Luciane Dutra Coletti
A influência do status social no forrageio de itens camuflados e avermelhados em
sagüis (Callithrix jacchus) cativos
Dissertação apresentada à
Universidade Federal do Rio Grande do
Norte, como requisito à obtenção do
título de mestre no Programa de
Pós-Graduação em Psicobiologia.
Natal
Luciane Dutra Coletti
A influência do status social no forrageio de itens camuflados e avermelhados em
sagüis (Callithrix jacchus) cativos
Dissertação apresentada à
Universidade Federal do Rio Grande do
Norte, como requisito à obtenção do
título de mestre no Programa de
Pós-Graduação em Psicobiologia
Orientador: Prof. Daniel Marques de
Almeida Pessoa.
Natal
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências Coletti, Luciane Dutra.
A influência do status social no forrageio de itens camuflados e avermelhados em sagüis (Callithrix jacchus) cativos / Luciane Dutra Coletti. – Natal, RN, 2010.
78 f. : Il.
Orientador: Daniel Marques de Almeida Pessoa.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Callithrix jacchus – Dissertação. 2. Visão de cores – Dissertação. 3. Sagüi – Dissertação. I. Pessoa, Daniel Marques de Almeida. II. Universidade Federal do Rio Grande do Norte. III. Título.
Título: A influência do status social no forrageio de itens camuflados e avermelhados em
sagüis (Callithrix jacchus) cativos
Autor: Luciane Dutra Coletti
Data da defesa: 20/5/2010
Banca Examinadora:
___________________________________
Prof. Valdir Filgueiras Pessoa
Universidade de Brasília, DF
___________________________________
Profa. Maria Emília Yamamoto
Universidade Federal do Rio Grande do Norte, RN
___________________________________
Prof. Daniel Marques de Almeida Pessoa
RESUMO
Enxergar cores pode ser vantajoso, já que elas são um importante sinal por fornecer informações a respeito do ambiente, tal como a localização do alimento. No entanto, nem todos os animais enxergam estes sinais cromáticos da mesma forma. Nos primatas, o grupo dos Plathyrrhini possui visão polimórfica ligada ao sexo, com os machos sempre dicromatas e as fêmeas podendo ser dicromatas ou tricromatas. Estudos indicam que os tricromatas teriam vantagens no forrageio por enxergar melhor que os dicromatas os frutos maduros avermelhados contra o fundo verde de folhagens. Já os dicromatas distinguiriam melhor insetos camuflados do que os tricromatas. O sagüi (Callithrix jacchus) é uma espécie de
primata neotropical que possui visão de cores polimórfica. Esta espécie constitui grupos familiares com alto viés reprodutivo e as fêmeas reprodutoras geralmente têm acesso preferencial ao alimento. Neste trabalho verificou-se se este contexto social exerce influência na habilidade de forrageio de itens camuflados e avermelhados em grupos de C. jacchus.
ABSTRACT
Seeing colors can be advantageous, because they are an important signal for providing information about the environment, such as the location of food. However, not every animals sees these chromatic signals in the same way. In primates, the group of Plathyrrhini has polymorphic sex-linked vision with males always dichromats and dichromats or trichromats females. Studies indicate that trichromats during foraging would benefit by seeing better than dichromats ripe fruits against the green foliage background. On the other hand, dichromats appear to distinguish camouflaged insects better than trichromats. The marmoset (Callihtrix jacchus) is a neotropical primate species that have color vision polymorphism. This species
establishes family groups with highly reproductive bias, with breeding females often having preferential access to food. This work aims to study whether the social context influences the foraging ability of camouflaged and red items in groups of C. jacchus. Four groups of captive
marmosets were presented to four food tasks, involving difficult, easy, reddish and camouflaged food targets. Foods were presented in a concentrated and dispersed manner, to check whether there was monopolization of the resources by the dominant subjects and if this would affect the ability of individuals to find the food targets. Success was measured by latency to food acquisition and number of targets consumed. Males and females differed in their foraging success for camouflaged and reddish items, although this difference has not appeared in all situations and experimental conditions. In general males were more successful for detecting camouflaged items while females succeeded more in identifying reddish items. There were no differences in foraging success between individuals of different social status, however, there were differences in the success of consumption of food items for different situations when food was concentrate compared with dispersed food. Taken as a role, there was a greater difficulty in detecting food items when they were presented in concentrated arrangement, which is supposed to be related to a higher difficulty to approach and stay near the food. Although it appears that there was no direct competition seems to have group's indirect influence on the detection of food items and foraging success of individuals, affecting mainly those items more difficult to detect.
SUMÁRIO
1.Introdução _________________________________________________ 1
1.1 Objetivos ________________________________________________ 10
1.2 Hipóteses e predições ______________________________________ 10
2. Metodologia________________________________________________ 12
2.1 Sujeitos________________________________________________ 12
2.2 Procedimento experimental_________________________________ 13
2.3 Teste de preferência alimentar ______________________________ 21
2.4 Observação comportamental _______________________________ 23
2.5 Análise estatística________________________________________ 24
3. Resultados ________________________________________________ 26
3.1 Habituação e experimento piloto____________________________ 26
3.2 Visão de cores e sucesso no forrageio ________________________ 28
3.3 Preferência alimentar______________________________________ 39
3.4 Relações de dominância____________________________________ 41
3.5 Status social e sucesso no forrageio ___________________________ 45
4. Discussão___________________________________________________ 53
5. Conclusões __________________________________________________ 64
LISTA DE FIGURAS
Figura 1 - Desenho esquemático do recinto das famílias estudadas _____________ 14
Figura 2 - Detalhe do interior de um recinto evidenciando a janela que interliga os recintos com a divisória de madeira fechada_______________________ 14
Figura 3 - Foto do interior do recinto casa de uma família ____________________ 15
Figura 4 - Potes de plástico com os pedaços de madeira no seu interior _________ 17
Figura 5 - Desenho esquemático com a distribuição dos potes dentro do recinto na condição dispersa ___________________________________________ 19
Figura 6 - Desenho esquemático com a distribuição dos potes dentro do recinto na condição concentrada ________________________________________ 20
Figura 7 - Desenho esquemático da disposição dos potes dentro do recinto no teste de preferência_______________________________________________ 23
Figura 8 - Mediana do número de balas consumidas por faixa etária em cada sessão experimental do experimento piloto_____________________________ 27
Figura 9 - Mediana do tempo de latência (em segundos) para a aquisição da primeira bala por faixa etária___________________________________ 28
Figura 10 - Mediana do número de balas consumidas em cada sessão experimental da condição dispersa A) para machos e B) para fêmeas______________ 30
Figura 11 - Mediana do número de balas consumidas por machos e fêmeas em cada sessão experimental da condição dispersa_________________________ 31
Figura 12 - Mediana do tempo de latência na aquisição da primeira bala na condição dispersa A) para machos e B) para fêmeas________________________ 32
Figura 13 - Mediana do tempo de latência na aquisição da primeira bala por machos e fêmeas na condição dispersa__________________________________ 33
Figura 14 - Mediana do número de balas consumidas pelos machos em cada sessão experimental da condição concentrada A) para machos e B) para fêmeas 34
Figura 15 - Mediana do número de balas consumidas por sexo na condição concentrada________________________________________________ 35
Figura 17 - Mediana do tempo de latência para obtenção da primeira bala por sexo na condição concentrada______________________________________ 37
Figura 18 - Mediana do número de balas consumidas por sessão experimental A) para machos e B) para fêmeas__________________________________ 38
Figura 19 - Mediana do número de balas consumidas por indivíduo por sessão experimental no teste de preferência alimentar_____________________ 39
Figura 20 - Mediana do número de balas consumidas por sessão experimental no teste de preferência alimentar para A) machos e B) fêmeas___________ 40
Figura 21 - Representação das relações de dominância na Família F1____________ 41
Figura 22 - Representação das relações de dominância na Família F5____________ 42
Figura 23 - Representação das relações de dominância na Família F10___________ 43
Figura 24 - Representação das relações de dominância na Família F12___________ 44
Figura 25 - Mediana do número de balas consumidas por sessão experimental para um dos grupos de status social na condição de recurso disperso________ 46
Figura 26 - Mediana do número de balas consumidas por sessão experimental para um dos grupos de status social na condição de recurso concentrado____ 47
Figura 27 - Mediana do número de balas consumidas por sessão experimental para cada um dos grupos de status social nas diferentes condições A) casal
reprodutor B) filhos adultos e C) filhos juvenis____________________ 49
Figura 28 - Mediana do tempo de latência para cada um dos grupos de status social na condição de recurso disperso________________________________ 50
Figura 29 - Mediana do tempo de latência para cada um dos grupos de status social na condição de recurso concentrado_____________________________ 51
LISTA DE TABELAS
Tabela 1 - Perfil dos sujeitos experimentais ____________________________ 12
Tabela 2 - Situações experimentais ___________________________________ 17
Tabela 3 - Matriz das relações de dominância da família F1________________ 41
Tabela 4 - Matriz das relações de dominância da família F5________________ 42
Tabela 5 - Matriz das relações de dominância da família F10_______________ 42
AGRADECIMENTOS
Gostaria de agradecer inicialmente aos colegas do mestrado da psicobiologia que sempre foram um grande incentivo e também uma fonte de conhecimento e discussão constante.
