra ç ã o e m M in u to s
Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 50 100 150 200 C
6 horas 4 horas
Du ra ç ã o e m M in u to s
Figura 19. Comparação da duração de cada fase do ciclo sono e vigília entre as seis e quatro primeiras horas de registro no grupo controle. A – Comparação entre as médias da duração de sono SOL nas primeiras seis e quatro horas de registro. B – Comparação entre as médias da duração de sono MRO nas primeiras seis e quatro horas de registro C – Comparação entre as médias da duração de vigília nas primeiras seis e quatro horas de registro. Todas as comparações foram realizadas com teste t não paramétrico de Mann Whitney.
4.4.3 – Análise da duração do ciclo sono e vigília no grupo exposição
52 para as quatro primeiras horas, tal resultado indica que os animais com tratamento de haloperidol 0,3 mg/Kg possuem uma média significativamente menor de sono MRO nas quatro primeiras horas de registro comparada ao seu grupo controle e uma tendência nas primeiras seis horas de registro. Para a fase de vigília foi revelado (p = 0,34, U = 7,5) nas seis primeiras horas e (p = 0,75, U = 10,5) para as quatro primeiras horas. Os resultados podem ser vistos na figura 20.
Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 50 100 150 200 A
6 horas 4 horas
Du ra ç ã o e m M in u to s
Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 20 40 60
B
6 horas 4 horas
* Du ra ç ã o e m M in u to s
Veículo Halo 0,3mg/Kg Veículo Halo 0,3mg/Kg
0 50 100 150
C
6 horas 4 horas
Du ra ç ã o e m M in u to s
53 4.4.4 – Análise da duração do ciclo sono e vigília na comparação entre os grupos exposição Vs controle
54 Exposição Naive 0 50 100 150 200 250 Veiculo Haloperidol A Dur a ç ã o e m M inut os Exposição Naive 0 20 40 60 Veiculo Haloperidol ** B Dur a ç ã o e m M inut os Exposição Naive 0 50 100 150 200 Veiculo Haloperidol C Dur a ç ã o e m M inut os
Figura 21. Comparação através da Anova de duas vias entre as condições de controle/exposição e o tratamento veículo/halo 0,3 mg/Kg nas seis primeiras horas de registro. A – Comparação entre as médias da duração de sono SOL entre os grupos controle/exposição e seus respectivos tratamentos. B – Comparação entre as médias da duração de sono MRO entre os grupos controle/exposição e seus respectivos tratamentos C – Comparação entre as médias da duração de vigília entre os grupos controle/exposição e seus respectivos tratamentos *p < 0,05, **p < 0,01.
55 efeito da condição sobre os dados. Por ultimo, a comparação da duração da vigília nas entre os grupos veículo exposição (63,20 ± 8,732), tratado exposição (69,00 ± 6,760), veículo controle (104,0 ± 18,33) e tratado controle (69,50 ± 8,312) revelou um (p = 0,084, F = 3,46) para interação entre os dados, (p = 0,2064, F = 1,76) para o efeito do tratamento sobre os dados e (p = 0,0773, F = 3,63) para o efeito da condição sobre os dados. Os resultados podem ser vistos na figura 22.
Exposição Controle 0 50 100 150 Veiculo Haloperidol A Dur a çã o e m M in u to s Exposição Controle 0 10 20 30 40 Veiculo Haloperidol *** * * B Dur a çã o e m M in u to s Exposição Controle 0 50 100 150 Veiculo Haloperidol C Dur a çã o e m M in u to s
56 4.4.5 – Comparação de dados normalizados
Para verificar apenas a variação dos dados do grupo tratado em relação ao controle, os dados foram normalizados em relação ao veículo (veículo/veículo, Haloperidol/veículo), dessa forma os dados dos grupos controle foram transformados no valor de 1 e o grupo de haloperidol se transforma na variação que possui do veículo. Com esse tratamento a condição de exposição e controle é anulada e os dados podem ser juntados aumentando o poder da estatística. A comparação de todos os dados foi feito com o teste não paramétrico e pareado de Wilcoxon. Comparando a variação do sono SOL encontramos uma média de (1,311 ± 0,2080) para o grupo tratado, o teste t pareado revelou (p = 0,3594, W = -17,0). Na comparação do sono MRO (0,6792 ± 0,1068) o teste revelou (p = 0,0273, W = 37,0). O teste com a vigília (1,005 ± 0,1806) revelou (p = 0,5703, W = 11,0). Os resultados podem ser visualizados na figura 23.
