U IVERSIDADE FEDERAL DO RIO GRA DE DO ORTE DEPARTAME TO DE FISIOLOGIA
PROGRAMA DE PÓS GRADUAÇÃO EM PSICOBIOLOGIA
A I FLUÊ CIA DA RECOMPE SA E DA EXPERIÊ CIA A
DISCRIMI AÇÃO VISUAL SIMULTÂ EA EM SAGUI COMUM
(Callithrix jacchus)
TAULLI BRAGA LIMA
TAULLI BRAGA LIMA
A I FLUÊ CIA DA RECOMPE SA E DA EXPERIÊ CIA A
DISCRIMI AÇÃO VISUAL SIMULTÂ EA EM SAGUI COMUM
(Callithrix jacchus)
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para a obtenção do título de mestre em Psicobiologia.
Orientadora: Profa Dra Fívia de Araújo Lopes
Seção de enformação e Referência
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Lima, Taulli Braga.
A influência da recompensa e da experiência na discriminação visual simultânea em sagüi comum (Callithrix jacchus) / Taulli Braga Lima. – Natal, RN, 2012.
83f.
Orientadora: Fívia de Araujo Lopes.
Dissertação (Mestrado em Psicobiologia) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Departamento de Fisiologia. Programa de Pós9Graduação em Psicobiologia.
1. Sagui – Dissertação. 2. Callithrix jacchus 9 Dissertação. 3. Discriminação visual simultânea 9 Dissertação. 4. Influência da recompensa – Dissertação. I. Lopes, Fívia de Araujo. II. Universidade Federal do Rio Grande do Norte. III. Título.
TAULLI BRAGA LIMA
A I FLUÊ CIA DA RECOMPE SA E DA EXPERIÊ CIA A
DISCRIMI AÇÃO VISUAL SIMULTÂ EA EM SAGUI COMUM
(Callithrix jacchus)
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para a obtenção do título de mestre em Psicobiologia.
BANCA EXAMINADORA
Profa Dra Nicola Schiel
Universidade Federal Rural de Pernambuco
Profa Dra Maria de Fátima Arruda Universidade Federal do Rio Grande do Norte
Profa Dra Fívia de Araújo Lopes Universidade Federal do Rio Grande do Norte
Este trabalho foi escrito de acordo com o formato do
“O conhecimento torna a alma jovem e diminui a amargura da velhice. Colhe, pois, a sabedoria. Armazena suavidade para o amanhã.”
Agradecimentos
Agradeço à minha tia Lilia que me apoiou e auxiliou. A minha mãe pelo suporte
financeiro para que eu pudesse vir a Natal terminar a graduação e fazer o mestrado.
Aos pesquisadores da base do Professor Wagner Molina, principalmente, à
Leonardo Calado, Uedson, grandes amigos que sempre me deram força e me apoiaram
nos momentos em que precisei.
A Felipe Nalon Castro, Wallisen Tadashi Hattori e Pablo (argentino) que
ajudaram bastante com os testes estatísticos.
A professora Maria de Fátima Arruda que forneceu sugestões valiosas para a
execução da segunda fase projeto. E pelas discussões de artigos que eram realizados
semanalmente que contribuiu para maior aquisição de conhecimento. Aos professores
Daniel Marques de Almeida Pessoa e Maria Teresa da Silva pela concessão dos animais
para a realização do projeto.
Aos colegas da turma pelos momentos de descontração que foram
indispensáveis para relaxar em momentos que estava cansado e estressado.
A CAPES pela concessão da bolsa que foi indispensável para a realização do
projeto.
A Thieza, Thiago (namorado de Priscilla) e Priscilla que auxiliaram com dicas
importantes para a montagem do aparato que foi utilizado em ambas as fases do
trabalho. Assim como pelos momentos de descontração e apoio.
A professora Fivia de Araujo Lopes pela orientação e pela paciência.