Gostaria de agradecer pelas ajudas estatísticas ao Wall Hatori e ao Felipe Nalon, pela disponibilidade e grande esclarecimento.
À CAPES pela concessão da bolsa de mestrado que foi de fundamental importância para a permanência no mestrado e desenvolvimento do trabalho.
À Docile, na pessoa de Nestor Heineck, por gentilmente fornecerem todas as balas de goma utilizadas neste trabalho.
À Profa. Renata Ferreira pela ajuda e esclarecimento referentes ao DOMINA.
À Profa. Fívia Lopes pelo auxílio durante todo o desenvolvimento das observações comportamentais assim como pelos esclarecimentos na parte de análise social do trabalho.
Ao Tiago Eugênio (Kutako) não só pela ajuda estatística, mas pela ajuda na fase de delineamento, e de atordoamento e pelo apoio durante todo o desenvolvimento do projeto.
Ao Ricardo Emídio (Cabrunco) pelas longas discussões primatológicas, em especial aquelas no Tomaz, e pelo apoio e ajuda constantes.
Às colegas de trabalho e de núcleo Daniela Menezes e Alessandra Santiago, pelas colaborações tanto no núcleo como na coleta de dados, em especial à Alessandra que foi de crucial ajuda no final do desenvolvimento deste trabalho.
Aos colegas de casa Tiago Cabral (Jordy) e Frederico Horie (Zaca) por tornarem sempre boa a volta casa e pelos almoços, claro.
Aos funcionários do Núcleo de Primatologia da UFRN, que sempre foram muito prestativos e atenciosos durante toda a execução do projeto no núcleo.
À minha família e aos meus amigos do Rio Grande do Sul, que apesar de estarem tão distantes sempre estiveram ao meu lado.
E por último aos sagüis, por terem sido não só o objeto de estudo, mas por terem tornado o trabalho algo muito mais interessante e muito mais agradável, sendo por vezes o maior incentivo a continuar.
Muito obrigada!
1 1. INTRODUÇÃO
As cores são importantes sinais para os animais, pois permitem segmentar áreas de
coloração diferente e unificar áreas de mesma coloração como pertencendo a um mesmo
objeto (Derrington et. al., 2002). As células que geralmente estão envolvidas na visão de cores
para a maioria dos vertebrados são os cones (Kelber et. al., 2003). A luz oriunda do ambiente
é captada por estas células na retina, pelos seus fotopigmentos, através do retinal ligado a uma
proteína específica chamada opsina. O retinal absorve preferencialmente comprimentos de luz
específicos, dependendo da constituição de aminoácidos da opsina.
Cada fóton de luz de comprimento de onda específico tem uma probabilidade diferente
de ser absorvido por cada um dos tipos de cones, sendo esta maior quanto mais aproximado
estiver do ótimo de absorção do cone. Quando um animal possui dois cones com
fotopigmentos que absorvem preferencialmente diferentes comprimentos de onda, o sistema
nervoso consegue contrastar estes dois sinais. Através deste contraste é possível compreender
exatamente que tipo de composição espectral o objeto que está sendo visualizado está
emitindo (Gegenfurtner & Kiper, 2003). Quanto maior o número de fotopigmentos que um
animal possui, melhor irá ser sua visão de cores, pois este irá possuir um maior número de
canais de oponência e irá também absorver uma faixa maior de comprimentos de onda
(Kelber et. al., 2003).
Acredita-se que o ancestral amniota possuía uma visão de cores tetracromática, ou
seja, possuía quatro tipos de fotopigmentos, e que no decorrer da evolução os primeiros
mamíferos, que seriam possivelmente noturnos, teriam perdido dois deles, ficando com dois
tipos de fotopigmentos como uma adaptação à vida noturna (Jacobs, 1993; Kelber et. al.
2
De todos os mamíferos eutérios os únicos que apresentam visão de cores tricromática,
ou seja, que possuem além do cone S (short), dois outros tipos de cones, são os primatas
(Dominy et. al. 2003). Dentro da ordem Primates todos os animais parecem ter um cone S
cujo pico de absorção ( max) fica em torno de 433 nm, e o gene que codifica este pigmento é
autossômico. Já os cones de absorção de ondas mais longas (M/L de middle e long),
localizam-se no cromossomo sexual X, existindo grandes variações dentro dos primatas
quanto a seus picos de absorção (Dulai et. al., 1999). Supõe-se que a visão de cores entre os
primatas teria se originado de um ancestral comum que seria dicromata e que a visão
tricromata teria se originado posteriormente, no entanto, esta origem ainda está sendo
discutida (Jacobs, 2007).
Os primatas do Velho Mundo (Catarrhini) são os únicos a apresentar tricromacia
uniforme, ou seja, todos os indivíduos são tricromatas. Isto foi possível devido a uma
duplicação do gene que codifica o fotopigmento longo (Jacobs, 2007). Os prossímios
apresentam linhagens possivelmente monocromatas (algumas espécies noturnas) e duas das
maiores linhagens diurnas, que se saiba até o momento, com polimorfismo ligado ao sexo, da
mesma forma que muitos primatas do Novo Mundo (Tan & Li, 1999; Leonhardt et. al., 2008).
Os Platyrrhini, primatas do Novo Mundo, que habitam toda região Neotropical,
possuem visão de cores polimórfica e ligada ao sexo. Ou seja, eles possuem, assim como os
Catharrini, um gene autossômico para a opsina S, porém, possuem apenas um gene que
codifica os pigmentos dos cones longos M/L situado no cromossomo sexual X, havendo
apenas um lócus para três ou mais versões alélicas diferentes deste gene (Dulai et. al. 1999;
Jacobs, 2007). Com isso os machos serão sempre dicromatas, bem como as fêmeas
homozigotas. Já as fêmeas heterozigotas serão tricromatas, devido ao mecanismo de
3
Existem, até onde se sabe no momento, duas exceções a este padrão. Uma delas são os
bugios, gênero Alouatta (Jacobs et al., 1996). Neste gênero houve uma duplicação do gene
codificador do fotopigmento longo M/L, independente e mais recente que aquela ocorrida nos
Catharrini e que ocorreu possivelmente depois da divergência dos dois grupos (Kainz et. al.,
1998). Com esta duplicação se torna possível tanto para machos quanto para fêmeas
possuírem dois tipos de cones longos, e, portanto, visão tricromática de cores (Araújo et. al.,
2008). Este é o único gênero de Platyrrhini a possuir tricromacia uniforme na sua população
(Jacobs, 1998). Outra exceção ao polimorfismo visual é o gênero Aotus. Trabalhos
verificaram através de exames eletrofisiológicos e psicofísicos que este primata noturno é
monocromata, tendo sido identificado apenas um único tipo de cone M/L com max 543 nm
(Jacobs, 1993). Sendo assim, ele não possui percepção de cores. Esta parece ser uma condição
secundária oriunda de mutações deletérias em genes codificadores de opsinas e discute-se que
pode representar uma adaptação à vida noturna (Jacobs et al., 1993; Jacobs, 1998).