Veiculo Halo 0.3mgKg 0.0 0.5 1.0 1.5 2.0 A Mé d ia d a r a z ã o nor m a li z a da
Veiculo Halo 0.3mgKg 0.0 0.5 1.0 1.5 * B Mé d ia d a r a z ã o nor m a li z a da
Vigilia Halo 0.3mgKg 0.0 0.5 1.0 1.5 C Mé d ia d a r a z ã o nor m a li z a da
57 Comparação das médias normalizada da duração de sono MRO dos grupos de veículo e halo 0,3 mg/Kg. C – Comparação das médias normalizada da duração da vigília dos grupos de veículo e halo 0,3 mg/Kg. *p < 0,05.
Para verificar se houve diferença significativa na comparação da variação dos grupos haloperidol na condição de controle e exposição foi realizado o teste estatístico não paramétrico Mann Whitney. Na comparação entre as médias normalizadas do grupo halo controle (1,828 ± 0,30) e o grupo halo exposição (0,8972 ± 0,074) no sono SOL revelou (p = 0,0159, U = 0,0). Na comparação entre os grupos halo controle (0,5698 ± 0,1814) e o grupo halo exposição (0,7668 ± 0,1305) no sono MRO foi revelado (p = 0,4127, U = 6,0). Por ultimo, a comparação entre os grupos halo controle (0,7354 ± 0,1566) e o grupo halo exposição (1,220 ± 0,2767) na vigília revelou (p = 0,1905, U = 4,0), resultados podem ser vistos na figura 24.
Controle Exposição 0.0 0.5 1.0 1.5 2.0 2.5 * A Mé d ia d a r a z ã o nor m a li z a da Controle Exposição 0.0 0.2 0.4 0.6 0.8 1.0 B Mé d ia d a r a z ã o nor m a li z a da Controle Exposição 0.0 0.5 1.0 1.5 2.0 C Mé d ia d a r a z ã o nor m a li z a da
58 Comparação das médias normalizada da duração de sono MRO entre os grupos tratado na condição controle e exposição. C – Comparação das médias normalizada da duração da vigília entre os grupos tratado na condição controle e exposição. *p < 0,05.
4.4.6 – Curso temporal do Sono MRO
61 4.4.6 – A duração de sono MRO é correlacionado com a razão de exploração de objetos.
Para verificar se há correlação entre os dados do parâmetro comportamental de razão na exploração de objetos e os dados de duração de sono MRO nas quatro primeiras horas de registro, foi realizado uma correlação de Pearson. O teste revelou [r = 0,85, p = 0,006]. Foi realizado uma análise de regressão linear afim de verificar se a razão na exploração de objetos poderia predizer a duração do sono MRO. Foi revelado [r² = 0,73, p = 0,006].
Figura 26. A razão de exploração de objetos pode predizer a duração de sono. Correlação de Pearson e regressão linear com os dados de duração de sono MRO e
62
6 – DISCUSSÃO
5.1 – ANIMAIS SELVAGENS
A dopamina possui papel modulatório em diversas funções exercidas pelo encéfalo. Como já mencionado na introdução, a dopamina desempenha um papel importante desde a modulação do sistema límbico até atividade motora no controle de movimentos finos. (Colebrooke et al 2006, Tricomi et al 2010, Monti e Monti 2007, Cave e Baker 2009). A dopamina também participa na modulação do processo de consolidação de memórias (Morice et al 2007). Devido à atuação da dopamina em diversos processos cognitivos, todos os grupos comportamentais obtiveram um controle para servir de indicativo na mudança da atividade locomotora (distancia total percorrida), interesse na exploração dos objetos (tempo total de exploração dos objetos) e a medida indireta de consolidação de memória (razão entre a exploração de objetos não familiares/familiares).