Aos funcionários do Núcleo de Primatologia por terem proporcionado cuidados
Resumo
A aprendizagem discriminatória é analisada através de tarefas instrumentais nas quais o indivíduo é recompensado por escolher um item ao invés de outro. Desse modo, na discriminação visual simultânea de objetos o animal deve aprender que somente um dos objetos será recompensado. A discriminação visual simultânea é relativamente simples, e já foi observado que Callithrix jacchus é capaz de realizar essa tarefa. Como ainda não foi verificada a influência dos aspectos qualitativos das recompensas no desempenho da discriminação visual simultânea de objetos em primatas não9humanos, e como na maioria dos testes são utilizados animais isolados, o presente trabalho apresentou duas etapas: na primeira tivemos como objetivo analisar a influência do valor calórico da recompensa sobre o desempenho na discriminação visual simultânea de objetos em animais isolados; na segunda, tivemos o intuito analisar o desempenho de C. jacchus na realização da tarefa discriminatória em diferentes contextos sociais, assim como, analisar a influência da experiência anterior no desempenho da tarefa. Na primeira etapa (Estudo 1), os animais não foram capazes de discriminar os alimentos que apresentavam pequenas diferenças calóricas. Essa incapacidade em discriminar as recompensas foi responsável por gerar aleatoriedade na tarefa de discriminação visual simultânea de objetos. Na segunda etapa (Estudo 2), observamos que, independente do contexto social no qual a tarefa foi apresentada, o desempenho tanto dos animais experientes quanto dos animais inexperientes tendeu a aleatoriedade. No primeiro caso, é provável que o padrão de respostas dos animais experientes seja um reflexo do seu próprio desempenho quando foram observados isoladamente. No segundo caso, por sua vez, a aleatoriedade foi devida provavelmente ao pequeno número de sessões. Apesar de apresentar padrão de desempenho semelhante aos indivíduos inexperientes, nós verificamos que os animais experientes monopolizaram o consumo do alimento quando estavam na presença de indivíduos inexperientes. Isso foi uma consequência dos animais experientes terem apresentado menor latência de aproximação do aparato e, consequentemente, obterem mais alimento. Por sua vez, os animais inexperientes, quando estavam na presença de experientes, tiveram que adotar estratégias alternativas para obter o alimento. Desse modo, C. jacchus é capaz de utilizar a informação anterior que tinha sobre a resolução da tarefa em benefício próprio.
Abstract
The discrimination learning is assessed through instrumental tasks in which the individual is rewarded for choosing one item over another. Thus, in concurrent visual discrimination of objects the animal must learn that only one of the objects will be rewarded. The concurrent visual discrimination is relatively simple, and already been observed Callithrix jacchus is able to accomplish this task. As yet wasn't seen the influence of the qualitative aspects of the rewards, in the performance of concurrent visual discrimination of objects in nonhuman primates, and as in most tests are used isolated animals, the present study had two stages: at first we had as objective to analyze the influence of the caloric value of the reward on the performance in concurrent visual discrimination of objects in isolated animals; in the second, we had the intention analyze performance of C. jacchus in realization of discrimination task in different social contexts, as well as, analyze the influence of previous experience in task performance. In the first stage (Study 1), the animals were not able to discriminate foods that presented small caloric differences . This incapacity in discriminates the rewards was responsible by generating randomness in task of concurrent visual discrimination of objects. In the second stage (Study 2), observed that, independent of social context in which the task was presented, the performance both of the experienced animals as the inexperienced animals tended to randomness. In the first case, is likely that the pattern of responses of the experienced animals is a reflection of their own performance when they were observed in isolation. In the second case, in turn, the randomness was probably due to the small number of sessions. Although present a pattern of performance similar to inexperienced individuals, we verify that the experienced animals monopolize the food consumption when they were in the presence of inexperienced individuals. This was a consequence of the experienced animals have presented lower latency the approximation of apparatus and, consequently, obtain more food. In turn, the inexperienced animals, when were in the presence of experienced, had to adopt alternative strategies to obtain food. Thus, C. jacchus is able to use the previous information he had about the task of solving their own benefit.