Os calitriquídeos, assim como a maioria dos Plathyrrhini, possuem polimorfismo na
visão de cores ligado ao sexo e, até onde se sabe no momento, todos possuem em comum três
picos de absorção de pigmentos longos ( máx 543, 556 e 562 nm) que variam ligeiramente dos
picos encontrados para espécies de outras famílias de Platyrrhine (Tovée et al., 1992; Jacobs,
1998; Surridge & Mundy, 2002; Jacobs, 2007). Em Callithrix penicillata estudos
comportamentais demonstraram polimorfismo na visão de cores (Pessoa et al., 2005). Para a
espécie do presente estudo, o sagüi-comum (Callithrix jaccus), estudos genéticos (Williams
et. al., 1992; Hunt et. al., 1993) microespectrofotometricos e comportamentais (Tovée et. al.,
1992; Derrington et. al., 2002) comprovaram que C. jacchus apresenta distinção de cores, e,
como os outros calitriquídeos, polimorfismo na visão de cores ligado ao sexo. Segundo estes
4
espera-se para sua população uma proporção de dois terços de fêmeas tricromatas e um terço
delas dicromatas (Surridge & Mundy, 2002; Jacobs, 2007)
Acredita-se que exista uma explicação adaptativa para a manutenção deste padrão
polimórfico encontrado na visão dos primatas do Novo Mundo, já que ele tem se mantido por
aproximadamente 13 milhões de anos (Surridge & Mundy, 2002; Surridge et. al., 2003;
Jacobs, 2007).
A hipótese mais difundida para explicar esta manutenção é de que existiria uma
vantagem no forrageio para os tricromatas na detecção de frutos maduros e de folhas jovens,
já que estes fenótipos visualizam melhor que os dicromatas as tonalidades avermelhadas
destes itens (Sumner e Mollon, 2000a; Sumner e Mollon, 2000b; Caine & Mundy, 2000;
Caine et. al., 2003; Smith et. al., 2003). Como esta tricromacia só existe na condição de
heterozigose (para fêmeas), haveria vantagem para os fenótipos heterozigotos, mantendo
assim a variedade de alelos dentro das populações, e, assim, o polimorfismo (Surridge et. al.,
2003; Melin et. al., 2007). Esta hipótese foi parcialmente corroborada por experimentos de
cativeiro que verificaram vantagem na detecção de alvos avermelhados, que simulavam frutos
maduros, pelos tricromatas em relação aos dicromatas (Caine & Mundy, 2000; Caine et. al.
2003; Smith et. al. 2003). Outros trabalhos, através de medidas de coloração de alvos naturais,
verificaram que os pigmentos dos Catharrinos são quase ótimos para distinguir tanto folhas
jovens quanto frutas maduras, do fundo de folhas maduras, sendo que as últimas são ainda
melhor distinguidas que as primeiras (Sumner e Mollon, 2000ª; Dominy & Lucas, 2001). No
entanto, ao tentar verificar se os mesmos fotopigmentos eram otimizados para a diferenciação
de frutos maduros de frutos verdes, ou seja, para melhor distinguir o grau de maturidade do
fruto, os mesmos autores do primeiro trabalho não encontraram resultado positivo (Sumner e
5
não-maduros, o canal M/L dos Catharrinos ainda assim traria vantagem na detecção dos frutos
maduros em relação ao canal ancestral para a maioria dos frutos analisados neste estudo.
Mesmo havendo evidências que corroborem a hipótese de vantagem na detecção de
folhas e frutos maduros, ainda existem questões em aberto em relação a ela. Uma delas é o
fato de existirem espécies de Plathyrrhini que apresentam polimorfismo na visão de cores que
não são folívoras (Surridge & Mundy, 2002) ou que se alimentam de variados frutos não
maduros ou crípticos (Perini et. al., 2009). Soma-se a isso existirem outras espécies cuja base
da dieta é o exsudato de árvores, como no caso de diversos calitriquídeos (Smith, 2000; Vilela
& Faria, 2002). Por causa destas diferenças na ecologia alimentar destas espécies é possível
que existam outras pressões seletivas que estariam mantendo este polimorfismo nas
populações dos macacos do Novo Mundo (Melin et. al. 2008).
Uma hipótese alternativa seria de que existiriam também vantagens de detecção para
os indivíduos dicromatas, o que poderia sugerir uma seleção dependente de freqüência, ou,
seja, que indivíduos com diferentes tipos de visão de cores teriam vantagens no forrageio de
itens alimentares distintos (Surridge et. al., 2003; Buchanan-Smith, 2005; Melin et. al., 2008).
Relativo a esta hipótese trabalhos recentes em cativeiro verificaram vantagem dos dicromatas
em relação aos tricromatas no que concerne à distinção de padrões camuflados (Saito et. al.,
2005; Caine et. al., 2003; Leonhardt et. al., 2008) que se daria através da quebra de padrões
cromáticos que dificultam a percepção de formas, tornando mais fácil a visualização dos
contornos dos objetos. Isso se refletiria em uma melhor detecção de frutos crípticos e de
insetos camuflados, trazendo vantagens no forrageio destes itens para os fenótipos dicromatas,
e, portanto, em uma vantagem seletiva (Melin et. al., 2008). Estudo de campo comprovou que
existe uma melhor detecção de insetos camuflados por primatas dicromatas em grupos livres
6
Apesar de muitos trabalhos terem recentemente se proposto a compreender melhor a
manutenção do polimorfismo na visão de cores dos primatas do Novo Mundo a questão ainda
continua em aberto. Isto se evidencia principalmente em trabalhos de campo recentes que não
conseguiram verificar uma vantagem significativa no forrageio tanto para indivíduos
tricromatas quanto para dicromatas em grupos de vida livre. Vogel et. al. (2006) não
encontrou vantagem no forrageio pelos indivíduos tricromatas em um grupo selvagem de
Cebus capucinus, assim como Dominy et. al. (2003), estudando grupos de Saguinus imperator imperator e S. fuscicolis wedelli. Já, Smith et. al. (2005) não encontrou diferenças
entre o tempo despendido em forrageio entre diferentes fenótipos de visão de cores em grupos
mistos de Saguinus fuscicolis e S. mystax. Da mesmo forma, Melin et. al. (2007) estudando
grupos livres de Cebus capucinus, aonde apesar de dicromatas conseguirem detectar melhor
insetos camuflados eles não despendiam mais tempo nem obtinham sucesso maior nesta
atividade quando comparados aos indivíduos tricromatas.
A espécie do presente trabalho, Callithri. jacchus, como é característico nos
calitriquídeos, é uma espécie onívora, tendo como base de sua dieta o exsudado de árvores
(Scanlon et. al. 1989; Power & Oftedal, 1996). A importância da goma na sua dieta varia
relacionada tanto ao hábitat quanto à estação do ano, podendo variar de 20 a 70% da sua dieta
(Power, 1996). A taxa de consumo de goma parece estar relacionada à disponibilidade dos
outros itens alimentares, como os frutos, sendo que quando estes ficam mais escassos a
ingestão de goma aumenta (Castro & Araújo, 2007). Os frutos são geralmente o segundo item
mais consumido pela espécie, havendo flutuações na sua taxa de consumo ao longo do ano
(Dias, 2007; Castro & Araújo, 2007). As espécies vegetais consumidas variam muito,
podendo inclusive conter em sua dieta espécies exóticas (Castro & Araújo, 2007; Dias, 2007).