No parâmetro da razão na exploração dos objetos obtivemos resultado comum nos dois grupos com injeção imediatamente após a exploração tratados com 0,3 mg/Kg de haloperidol. Observamos uma diferença estatística significante entre os grupos tratado e veículo, os grupos de animais selvagens tratado com 0,3 mg/Kg obtiveram médias da exploração de objetos próximas a 1 e os animais controles desses grupos obtiveram médias próximas a 2 (13A). Tais resultados indicam que quando a razão está próxima a 1 os animais exploram de forma semelhante e indiferenciada os objetos não familiares e familiares. A falta de distinção entre os objetos é um indicativo que há um possível prejuízo nos níveis de consolidação de memória do animal no parâmetro, já que o teste se baseia na característica natural dos roedores de exploração de ambientes novos; portanto, ao apresentar objetos novos os animais tendem a explorar significativamente mais os objetos não familiares em relação aos previamente expostos (Hughers 2007, Heyser e Chamero 2011).
63 medo condicionado. É sugerido que por esse trabalho que a ação sob a consolidação dessas memórias se deu através da modulação na produção de BDNF. Outro exemplo encontrado na literatura é o trabalho de Morice et al (2007). Nesse trabalho é descrito o déficit na memória no parâmetro comportamental do water maze sob a manipulação farmacológica com haloperidol e também com o uso de animais DAT-nocautes. No mesmo trabalho foi proposta a hipótese de que tal déficit seja devido a diminuição significativa do mecanismo de DLD no hipocampo. É importante ressaltar no caso que tais dados eletrofisiológicos foram encontrados com experimento in-vitro com fatias de hipocampo. A modulação do dopaminérgica realizada com agonistas dopaminérgicos D2 apontou também prejuízos na consolidação de memória no medo condicionado (Nader e LeDoux 1999), bem como para a facilitação da extinção da memória de medo condicionado (Ponnusamy et al 2012). O resultado visto no presente trabalho vai de acordo com resultados já descritos na literatura. Indicando que o bloqueio parcial da atividade dopaminérgica em receptores da família D2 realizada pelo haloperidol (0,3 mg/Kg) pode estar ligada a falha no processo de consolidação da memória.
Resultado semelhante foi encontrado no parâmetro de tempo total de exploração. O grupos vistos na (figura 13) obtiveram, respectivamente, uma tendência e uma diferença significativa no tempo em que exploraram os objetos. O haloperidol é uma droga que atua em todas as vias dopaminérgicas (Niemegeers e Laduron 1986), podendo então interferir na atividade locomotora, no sistema de recompensa e punição ou em processos emocionais e tomada de decisão (Colebrooke et al 2006, Tricomi et al 2010, Monti e Monti 2007, Cave e Baker 2009). Podemos sugerir que o resultado se deu devido a atuação do haloperidol nas vias ligados a processos emocionais ou do sistema de recompensa, de forma que os animais não obtiveram tanto interesse em explorar os objetos.
64 O haloperidol é uma droga que primariamente atua sobre os receptores da família D2, mas em baixas doses ele parece ter afinidade com receptores da família D1 (Brunton et al 2005). O fato de os receptores das famílias D1 e D2 atuarem, em geral, de forma antagônica nas vias dopaminérgicas (Rang e Dale 2007, Stephen 2008 e Brunton et al 2005) pode justificar o fato de o grupo com injeção imediatamente após não ter afetado significativamente os níveis de consolidação de memória no teste de objetos novos dos camundongos.
Quando o grupo halo 0,05 mg/Kg submetido a injeção trinta minutos antes da exploração foi comparado com os demais grupos submetidos a injeção imediatamente após a exploração (figura 13) foi revelado que sua média é significativamente maior que os outros grupos. Podemos sugerir que quando a droga atua durante a aquisição e consolidação da memória em baixas doses de haloperidol é capaz de aumentar o desempenho na exploração de objetos novos.