Lista de Figuras
ESTUDO 19 A I FLUÊ CIA DA RECOMPE SA A DISCRIMI AÇÃO
VISUAL SIMULTÂ EA O SAGUI DO TUFO BRA CO (Callithrix jacchus)
Figura 1. Aparato experimental utilizado no experimento.(a) Alimentos presente no poço do aparato (b) Alimentos presentes no interior das formas geométricas tridimencionais de plástico. Neste exemplo, no quadrado foi colocado manga e no triângulo melancia ...21
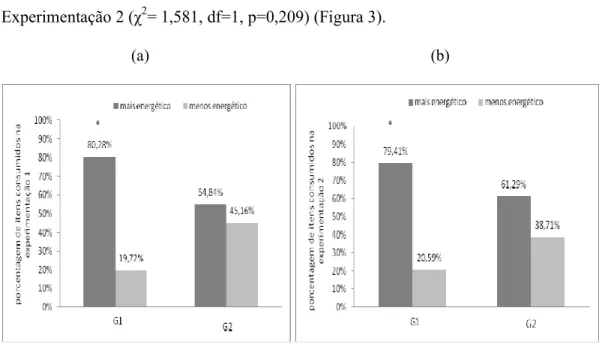
Figura 2. Porcentagem dos alimentos que as fêmeas consumiram nos grupos nas diferentes etapas do experimento: (a) Experimentação 1 (b) Experimentação 2.* Fêmeas consumiram mais significativamente alimentos mais energéticos do que menos energéticos. ** Fêmeas consumiram mais significativamente alimentos menos energéticos do que mais energéticos. ...24
Figura 3. Porcentagem dos alimentos que os machos consumiram nos grupos nas diferentes etapas do experimento: (a) Experimentação 1 (b) Experimentação 2.* Machos consumiram mais significativamente alimentos mais energéticos do que menos energéticos...25
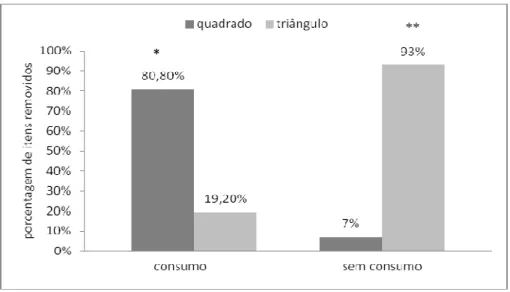
Figura 4. Remoção das formas pelas fêmeas no G1. Após essa remoção verificamos tanto eventos seguidos (esquerda) ou não de consumo (direita).* O quadrado foi removido mais frequentemente do que o triângulo quando houve consumo. ** O triângulo foi removido mais frequentemente do que o quadrado quando não houve consumo...27
ESTUDO 2 9 I FLUÊ CIA DA EXPERIÊ CIA A DISCRIMI AÇÃO VISUAL
SIMULTÂ EA EM SAGUI COMUM (Callithrix jacchus)
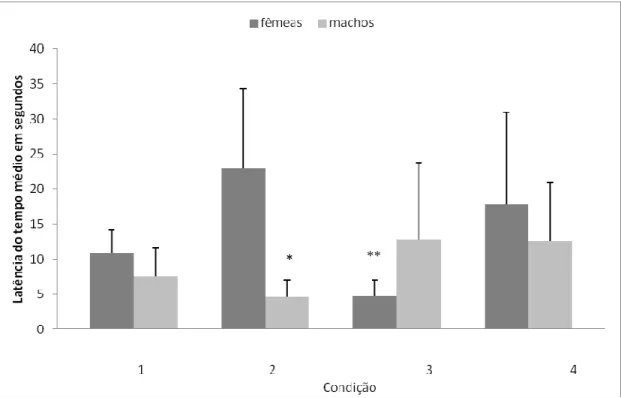
Figura 1. Latência média de aproximação em segundos de machos e fêmeas para cada uma das condições analisadas no presente trabalho: 1 machos experientes x fêmeas experientes; 2 machos experientes x fêmeas inexperientes; 3 fêmeas experientes x machos inexperientes; 4 machos inexperientes x fêmeas inexperientes.* Machos experientes tiveram tempo de aproximação significativamente inferior ao das fêmeas inexperientes.** Fêmeas experientes tiveram tempo de aproximação significativamente inferior ao dos machos inexperientes...50
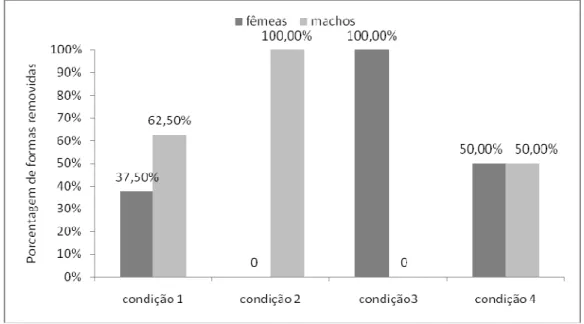
Figura 2. Porcentagem das formas removidas em cada uma das quatro condições. Nas Condições 2 e 3 somente os indivíduos experientes removeram as formas...52
Lista de Tabelas