Em menor porcentagem há o consumo de itens de origem animal, dentro dos quais se destaca
7
dentro de ocos. Estes itens, apesar de serem menos representativos na sua dieta, são essenciais
por ser a fonte prioritária de proteínas em sua dieta (Sussman & Kinzey, 1984).
O sagüi (Callithrix jacchus), assim como outros calitriquídeos, possui uma estrutura
social muito peculiar, sendo os grupos em sua maioria constituídos de indivíduos aparentados,
com alta interação social e com cuidado cooperativo da prole (Digby, 1995a; Mills et. al.,
2004). Em geral os grupos são monogâmicos com uma só fêmea reprodutora, porém existem
grupos com mais de uma fêmea reproduzindo (Digby, 1995b; Sousa et. al., 2005; Yamamoto
et. al., 2009). No entanto, grupos com múltiplas reprodutoras não são comuns (Sousa et. al.,
2005), pois a fêmea dominante faz a supressão da reprodução das outras fêmeas do grupo
através de supressão da ovulação, e também ativamente através de agressões direcionadas e
que podem incluir infanticídio de filhotes alheios (Digby, 1995b; Albuquerque et. al., 2001).
No caso de existir mais de uma fêmea reproduzindo no grupo isso parece ser reflexo de não
haver uma dominância bem estabelecida entre as fêmeas reprodutoras (Saltzman et. al., 2004;
Alencar et. al., 2006; Yamamoto et. al., 2009).
Os sagüis apresentam cuidado cooperativo da prole, onde não só o pai, mas também os
animais adultos e subaldutos do grupo, ajudam no transporte, na supervisão e na transferência
de alimentos para os filhotes (Locke-Haydon, 1984; Arruda et. al. 1986; Yamamoto & Box,
1996; Ximenes & Sousa, 1996; Mills et. al., 2004). Este cuidado é mais representativo nas
primeiras semanas de vida dos infantes diminuindo com o crescimento dos filhotes
(Locke-Haydon, 1984; Yamamoto & Box, 1996; Mills et. al., 2004). Apesar dos pais serem os
maiores cuidadores, principalmente o provável pai, o cuidado cooperativo dentro do grupo
diminui o gasto energético total do casal com a reprodução (Ximenes & Sousa, 1996; Hattori,
2004). Este é um comportamento característico da espécie, tendo sido observado tanto em
campo como em cativeiro, sem sofrer grandes modificações nos diferentes ambientes, ao
8
Em Callithrix jacchus verificou-se que existe acesso diferenciado ao recurso alimentar
pelos indivíduos do grupo relacionado a fatores tal como sexo e status reprodutivo, com as
fêmeas tendo acesso preferencial ao alimento (Box, 1997; Lopes et. al., 1997; Box et. al.,
1999). Em trabalhos de cativeiro foi verificada esta preferência no acesso da fêmea, assim
como um maior consumo de itens alimentares, que fica mais evidente em certas fases
reprodutivas, como gravidez e logo após o parto. Um dos fatores que parece determinar a
aquisição e manutenção deste acesso preferencial é a maior agressividade das fêmeas
reprodutivas em relação aos demais indivíduos, tanto entre o casal como dentro do grupo
(Lopes et. al., 1997; Michels, 1998). Já em grupos de sagüis em que a fêmea não está grávida
ou com prole dependente não existe um acesso preferencial entre o casal reprodutor (Lopes et.
al., 1997). É sugerido então que este acesso preferencial da fêmea ocorra devido a uma maior
demanda energética da fêmea nestes períodos (Box, 1997; Lopes et. al., 1997; Box et. al.,
1999; Tardif et. al., 2001; Tardif et. al., 2005) que influenciaria tanto a agressividade da fêmea
como a permissividade dos outros indivíduos do grupo (Box, 1997; Michels, 1998). No
entanto, Yamamoto et. al. (2004) encontrou diferenças na solução de uma tarefa alimentar
entre os gêneros, entre casais não reprodutivos, com as fêmeas resolvendo-a mais facilmente.
Box et. al. (1999) também sugere que as fêmeas sejam mais responsivas a tarefas alimentares
do que os machos. Isto leva a crer que podem haver outros fatores além da necessidade
energética influindo no acesso preferencial das fêmeas, como uma maior responsividade das
mesmas.
Estudando um grupo de vida livre, Dias (2007) verificou um padrão semelhante. A
fêmea dominante (reprodutora) sempre tinha acesso preferencial ao alimento e na maioria das
vezes não permitia a aproximação de outros indivíduos enquanto se alimentava. No entanto,
isto se modificava com o tipo de alimento, sendo mais restritiva em relação a alimentos de
9
relação aos alimentos de origem animal, quando um animal subordinado conseguia capturar
uma presa, era freqüente o roubo por parte dos animais dominantes. Ele viu ainda que quando
o alimento era espalhado e abundante a tendência era não haver competição por este, com os
indivíduos se distribuindo por toda área. Já quando o alimento era concentrado, por exemplo,
somente um ou dois frutos em uma determinada árvore, a agressão se tornava freqüente
dentro do grupo.
Tendo em vista que a alimentação natural de Callithrix jacchus se constitui tanto de
frutas maduras que devem ser apanhadas nas árvores como de insetos que devem ser
visualizados tanto no solo como nos troncos das árvores espera-se que a visão de cores tenha
influência no sucesso de forrageio dos indivíduos. No entanto, num contexto de grupo
existem outros fatores que podem estar influenciando esta detecção. Trabalhos como o de
Caine (2000) e Caine et. al. (2003), que visaram avaliar as vantagens na detecção de alvos
alimentares por dicromatas e tricromatas, utilizou o grupo todo na realização da tarefa
alimentar. No entanto, este trabalho, apesar de ser executado em meio a um ambiente social,
não levou a variável da hierarquia social dos indivíduos em consideração. Porém, pelas
interações sociais serem tão importantes no sucesso de forrageio em C. jacchus é de se
esperar que os animais subordinados, ainda que detectando visualmente mais rapidamente os
alvos (alimentos), não consigam sucesso alimentar maior que o de animais de seu grupo de
hierarquia social mais elevada. Tanto pode ser restringido seu acesso ao alimento, quanto,
ainda que detectado e apanhado antes, este venha a ser roubado por outros membros do grupo
de status superior, como já encontrado em outros trabalhos (Box, 1997; Lopes et. al., 1997;
Michels, 1998; Box et. al., 1999; Dias, 2007). Em vista de que a fêmea reprodutora, na maior
parte das vezes, é o indivíduo com maior status dentro do grupo, é esperado que esta obtenha
maior sucesso, devido a seu acesso preferencial aos alimentos (Box, 1997; Lopes et. al., 1997;
10
Dentro deste contexto, pretende-se com este trabalho testar o sucesso de forrageio de
animais de ambos os sexos e de distintas hierarquias sociais quanto a sua detecção dos alvos,
e eficiência na aquisição do alimento, e, assim, compreender melhor como as interações
sociais podem influenciar na sua habilidade de forrageio.
1.1 Objetivos
Objetivo geral:
Verificar se o contexto social exerce influência na habilidade de forrageio de itens
crípticos e conspícuos em machos e fêmeas de Callithrix jacchus cativos.
Objetivos específicos:
- Determinar se em grupos de sagüis cativos a hierarquia social se correlaciona ao
desempenho em tarefas de forrageio.
- Investigar se machos e fêmeas diferem com relação aos seus desempenhos em tarefas
de detecção de itens alimentares avermelhados e camuflados.
- Analisar se a defensibilidade do recurso modifica o desempenho dos sagüis de
diferentes hierarquias sociais na tarefa apresentada.
1.2 Hipóteses e predições
11
O status social do indivíduo irá influenciar o seu acesso ao alimento, e por
conseqüência, o seu sucesso de forrageio nas diferentes tarefas apresentadas.
