Superação de dormência de sementes como estratégia para restauração florestal de pastagem tropical.
Texto
Imagem
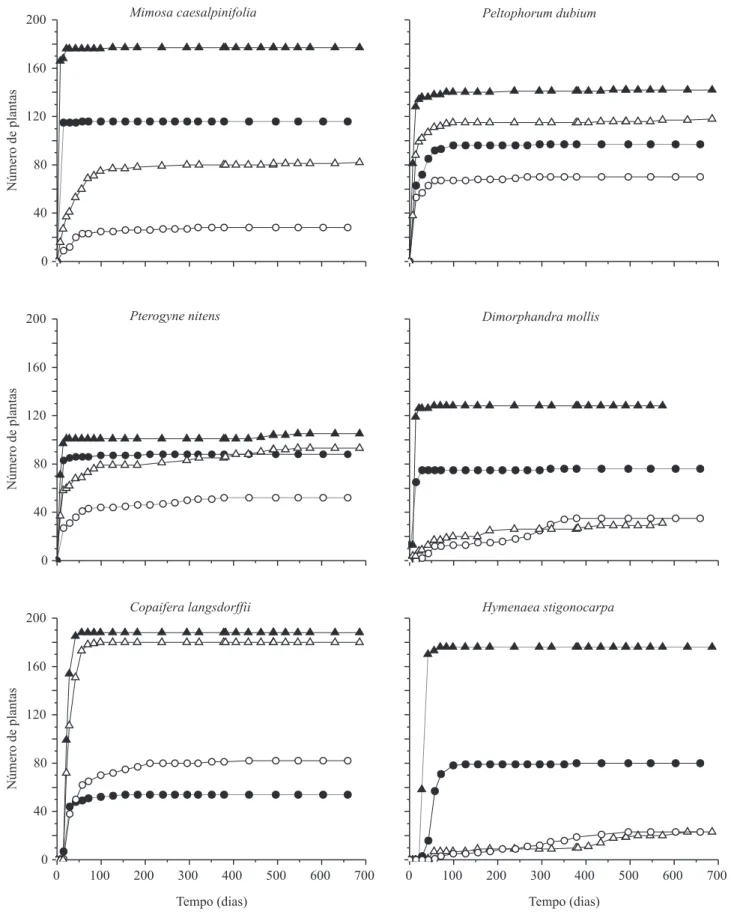
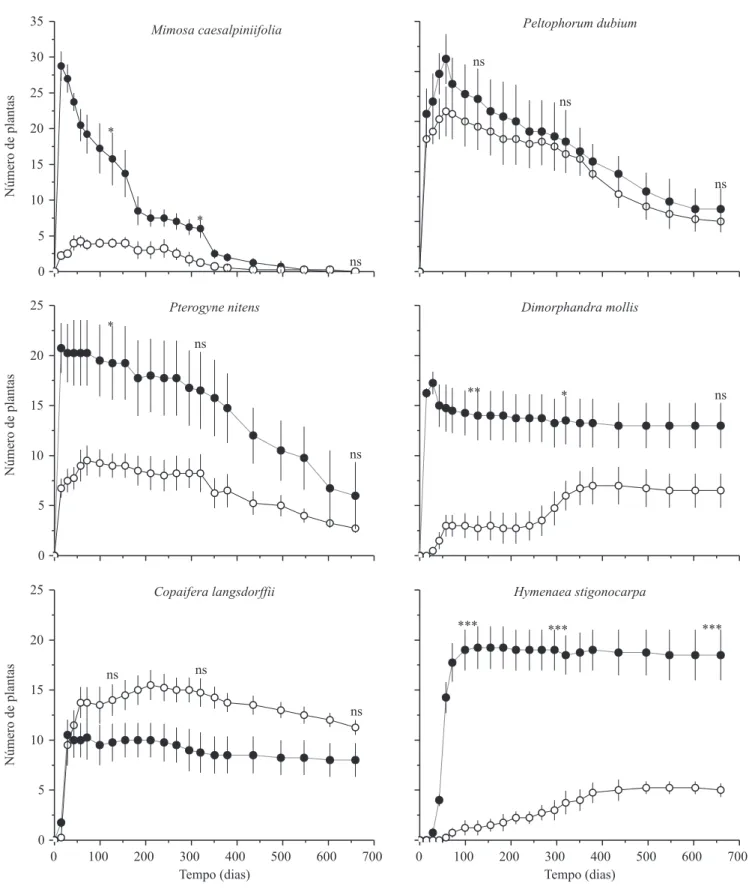
Documentos relacionados
O objetivo do curso foi oportunizar aos participantes, um contato direto com as plantas nativas do Cerrado para identificação de espécies com potencial
O abate ocorreu quando os animais atingiram peso médio de 325 kg (±20), entre 28 e 30 meses de idade, sendo avaliados os parâme- tros comprimento de carcaça fria, peso dos
é bastante restrita, visto que tanto suas duas entradas, quanto as galerias e condutos que interligam os pequenos salões são bastante estreitos, e a umidade na maioria dos salões
Internal sac armature (Fig. 5) consisting of two basal spine-shaped straight sclerites, 2.8 times as long as wide (Fig. 5a) or as in figure 5A; two long, laminar and median
var sparcialE, sparcialD: integer; procedure Soma; var n: integer; begin read(n); if n <> 0 then begin sparcialE := sparcialE + n; write(sparcialE, ’ ’); Soma; sparcialD
Tabela 1 - Germinação (%), índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG), em dias, de sementes de Rheedia brasiliensis, submetidas a
a) AHP Priority Calculator: disponível de forma gratuita na web no endereço https://bpmsg.com/ahp/ahp-calc.php. Será utilizado para os cálculos do método AHP