ESTUDO 1 A I FLUÊ CIA DA RECOMPE SA A DISCRIMI AÇÃO
VISUAL SIMULTÂ EA O SAGUI DO TUFO BRA CO (Callithrix jacchus)
Tabela 1: Combinações dos alimentos utilizados em cada sessão acompanhados de seu valor energético...20
ESTUDO 2 9 I FLUÊ CIA DA EXPERIÊ CIA A DISCRIMI AÇÃO VISUAL
SIMULTÂ EA EM SAGUI COMUM (Callithrix jacchus)
Tabela 1: Formação dos casais para o experimento...45
Tabela 2: Prioridade de acesso ao alimento entre os membros dos casais...49
Sumário
INTRODUÇÃO GERAL ... 1
Callithrix jacchus (Linneaus, 1758): Caracterização e hábitos alimentares ... 2
Transferência de alimentos ... 5
O papel dos nutrientes no hábito alimentar ... 7
Discriminação Visual Simultânea de Objetos ... 10
OBJETIVOS E HIPÓTESES ... 12
Objetivo Geral ... 13
Objetivos específicos ... 13
Hipóteses e Predições ... 13
ESTUDO 1. A INFLUÊNCIA DA RECOMPENSA NA DISCRIMINAÇÃO VISUAL SIMULTÂNEA NO SAGUI DO TUFO BRANCO (Callithrix jacchus)...15
Introdução ... 17
Métodos ... 19
Resultados ... 24
Discussão ... 29
Referências bibliográficas ... 34
ESTUDO 2. INFLUÊNCIA DA EXPERIÊNCIA NA DISCRIMINAÇÃO VISUAL SIMULTÂNEA EM SAGUI COMUM (Callithrix jacchus)... ...39
Introdução ... 41
Resultados ... 49
Discussão ... 55
Conclusões... 61
Referências bibliográficas ... 62
DISCUSSÃO GERAL ... 67
Discussão Geral ... 68
Considerações finais ... 71
REFERÊNCIAS BIBLIOGRÁFICAS ... 72
ANEXOS ... 81
Anexo 1: Planilha de observação dos animais isolados ... 82
(Linneaus, 1758): Caracterização e hábitos alimentares
Callithrix jacchus (sagui comum) é um primata neotropical pertencente à Família Callithrichidae. Essa família faz parte da Infraordem Platyrrhine (primatas do
Novo Mundo) e é constituída por sete gêneros: Leontopithecus, Cebuella, Saguinus, Callithrix, Mico, Callimico (Rylands et al., 2000) e Callibella (Van Roosmalen & Van Roosmalen, 2003). Com exceção do Callimico, todos os demais membros dessa família são caracterizados por não possuir o terceiro molar e por gerar uma prole gemelar a
cada período reprodutivo (Cortés9Ortiz, 2009; Fleagle, 1988). Além disso, todos os
membros dessa família apresentam tamanho corporal reduzido, hábitos diurnos, cauda
não preênsil, pequeno peso corporal, e unhas em forma de garra em todos os dígitos
exceto o hállux (Fleagle, 1988; Garber, 1992; Sussman & Kinzey, 1984).