Predições:
1: Quando o alimento for defensável haverá a monopolização deste pelos indivíduos
dominantes que se refletirá na identificação de um número maior de itens alimentares e/ou
pela menor latência média no tempo de identificação.
2: Quando o alimento não for defensável sua monopolização por indivíduos
dominantes será dificultada e haverá uma maior paridade no desempenho de indivíduos do
grupo de diferentes status sociais.
Hipótese 2:
Machos e fêmeas por possuírem diferentes visões de cores, na maioria dos casos,
diferem no sucesso de forrageio de itens avermelhados e camuflados.
Predições:
1: Machos obterão um maior número de itens alimentares camuflados e/ou menor
latência na aquisição, do que as fêmeas.
2: Fêmeas obterão um número maior de itens avermelhados e/ou menor latência na
12 2. METODOLOGIA
2.1 Sujeitos
Os sujeitos experimentais foram vinte e dois sagüis (Callithrix jacchus) residentes do
Núcleo de Primatologia da Universidade Federal do Rio Grande do Norte. Estes vivem em
unidades familiares já estabelecidas, sendo utilizadas neste trabalho quatro famílias,
estruturadas como segue:
Tabela 1: Perfil dos sujeitos experimentais.
Família Indivíduo Sexo† Faixa etária*
F 12 815 – Otávio M Adulto
918 – Filó F Adulto
1071 – Firmino M Adulto
1048 – Flip F Adulto
1087 – Felinto F Adulto
1105 – Flocão M Juvenil
1070 - Fátima F Juvenil
F 1 993 – Osvaldo M Adulto
998 – Joana F Adulto
1026 – Jurema F Adulto
1107 – Jerônimo M Juvenil
1109 – Jordão M Juvenil
F 10 1015 – Bekaya M Adulto
1030 – Florize F Adulto
1099 – Fernando M Subadulto
1111 – Otelo M Juvenil
Oberon M Infante
Odrade F Infante
F 05 982 - Olganita F Adulto
785 - Deribaldo M Adulto
13
11013 - Opus M Juvenil
* Segundo Yamamoto (1993); † M machos e F Fêmeas.
Não se realizou a análise genotípica da visão de cores dos animais de estudo, portanto,
espera-se que todos os animais do sexo masculino sejam dicromatas e que para as fêmeas
exista uma proporção de um terço de fêmeas dicromatas e dois terços de fêmeas tricromatas
(Surridge & Mundy, 2002; Jacobs 2007)
As fêmeas reprodutoras dos grupos estavam em diferentes fases reprodutivas,
incluindo gravidez e lactação, já que não houve restrição reprodutiva nas famílias estudadas.
Pelo mesmo motivo todas as famílias possuíam animais em variadas faixas etárias, incluindo
animais imaturos, como juvenis e/ou infantes. Durante a execução do experimento o número
de indivíduos nas famílias se alterou devido ao nascimento de filhotes.
Para uma maior motivação dos animais em realizar as tarefas eles eram deixados em
privação alimentar até o término da sessão experimental, atrasando-se a alimentação matinal
de 30min a até 1h30min, dependendo da ordem de experimentação da família, descrita
posteriormente. A água foi sempre ad libitum.
2.2 Procedimento experimental
O experimento foi realizado nos próprios recintos das famílias. Estes são cativeiros de
2x2x2m, e possuem em seu interior prateleiras de diferentes alturas em duas de suas paredes e
três poleiros de diferentes alturas interconectando as prateleiras, bem como a caixa ninho (Fig.
1). Adjacente a este, existe um recinto de 1x1x2m, interligado através de uma abertura (Fig.
2), utilizado exclusivamente para o manejo dos animais (Fig. 1). Ambos os recintos possuem
paredes de alvenaria em dois lados opostos, além de um visor unidirecional e uma porta de
14
contato sonoro e olfativo com outras famílias residentes em recintos vizinhos. Os recintos são
apenas parcialmente cobertos, sendo desta forma iluminados por luz natural (Fig.3).
Figura 1. Desenho esquemático do recinto das famílias estudadas. A) prateleira contendo os pratos da alimentação regular das famílias; B) localização da divisória entre os recintos; C) prateleiras do recinto, com diferentes alturas; D) poleiros do recinto, em diferentes alturas.
Figura 2: Detalhe do interior de um recinto evidenciando a janela que interliga os recintos com a divisória de madeira fechada.
Recinto
Adjacente Recinto Casa
A B
D C
D
C
15 Figura 3. Foto do interior do recinto casa de uma família. À frente vidro unidirecional, ao fundo parede de grades, no canto superior à esquerda a caixa ninho, e, em diferentes alturas, os poleiros.
Antes de iniciar qualquer atividade com os animais foram realizadas 63hs de
habituação dos sagüis com os dois experimentadores, através da observação por 30min de
cada uma das famílias do núcleo, durante cinco dias da semana, por cinco semanas seguidas.
A habituação inicial dos animais com os experimentadores visou minimizar a influência da
presença e da voz dos experimentadores no comportamento dos animais durante as sessões
experimentais subseqüentes.
A fim de condicionar os animais a que se dirigissem ao recinto adjacente antes de cada
sessão experimental foram feitas sessões de treino. Este condicionamento foi feito
recompensando-se os animais com pequenos pedaços de banana assim que estes entravam
neste recinto. Tomou-se cuidado para que todos fossem igualmente recompensados e para que
16
dos animais. Este condicionamento foi importante para que os sagüis se dirigissem por livre
vontade ao recinto adjacente, de forma a evitar estressá-los durante a execução do
experimento.
Concomitantemente, durante estas sessões de treino, foi feita a habituação dos animais
com o aparato experimental. Esta habituação foi importante para evitar que os animais não
interagissem com o aparato por medo ou por desconhecerem que havia nele recompensas.
Durante a habituação foram utilizados os mesmos alimentos alvo que seriam posteriormente
utilizados no experimento, sendo as balas de goma de coloração vermelha. Os animais eram
deixados em contato com o aparato até que a maioria do grupo tivesse entrado em contato
com este e encontrado as balas. Estas sessões foram conduzidas ao longo de 5 dias para se
certificar que todos os animais tivessem interagido com o aparato e tivessem conseguido obter
dele a recompensa (balas de goma) pelo menos em uma das sessões. Seguiu-se a habituação
dos animais com a execução do experimento piloto, durante dez dias. Este experimento piloto
inclui todas as etapas que seriam posteriormente realizadas durante o experimento em si,
porém com as balas de coloração roxa que eram apresentadas em contraste com o fundo de
madeiras de cor verde, condição considerada de fácil detecção para todos os fenótipos de
visão de cores, neste caso, tanto para machos como para fêmeas.
Depois de completada a habituação deu-se início ao experimento. Para o experimento
os animais inicialmente eram contidos no recinto adjacente durante o arranjo dos aparatos
experimentais no recinto casa. Esta contenção era realizada apenas durante o arranjo do
aparato. Depois disso todos os animais tinham acesso ao recinto casa através da janela
existente entre os compartimentos que era novamente aberta. Utilizou-se como alimento alvo
balas de goma (Ingredientes: xarope de glicose, água, amido de milho, aromatizantes e
corantes; marca Docile®) roxas, laranjas e/ou verdes. Como fundo foram utilizados cubos de
17
realizados ao longo da manhã, das 7h00min às 9h30min. A ordem de experimentação das
famílias deu-se através de sorteio, sendo feita de ordem aleatória, de forma a contemplar
todos os possíveis horários deste intervalo.
Foram utilizados 12 potes de plástico transparentes, de medidas 12x17cm, aonde
foram colocados sobre o fundo pedaços de madeira pintados. Sobre e entre os pedaços de
madeira foram colocadas as balas de goma, Fig. 4.
Figura 4: Potes de plástico com os pedaços de madeira no seu interior. Os círculos indicam as balas verdes entre os cubos de madeira verdes.
As famílias foram testadas em quatro situações diferentes, como segue na tabela 2:
Tabela 2: Situações experimentais.