No parâmetro de tempo total de exploração a dose de 0,05 mg/Kg atuou de forma semelhante ao parâmetro anterior. No grupo com injeção imediatamente após exploração os animais do grupo tratado obtiveram uma média significativamente maior do que o grupo veículo. Dessa forma a dose baixa de 0,05 mg/Kg se comportou de forma antagônica a dose alta de 0,3 mg/Kg. O resultado fica mais evidente como é mostrado na figura 13, onde os grupos veículo e halo 0,3 mg/Kg foram significativamente menores na média do tempo total de exploração. Assim, pode-se sugerir que a mesma argumentação da atuação antagônica do haloperidol dada ao parâmetro de memória pode ser aplicada ao tempo total de exploração.
No parâmetro de distância total percorrida não foi obtido nenhuma diferença estatística entre os animais tratados e seus respectivos grupos controle (figura 13). O mesmo resultado foi obtido com a comparação entre grupos. Em geral o haloperidol é utilizado para provocar déficits motores agudos por um curto período de tempo, de modo que seu efeito sobre a via nigro-estriatal não perdura por muito tempo após a aplicação (Rang e dale 2007, Durleux et al 2011).
O resultado do grupo submetido a injeção de 0,3 mg/Kg seis horas após a exploração vai ser discutido junto dos dados de eletrofisiologia.
65 Os animais heterozigotos submetidos a injeção de halo 0,3 mg/Kg e 0,05 mg/Kg imediatamente após a exploração exibiram um comportamento semelhante. No parâmetro da razão entre a exploração dos objetos os animais do grupo controle e dos tratamentos obtiveram razões próximas a 1. Esse resultado indicou que mesmo sem intervenção farmacológica os animais heterozigotos para o gene de DAT possuem um déficit natural na tarefa de reconhecimento em objetos novos. O resultado também revelou que as doses de 0,3 e 0,05 mg/Kg de haloperidol não modificaram a tendência a não reconhecer os objetos novos.
A justificativa para os animais heterozigotos possuírem um déficit natural na tarefa pode estar no desequilíbrio nos níveis de dopamina encontrados por todas as vias dopaminérgicas no cérebro desses animais. Os animais Nocautes possuem um déficit na produção de dos transportadores de dopamina da fenda sináptica, isso resulta na permanência do neurotransmissor na fenda sináptica 100X mais do que animais selvagens (Giros et al 1996). O excesso de dopamina na fenda sináptica aumenta a atividade dos neurônios pré e pós sinápticos, esse excesso de atividade pode estar ligado ao déficit na memória encontrado no presente trabalho.
O grupo que foi submetido a injeção (0,05 mg/Kg) trinta minutos antes da exploração revelou uma média significativamente maior que seu grupo controle. Esse resultado mostrou que a dose de 0,05 mg/Kg atuando não só na consolidação, mas também na aquisição resultou em um exploração dos objetos novos significativamente maior que os velhos. O resultado visto na figura 16 revela que a razão do grupo de heterozigotos ficou estatisticamente semelhante à encontrada nos animais selvagens.
Resultado semelhante foi encontrado no parâmetro de tempo total de exploração, podemos ver nas figuras 16 que os grupos que foram submetidos a injeção imediatamente após a exploração não obtiveram diferença estatística entre os grupos controle e tratado. O resultado entre o grupo submetido a injeção trinta minutos antes da exploração figura 16 também não revelou diferença estatística entre o controle e o grupo tratado, bem como não houve diferença entre grupos.
66 um aumento significativo da média da distância total percorrida, na figura 16 podemos visualizar que a média da distância total percorrida no grupo halo 0,05 mg/Kg, submetido a injeção 30 minutos antes da exploração, é significativamente maior que os grupos 0,3 mg/Kg e do veículo submetidos a injeção trinta minutos após a exploração
Curiosamente, o haloperidol agiu de forma semelhante no parâmetro de consolidação de memória em animais selvagens e animais heterozigotos. Os animais selvagens, no entanto, verificou-se também alterações no o tempo total de exploração enquanto nos animais heterozigotos foi afetado o parâmetro de distância total percorrida.