O sagui comum é endêmico do Brasil, sendo encontrado nos estados de Alagoas,
Pernambuco, Paraíba, Rio Grande do Norte, Maranhão, Ceará e Piauí, além de também
já ter sido verificada a sua introdução na região sul e sudeste do país (Rylands et al., 1993). Habita praticamente todos os tipos de vegetação com uma marcante preferência
pelas florestas secundárias e habitats alterados ou em processo de regeneração natural
(Rylands & Faria, 1993).
Esse primata vive em grupos estáveis, normalmente constituídos por três a
quinze indivíduos; esses grupos, por sua vez, são constituídos pelo casal reprodutor, a
prole do casal e indivíduos não aparentados (Hubrecht, 1984; Silva & Cruz, 1993). O
casal reprodutor é dominante sobre todos os outros membros do grupo; caso exista mais
de uma fêmea reprodutora também existirá hierarquia entre as mesmas (Digby, 1995). O
seu sistema de acasalamento é flexível, tendo sido descrita a monogamia, a poliandria e
forma cooperativa, com a participação de todos os indivíduos do grupo, incluindo
juvenis e subadultos, bem como outros animais adultos não reprodutivos e o macho
reprodutor, sendo este último o principal responsável por carregar os filhotes (Sussman
& Kinzey, 1984; Yamamoto, 1991).
Com relação ao hábito alimentar, tal como Cebuella pygmaea e Callithrix penicillata, C. jacchus consome quantidades consideráveis de exsudado (resina, goma). Além do exsudado, o sagui comum pode incluir em sua dieta itens alimentares bastante
variados: frutos, insetos (besouros, formigas, aranhas, entre outros), pequenos
vertebrados (lagartos, filhotes de aves) e ovos de aves em sua dieta, mas, em função da
proporção dos itens, é considerada uma espécie exsudatívora9insetívora (Garber, 1992;
Sussman & Kinzey, 1984). Desse modo, é de se esperar que esse primata, assim como
os demais indivíduos da Família Callithrichidae, utilize diferentes técnicas de exploração e estratégias de forrageio (Sussman & Kinzey, 1984).
O exsudado é fonte rica tanto em cálcio quanto em carboidrato para esse
primata. Para se alimentar desse item, C. jacchus apresenta adaptações morfológicas e fisiológicas. As adaptações morfológicas seriam a dentição especializada (incisivos
alargados na forma de “coifa” que são utilizados para “roer” os troncos das árvores e
desencadear o fluxo de goma) e unhas em forma de garras modificadas que permitem a
esses primatas se agarrar a troncos de árvores para se alimentar tanto de goma, como
insetos (Feagle,1988; Garber, 1992; Sussman & Kinzey, 1984). A adaptação fisiológica
seria a presença de um cecum modificado que apresentaria vários pequenos “bolsos”
que abrigam bactérias responsáveis por fermentar os polissacarídeos presentes na goma
(Power & Myers, 2009).
inteiros na boca e retirar parte do pericarpo, jogando fora a semente (no caso de frutos
pequenos), ou utilizar as mãos para levar o fruto até a boca no caso de frutos maiores
(Alonso & Langguth, 1989).
O consumo de insetos lhes dá acesso a um recurso alimentar rico em proteínas.