Situação Fundo (Pedaços de madeira)
Alvo (Balas de goma)
Resultado esperado
1 Verde quadrado Verde quadrado Situação difícil para todos
2 Verde quadrado Roxo quadrado Situação fácil para todos
3 Verde quadrado Laranja quadrado Vantagem para
Tricromatas 4 Verde e laranja quadrado Laranja esférico Vantagem para
18
Para se certificar da sobreposição das colorações das balas verde/laranja com o fundo
verde elas foram testadas apresentando-se ambas a um humano com protanopia.
A situação 1 foi projetada para ser uma situação impossível, de difícil detecção tanto
para dicromatas quanto para tricromatas, afim de se verificar se os animais poderiam estar
detectando as balas através de algum outro tipo de sistema sensorial, ex. pelo odor.
A situação 2 com fundo verde quadrado e alvos de coloração roxa, é a situação
considerada fácil para ambos os fenótipos de visão de cores, sendo o alvo facilmente
diferenciado do fundo. Nessa situação todos os fenótipos deverão ter respostas semelhantes e,
portanto, serve como base de comparação para as outras situações experimentais.
A situação 3, fundo verde quadrado e alvo laranja quadrado foi projetada para que haja
uma vantagem de detecção pelos indivíduos tricromatas, já que estes poderão melhor
visualizar a localização dos alvos laranjas sobre o fundo verde, ao contrário dos indivíduos
dicromatas, para qual as balas e o fundo terão a mesma coloração (Caine & Mundy, 2000).
Desta forma espera-se que as fêmeas tenham vantagem na visualização destes alvos
alimentares.
A situação 4, com alvos esféricos laranja e fundo de coloração verde e laranja
quadrados, foi projetada para ser uma situação vantajosa para os indivíduos dicromatas já que
estes conseguem quebrar camuflagens mais facilmente do que tricromatas. Assim, com os
alvos laranja “camuflados” em um fundo bicolor (verde e laranja) a informação de forma
(alvo esférico sobre um fundo quadrado) será melhor visualizada pelos indivíduos dicromatas
que pelos tricromatas (Caine et. al., 2003). Assim, espera-se que os machos tenham vantagens
nesta situação experimental.
A ordem das situações foi também determinada de forma aleatória de modo a tentar
19
Estas diferentes situações foram apresentadas em duas condições diferentes. Na
primeira condição os potes foram apresentados separados uns dos outros, simulando um
recurso disperso. Existiam seis pontos do recinto com potes, organizados dois a dois, tanto
sobre bancos como sobre prateleiras, estas situadas a diferentes alturas, totalizando seis pares
de potes (12 potes ao todo), Fig. 5. Em cada um dos pares de potes apenas um deles continha
balas, dificultando a tarefa e estimulando a detecção prioritariamente visual dos alvos. Os
potes que continham balas dispunham de cinco balas cada um, totalizando também trinta
balas nesta condição experimental, Fig. 5.
Figura 5: Desenho esquemático com a distribuição dos potes dentro do recinto na condição dispersa. Círculos representam os bancos de madeira. Quadrados em cinza representam potes contendo balas; e os quadrados em branco, os potes sem recompensa.
Na segunda condição todos os 12 potes foram apresentados agrupados no centro do
recinto, estando em uma plataforma de madeira amparada por dois bancos, à 1m de altura,
Fig. 6. Dos 12 potes apresentados apenas dois deles continham balas, sendo 15 balas em cada
um, totalizando trinta balas. A disposição dos dois potes que continham a recompensa
20
animais quanto à disposição das balas no aparato experimental. Esta condição visa simular um
recurso concentrado, Fig. 6.
Figura 6: Desenho esquemático com a distribuição dos potes dentro do recinto na condição concentrada. Quadrados em cinza representam potes contendo balas; e os quadrados em branco, os potes sem recompensa.
Deste modo, as duas condições tinham uma área de procura igual (12 potes), uma
quantidade de recursos idêntica (30 balas) e diferiam apenas na área que o recurso ocupava.
Existiam apenas dois potes com recompensa localizados juntos na condição 1, ou seis potes
localizados em seis pontos diferentes do recinto na condição 2. Com isto, visou-se aumentar a
competição quando o recurso estivesse concentrado, já que os indivíduos dominantes podiam
mais facilmente dominar o recurso (Tardif & Richter, 1981; Box et. al., 1999; Dias, 2007).
Por outro lado, na condição dispersa, a defesa dos recursos não deveria ocorrer, já que a área
que o recurso ocupava era possivelmente grande demais para que os indivíduos dominantes
21
As duas condições foram apresentadas em blocos, sendo primeiramente apresentadas
as diferentes situações na condição espalhada, compreendendo 20 dias de execução; e
posteriormente na condição concentrada, com também 20 dias de execução do experimento.
Durante as sessões os experimentadores se posicionavam cada um de um lado da
gaiola para melhor visualização de todos os potes e animais do recinto, um posicionado do
lado do vidro unidirecional e outro no lado oposto, nas portas de grade. Cada um observava os
potes que estavam mais próximos de si. A posição dos observadores foi a mesma em todas as
famílias e em todos os dias, assim como cada observador registrava os potes de mesma
posição em todas as sessões experimentais. Eram registrados os tempos de latência gastos por
cada animal até encontrar cada item alimentar e também o número de balas encontradas por
cada indivíduo. Este registro foi feito com o auxílio de um gravador de voz. Os animais eram
deixados em contato com o aparato experimental por cinco minutos ou até acharem todas as
balas nos potes, o que ocorresse primeiro.
O número de balas por pote foi estipulado através de testes piloto com a maior família
aonde se verificou o número máximo de balas que a família conseguia consumir em cinco
minutos de contato com o aparato experimental.
2.3 Teste de Preferência Alimentar
As balas utilizadas no procedimento experimental já descrito, além de possuírem
colorações distintas (roxa, verde e laranja) possuem também sabores diferentes. A bala de cor
roxa possui sabor uva, a verde sabor limão e a laranja sabor laranja. Por causa disto, foi
realizado um teste de preferência alimentar para ver qual seria a influência do sabor da bala na
22
Este teste teve duração de dez dias e foi realizado entre as 07h30minh e 9h00min com
os animais ainda em jejum, com água ad libitum, sendo a alimentação matinal atrasada até o
término do teste. O teste foi realizado no próprio recinto das famílias, já descrito
anteriormente. Este teste foi realizado após o procedimento experimental descrito
previamente, sendo assim, os animais já haviam provado o sabor das balas.
Para a preparação do aparato os animais eram contidos no recinto adjacente da mesma
forma como descrito para o experimento anterior. Para o teste de preferência foram utilizados
seis potes plásticos transparentes idênticos aos já utilizados no procedimento experimental
(12x17cm), porém, os potes não continham nada em seu interior além das balas, a fim de
facilitar a visualização destas pelos animais. Seis potes foram distribuídos em dois pontos do
recinto, cada ponto contendo três potes, um localizado em uma prateleira e outro ponto sobre
um banco de madeira, a aproximadamente 1m e 1,2m do solo, em cantos opostos do recinto,
Fig. 7. Em cada ponto havia um pote contendo balas roxas, outro balas verdes e outro balas
laranja, de forma que todas as balas eram apresentadas concomitantemente em um mesmo
ponto. Cada pote possuía em seu interior seis pedaços de balas de goma, tendo estes pedaços
um quarto do tamanho de uma bala de goma. A ordem dos potes em cada um dos pontos era
23 Figura 7: Desenho esquemático da disposição dos potes dentro do recinto no teste de preferência. Círculo indica localização do banco; retângulos indicam localização dos potes, diferentes tons de cinza representam balas de colorações distintas no interior dos potes.
Cada sessão experimental tinha duração de três minutos, durante o qual era gravado,
com auxílio de gravador de voz, cada um dos pedaços de balas pegas pelos sagüis, qual sua
coloração e qual o animal que o pegou.