5.3 – ELETROFISIOLOGIA
O resultado do teste comportamental dos animais que passaram pela eletrofisiologia, visto na figura 18, foi semelhante aos animais que passaram pele mesmo procedimento, injeção de 0,3 mg/Kg de haloperidol imediatamente após a exploração. Assim como os resultados anteriores o grupo veículo ficou com a média da razão próxima a 2 e o grupo haloperidol ficou com a média próxima a 1, indicando que em média os animais não distinguiram os objetos após a utilização da droga. No parâmetro de tempo total de exploração os grupos não diferiram na comparação.
Os dados de PLC foram tratados de modo a fornecer a variação e duração de tempo em que os animais passaram em cada fase do ciclo sono e vigília. No presente trabalho foi explorado as diferenças na duração de tempo entre cada fase do ciclo sono e vigília com o tempo total de seis e quatro horas de registro.
67 quatro horas de registro (p = 0,02). A comparação entre os outros estados não revelou tendência ou diferença estatística.
Para comparar se as condições de exposição ou controle em que os animais foram submetidos são capazes de inteferir no ciclo de sono e vigília o grupo exposição e o grupo controle foram comparados entre si. Com a comparação de seis horas de registro (figura 21) pôde ser verificado que o sono SOL e a vigília não são afetados nem pelo tratamento da droga e nem pela condição de exposição de objetos novos. O teste de Anova de duas vias revelou que a condição (exposição e controle) influência significativamente o resultado, isso sugere que a exposição de objetos novos leva a um aumento significativo e específico de sono MRO. O resultado da análise das quatro primeiras horas de registro (figura 22) revelou a mesma tendência da análise das seis primeiras horas de registro. A vigília e o sono SOL não revelou diferenças significantes enquanto o sono MRO revelou diferença no efeito da condição e do tratamento sobre os dados. Deste modo foi possível verificar que houve efeito específico da droga haloperidol na modulação da fase de sono MRO com um efeito de diminuição da duração do mesmo em relação aos seus respectivos controles.
68 enquanto o grupo de exposição teve uma tendência a ter um aumento relativo de vigília.
Foi analisado o curso temporal de sono MRO para verificar se há diferença, entre as diferentes condições e tratamentos, no modo em que se comporta a duração de sono MRO ao longo do registro. Como pode ser visualizado na figura 25, foi verificado que as diferenças entre o tratamento (veículo e halo 0,3 mg/Kg) no grupo controle e no grupo exposição foi obtida entre a segunda e terceira hora de registro. A comparação entre todos os grupos (halo controle, halo exposição, veículo controle e veículo exposição) revelou também uma diferença na segunda e terceira hora.
O sono possui características fundamentais para o processo de consolidação da memória, seja em processos de plasticidade neural como também no fluxo de troca de informações entre várias áreas do nosso encéfalo e na reverberação das informações durante o sono que foram obtidas durante o estado de vigília (Ribeiro & Nicolelis 2004). O sono MRO possui um papel especial no que concerne a processos de neuroplasticidade e bioquímicos, como a ativação de genes imediatos, por exemplo, o gene zif-268 que possivelmente está relacionado com o tráfego de informação da passagem de memória do hipocampo para o córtex cerebral (Ribeiro et al 2002).
A partir da década de 1970 trabalhos foram publicados fazendo uma associação com tarefas cognitivas e o aumento específico de sono MRO. Os trabalhos de Lucero (1970), Fishbein et al (1974) e Smith (1982) demonstrou que tarefas cognitivas como esquiva discriminativa e exploração de labirinto em busca de recompensa aumentaram especificamente os episódios de sono MRO. O trabalho de Schiffelholz e Aldenhoff (2002) demonstrou que a tarefa de exploração de objetos aumenta o numero de episódios de sono MRO. O presente trabalho está de acordo com os resultados encontrados na literatura, verificamos em nosso paradigma que a duração de sono MRO aumenta significativamente nas quatro primeiras horas de registro após a exploração de objetos.