A estratégia adotada para a captura das suas presas depende do tamanho da presa e sua
capacidade de fuga. Nas presas maiores que dois centímetros, por exemplo, que
apresentam capacidade de fuga, o sagui exibe padrão mais cauteloso de locomoção
seguido de captura utilizando as mãos. Já ao capturar formigas e térmitas, por exemplo,
os saguis utilizam padrão de locomoção não tão cauteloso e captura a presa com a boca.
Além disso, o forrageio de insetos surge cedo em C. jacchus, visto que infantes (com a idade 394 meses) já apresentam habilidade para capturar certas presas (Schiel et al.,2010).
Além das diferentes estratégias adotadas durante o forrageio por C. jacchus, o comportamento alimentar desse primata é caracterizado pelo fato de que as fêmeas
adultas reprodutoras têm acesso prioritário ao alimento. Esse acesso é assegurado pelos
altos níveis de agressividade das mesmas (Lopes, 2002; Matias, 2001 Yamamoto et al., 2004) e pela colaboração do macho reprodutor que também apresenta o comportamento
agonista em relação a outros membros do grupo (Lopes, 2002). A maior aquisição de
alimentos pelas fêmeas adultas de C. jacchus é devida também ao fato das mesmas apresentarem uma motivação maior do que os machos para procurar alimentos que são
difíceis de serem obtidos (Michels, 1998). Essa maior motivação das fêmeas adultas do
sagui comum reflete um desempenho superior aos machos em tarefas alimentares
Transferência de alimentos
A transferência de alimentos é o movimento de alimentos entre coespecíficos,
exceto amamentação, e normalmente envolve um animal fornecendo alimento a outro.
Essa transferência é observada em diversas espécies de animais (Stevens & Gilby,
2004), embora seja necessário salientar que a circulação de alimento entre vários
indivíduos também pode acontecer (Ruiz9Miranda et al.,1999).
Essa passagem do alimento entre os indivíduos é marcada por uma série de
comportamentos (Brown & Mack, 1978). Esses comportamentos podem advir do
doador (indivíduo possuí o alimento), e em alguns casos, são apresentados pelo receptor
(indivíduo que poderá receber o alimento) também. Já foi observado, por exemplo, que
o doador exibe vocalizações enquanto segura o alimento antes de transferi9lo. Esse
padrão comportamental é conhecido como oferta do item alimentar (Brown & Mack,
1978; Feistner & Price, 1990; Joyce & Snowdon, 2007; Rapaport & Brown, 2008; Ruiz9
Miranda et al.,1999). Além disso, os potenciais doadores também podem demonstrar resistência à partilha segurando o alimento mostrando relutância em liberá9lo, ou
mesmo se virar ou se afastar com o intuito de evitar a transferência de alimento
(Feistner & Price, 1990; Kasper et al.,2008; Rapaport, 2001; Rapaport & Brown, 2008; Xue & Su, 2011). Assim como o doador do alimento, o receptor também pode exibir
certos padrões comportamentais antes da transferência como vocalizações e se
aproximar do doador; solicitando assim a transferência (Feistner & Price, 1990; Price &
Feistner,2001; Rapaport & Brown, 2008; Ruiz9Miranda et al.,1999).
Em primatas têm sido observado que a transferência de alimento pode ocorrer
tanto do adulto em direção ao infante, quanto entre adultos. A transferência de
alimentos entre adultos já foi observada tanto em primatas do Velho9Mundo como
quanto em primatas do Novo Mundo como calitriquídeos (Brown & Mack, 1978;
Kasper et al.,2008; Rapaport, 2001; Ruiz9Miranda et al.,1999), Cebus (de Waal, 1997a, 2000); macacos9coruja (Wolovich, et al., 2006), mico de cheiro (Stevens,2004) e macaco guigó (Fragaszy & Mason, 1983).