2.4 Observação comportamental
Os grupos também foram observados durante o turno da manhã, entre as 9h00min e
11h30min, aonde eram registrados comportamentos visando identificar o status social de cada
um dos indivíduos analisados.
Foi realizada a técnica de animal focal com dez minutos de observação para cada
animal do grupo, com registros instantâneos a cada 30 segundos. A ordem de observação foi
determinada através de sorteio antes do início das observações. Foram observados todos os
24
de subordinação foram registrados ad libitum durante cada animal focal. As observações
foram realizadas três dias por semana durante doze semanas.
Os comportamentos analisados foram os seguintes, baseado no descrito por Stevenson
& Poole (1976):
Comportamentos agonísticos
Direcionados:
a) Agressão: quando um animal morde, arranha, puxa ou bate em outro animal
b) Vocalizações: do tipo “erh-erh”.
c) Perseguições: quando um animal persegue outro que se afasta e se esquiva.
Comportamentos afiliativos
a) Proximidade e Contato físico com outro animal: quando o animal está contato direto
ou até 15 cm de distância de outro.
c) Catação: quando um animal inspeciona o pêlo de outro animal do grupo, abrindo os
pêlos com as mãos e mordiscando o pêlo.
d) Brincadeira: contém elementos como perseguições, rolamento no chão, saltos e
diferentes tipos de simulações de lutas.
Foram identificados os indivíduos que estavam envolvidos em cada um dos
comportamentos. Seguindo Lazaro-Perea et al. (2004), quando um episódio agonístico for
respondido por outro episódio agonístico foram registrados como duas interações agonísticas,
uma partindo de cada animal.
2.4 Análise estatística
Inicialmente foi feita uma análise preliminar para se certificar de que não havia
25
em forrageio, a experiência do indivíduo ao longo dos anos de vida, possam influenciar no
sucesso do indivíduo na tarefa (Leonhardt et. al. 2009). Por esta razão foi medido se os
indivíduos de diferentes faixas etárias obtêm sucesso diferenciado de acordo com suas idades.
Para estas análises foram realizados testes de Kruskal-Wallis utilizando-se como post hoc
testes de Mann-Withney.
Para testar a predição de que indivíduos de diferentes sexos terão um desempenho
diferente nas tarefas apresentadas foram utilizadas ANOVA de Friedman com post hoc de
Wilcoxon, para as análises dentro de cada um dos sexos, e, testes de Man-Withney para
comparar o desempenho nas tarefas entre os dois sexos.
Para testar a predição de que os indivíduos de status superior obterão um maior
sucesso no forrageio nas diferentes situações serão utilizados testes de Kruskal-Wallis, com
post hoc de Man-Withney . Em uma primeira análise as variáveis dependentes serão o sucesso
na obtenção de balas (número de balas encontradas) e em uma segunda será a latência para a
obtenção das mesmas. Estes testes foram realizados dentro de cada uma das condições
experimentais (agrupada ou espalhada), aonde se obteve quais dos grupos (dominantes ou
subordinados) obtiveram um maior sucesso em cada uma das situações em cada uma das
condições apresentadas. Se os indivíduos dominantes obtiverem um sucesso maior no
forrageio na condição concentrada, mas não na condição espalhada então as nossas predições
estarão confirmadas.
Para analisar se existe uma preferência dos indivíduos por um sabor específico de
balas utilizadas no experimento foram realizadas ANOVA de Friedman com post hoc de
Wilcoxon para se certificar se existia um consumo significativamente maior de algum sabor
pelos sagüis. Estes testes também foram realizados separadamente para cada um dos sexos
para se certificar se existem diferenças na preferência de machos e fêmeas.
26
A fim de identificar o status social de cada indivíduo dentro de sua família foram
construídas árvores de dominância com a utilização do programa DOMINA com os dados
coletados nas observações comportamentais das famílias deste trabalho de acordo com Izar et.
al. (2005). Estas árvores têm como vantagem a ilustração gráfica que facilita a visualização
das relações de dominância entre os membros de um mesmo grupo.
3. RESULTADOS
3.1 Habituação e Experimento Piloto
Ao final do experimento piloto, todos os animais já exerciam a tarefa de forma
satisfatória. Todos os animais interagiram com o aparato com alguns poucos animais não se
aproximando em alguns dias específicos. O único animal a não conseguir obter recompensas
do mesmo foi a fêmea reprodutora de uma das famílias (Filó, família 12), aparentemente
devido a um problema de locomoção da mesma, pois foi observado que ela se locomovia com
dificuldade pelo recinto, tanto antes quanto durante o experimento, o que a impossibilitava de
chegar até o aparato experimental e, assim, não obtendo recompensas. Pelo mesmo motivo
esta fêmea foi retirada das análises posteriores.
Precedendo todos os testes deste trabalho foi rodado um teste de
Kolmogorov-Smirnov aonde se verificou que os dados não apresentavam distribuição normal, desta forma
para todas as análises foram realizados testes não-paramétricos.
Com os dados obtidos durante o experimento piloto (balas de coloração roxa) foi
realizado o teste de Kruskal-Wallis aonde se comparou o sucesso na obtenção de balas dos
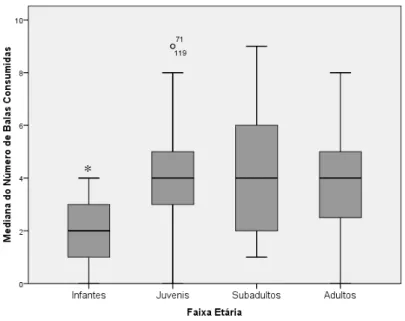
indivíduos de diferentes faixas etárias (infantes: 0-5 meses (n= 45); juvenis: 5-10 meses
27
realizada demonstrou haver pelo menos uma diferença significativa entre os grupos (H 3 =
38,1; p< 0,01). Testes de Mann-Whitney, com correção de Bonferroni, apontaram apenas um
grupo diferindo dos demais (U=671; p< 0,001), que se constitui dos indivíduos infantes (de 0
a 5 meses), Fig. 8.
Figura 8: Mediana do número de balas consumidas por faixa etária em cada sessão experimental do experimento piloto (barras indicam intervalo de 95% de confiança; ponto acima das barras representa os outliers).
O mesmo procedimento foi efetuado para comparar a latência na aquisição da primeira
bala entre as faixas etárias. Esta análise não indicou diferenças significativas neste quesito
(H3= 4,286; p>0,05), Fig. 9.
28 Figura 9: Mediana do tempo de latência (em segundos) para a aquisição da primeira bala por faixa etária (barras indicam intervalo de confiança de 95%; ponto acima das barras representa os outliers).
Levando-se em consideração os resultados acima apresentados, as análises seguintes
foram efetuadas sem os indivíduos infantes das famílias estudadas, sendo, daqui por diante,
vinte indivíduos experimentais de acordo com o apresentado na tabela 1.
3.2 Visão de cores e sucesso no forrageio
Para verificar a hipótese de que machos e fêmeas de Callithrix jacchus diferem na
identificação de itens crípticos e conspícuos foram realizados testes para averiguar se existem
diferenças significativas tanto na aquisição quanto na latência de aquisição dos itens
alimentares em cada uma das situações apresentadas sendo elas: Balas Verdes e
Fundo Verde; Balas Roxas e Fundo Verde; Balas Laranjas e Fundo Verde; Balas Laranjas
Redondas e Fundo Verde e Laranja. As duas condições (Recurso Disperso e Rccurso
Concentrado) foram testadas separadamente.
O tempo da sessão experimental foi de cinco minutos por família, como já descrito na
29
tempo máximo. Desta forma, foram contabilizadas as balas consumidas até que ainda
existissem balas disponíveis no aparato experimental, tendo sido então determinado o tempo
de três minutos como o tempo de corte, já que em nenhuma sessão as balas foram totalmente
consumidas antes deste tempo. Sendo assim, todos os resultados a serem apresentados se
baseiam em um tempo de interação com o aparato de 180 segundos.
Para a condição de recurso disperso inicialmente realizou-se testes ANOVA de
Friedman para comparar o sucesso na obtenção de balas e a latência na aquisição da primeira
bala nas quatro diferentes situações apresentadas tanto para os machos como para as fêmeas.
Houve diferenças significativas entre as diferentes situações tanto para machos ( 2 (3)=8,237;
p=0,041) quanto para fêmeas ( 2 (3)= 8,745; p=0,033) no sucesso de aquisição de balas.
Como análise post hoc realizou-se testes de Wilcoxon para ambos os sexos, com correção de
Bonferroni, sendo assim o nível de significância de 0,008. Para os machos as análises par a
par não apresentaram significância ao nível corrigido, (z>0,888; p>0,008), Fig. 10 A. Para
fêmeas somente duas situações diferiram significativamente, sendo elas a de Balas Roxas com
Fundo Verde e a de Bala Laranja Redonda e Fundo Verde e Laranja (z= 2,864; p=0,004).
Neste caso as fêmeas obtiveram uma quantidade maior de itens alimentares na situação de
Balas Roxas e Fundo Verde (Situação Fácil) do que na situação de Balas Laranjas Redondas e
Fundo Verde e Laranja (Situação Camuflada). Para todos os outros testes p>0,008, z> 0,884,
30 Figura 10: Mediana do número de balas consumidas em cada sessão experimental da condição dispersa A) para machos e B) para fêmeas (barras indicam intervalo de 95% de confiança; * p<0,008; pontos representam outliers).
Para averiguar as diferenças no sucesso de forrageio nas diferentes situações entre
machos e fêmeas foram realizados testes de Mann-Whitney para cada uma das situações. Não
houveram diferenças significativas entre as situações para machos e fêmeas nesta condição
experimental (U>811; p> 0,05), Fig 11. A)
31 Figura 11: Mediana do número de balas consumidas por machos e fêmeas em cada sessão experimental da condição dispersa (barras indicam intervalo de 95% de confiança; pontos representam outliers).
Seguido a isso, realizou-se os mesmos testes para averiguar o sucesso de aquisição de
itens alimentares através da latência para a aquisição das balas. Para os machos não houve
diferenças significativas na latência entre as situações experimentais (X2 (3)= 3,319; p=0,345)
Fig. 12 A. Para as fêmeas houve diferença significativa entre as latências nas situações
experimentais (X2 (3)= 8,638; p=0,035). No entanto, através de testes post hoc foi verificado
que as diferenças entre as situações não eram significativas ao nível corrigido (Bonferroni
32 Figura 12: Mediana do tempo de latência na aquisição da primeira bala na condição dispersa A) para machos e B) para fêmeas (barras indicam intervalo de 95% de confiança; * p<0,008; pontos representam outliers).
Em relação às diferenças nas latências entre machos e fêmeas testes de Mann-Withnay
demonstraram haver diferenças entre duas situações, Balas Laranja e Fundo Verde (U= 710;
p=0,043) e Balas Roxas e Fundo Verde (U= 579; p=0,002) sendo que em ambas as fêmeas
foram mais rápidas que os machos. As outras situações não apresentaram diferenças (U> 720;
p> 0,408) Fig. 13. A)
33 Figura 13: Mediana do tempo de latência na aquisição da primeira bala por machos e fêmeas na condição dispersa (barras indicam intervalo de 95% de confiança; * p<0,05; pontos representam outliers).
Para a condição de recurso concentrado foram efetuadas as mesmas análises da
condição dispersa. Para ANOVA de Friedman houveram diferenças significativas tanto para
machos ( 2 (3)= 61,96; p<0,001) quanto para fêmeas ( 2(3)= 25,71; p=0,001) em relação às
situações experimentais. Análises post-hoc (Wilcoxon), demonstraram haver diferenças
significativas entre todas as situações para os machos (z<3,84; p<0,0001) menos entre a
situação de Bala Roxa e Fundo Verde e a de Bala Laranja Redonda e Fundo Verde e Laranja
(z= 1,147; p>0,05) Fig. 14 A. Já em relação às fêmeas, a situação de Balas Verdes e Fundo
Verde diferiu significativamente da de Balas Roxas e Fundo Verde (z= 3,844; p<0,001).
Houve também diferença significativa entre a situação de Balas Laranjas e Fundo Verde e
Balas Roxas e Fundo Verde (z= 3,736; p<0,001) com um maior consumo de balas roxas pelas
fêmeas, Fig. 14 B.
34 Figura 14: Mediana do número de balas consumidas pelos machos em cada sessão experimental da condição concentrada A) para machos (* p<0,001) e B) para fêmeas (* p<0,001) (barras indicam intervalo de 95% de confiança; pontos representam outliers).
Foram realizados testes de Mann-Whitney para cada uma das situações da condição
concentrada para verificar as diferenças no sucesso de machos e fêmeas. Houve diferença
significativa no consumo de balas na situação de Balas Laranjas Redondas e Fundo Verde e *
*
*
* *
* A)
35
Laranja (U=709; p=0,029), com os machos consumindo mais balas que as fêmeas. Não
houveram diferenças no consumo para as outras situações (U>844; p>0,05), Fig. 15.
Figura 15: Mediana do número de balas consumidas por sexo na condição concentrada (barras indicam intervalo de confiança de 95%; * p<0,05; pontos representam outliers).
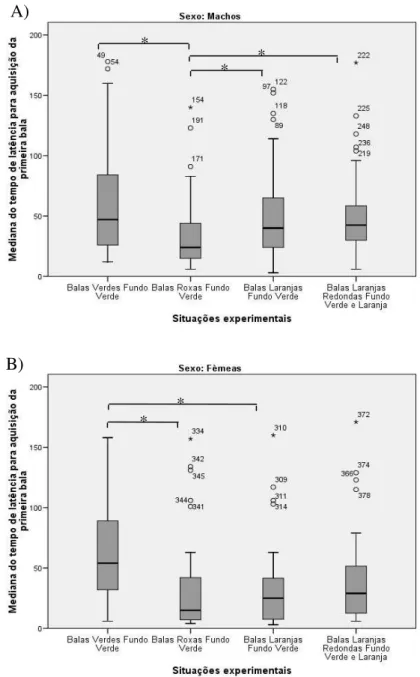
Para analisar a latência da aquisição dos itens alimentares na condição concentrada
foram realizadas as mesmas análises estatísticas. Tanto para machos quanto para fêmeas
houve diferenças significativas nas latências entre as situações experimentais nesta condição
(Machos X2 (3)= 19,541; p<0,001. Fêmeas X2 (3)= 20,921; p<0,001). Para os machos a
diferença se deu entre as situações Bala Verde e Fundo Verde e Bala Roxa e Fundo Verde
(Z= 4,333; p<0,001), Balas Laranjas e Fundo Verde e Balas Roxas e Fundo Verde (Z= 2,856;
p= 0,004) e Balas Laranjas Redondas e Fundo Verde e Laranja e Balas Roxas e Fundo Verde
(Z= 3,491; p<0,001), sendo que para todas as comparações a situação de Balas Roxas e Fundo
Verde apresentou as menores latências, Fig. 16 A. Em relação às fêmeas a situação Bala
36
2,769; p= 0,006) assim como a de Balas Verdes e Fundo Verde e Bala Roxa e Fundo Verde
(Z= 3,297; p=0,001) sendo que em ambas comparações a situação de Balas Verdes e Fundo
Verde foi a que apresentou as maiores latências, Fig. 16 B.
Figura 16: Mediana do tempo de latência para aquisição da primeira bala na condição concentrada A) para machos e B) para fêmeas (barras indicam intervalo de 95% de confiança; * p<0,008; pontos representam outliers).
Comparando os dois sexos em relação às latências nas diferentes situações através de
testes de Mann-Withnay encontrou-se que existem diferenças significativas apenas entre uma A)
B)
* * *