A transferência de alimento adulto9infante tem como intuito: transmitir aos
infantes itens alimentares difíceis de serem encontrados ou que necessitam de alguma
habilidade para obtê9lo; promover o desmame precoce do infante; fornecer itens
alimentares raros; e aprendizagem dos itens alimentares adequados (Ruiz9Miranda et al.,1999). Além disso, a transferência de alimentos tem o intuito de ensinar aos infantes onde obter alimento (Rapaport & Brown, 2008; Ruiz9Miranda et al.,1999).
A transferência de alimento entre adultos, por sua vez, pode surgir como produto
do altruísmo recíproco (de Waal, 1997b, 2000; Stevens & Gilby, 2004). Já foi
observado, por exemplo, que chimpanzés podem trocar alimento por catação
(“grooming”). Desse modo, se um indivíduo A cata um indivíduo B; o indivíduo B
tende a transferir alimento para A (de Waal, 1997b). Rapaport (2001), estudando
transferência de alimento entre micos9leões adultos (Leontopithecus spp.) também verificou, em um dos seus grupos, uma relação recíproca entre catação e partilha de
alimento; além disso, também foi observada relação recíproca entre catação e
coalimentação num comedouro monopolizável em Ateles geoffroyi (Pastor9Nieto, 2001). Fêmeas de Cebus apella também transferem alimento reciprocamente. Essa transferência recíproca, por sua vez, foi devida à simetria do comportamento afiliativo
entre as duas fêmeas (de Waal, 2000).
A partilha de alimento entre adultos também pode ser atribuída ao fato de que o
proprietário do alimento pode transferir parte dele com o intuito de evitar agressão e/ ou
proprietário iria se beneficiar por partilhar parte do alimento com o animal que pede o
alimento com o intuito de evitar danos ou custos energéticos gerados pela interferência
(Stevens,2004; Stevens & Gilby, 2004; Stevens & Stephens, 2002). Essa interferência
(assédio), por sua vez, pode ocorrer através da tentativa de roubo do alimento, roubo e
ou agressão física, por exemplo (Stevens,2004). Para aplicar esse modelo é necessário
que duas predições sejam mantidas: o assédio (interferência) deve ser oneroso para o
dono do alimento; e o aumento do assédio deve aumentar a probabilidade com que o
animal que pede alimento tenha acesso ao mesmo (Stevens & Gilby, 2004). Esse
modelo de partilha tem sido amplamente utilizado pelas seguintes razões: a partilha
fornece benefícios imediatos ao dono do alimento visto que evita o custo do assédio;
esse modelo não requer habilidades cognitivas complexas;além disso, ele não requer
que o doador e o receptor tenham relação especial (sejam parentes, por exemplo)
(Stevens,2004). Em Saimiri boliviensis e Pan troglodytes já foi observada uma relação direta entre frequência da partilha e assédio; desse modo, quanto maior o assédio mais
frequente a partilha do alimento (Stevens, 2004).
A transferência de alimento entre adultos e/ou entre adulto9infante pode também
ter surgido devido à seleção de parentesco. Em Leontopithecus spp., por exemplo, já foi observado que adultos ou sub9adultos vivendo em seu grupo natal têm tendência de
transferir mais alimentos para a mãe do que para ajudantes do sexo oposto. Entretanto,
nesse caso a partilha de alimento pode ter como o intuito evitar agressão da fêmea
reprodutora (Rapaport, 2001).
O papel dos nutrientes no hábito alimentar
De acordo com a teoria do forrageio ótimo, os indivíduos que maximizam a
atividades que beneficiem o seu fitness do que indivíduos que forrageiam de modo
menos eficiente (não são capazes de maximizar a ingestão de calorias) (Stephens et al., 2007). Com base nisso, é imperativo que os animais selecionem itens alimentares que
proporcionem um maior retorno energético após o consumo (Alcock, 2001).
A informação sensorial possui um papel fundamental na seletividade dos
alimentos, visto que, as pistas sensoriais advindas dos alimentos carregam consigo
informações importantes sobre os mesmos (Dominy et al., 2001). Já foi observado, por exemplo, que primatas apresentam sensibilidade olfatória elevada para determinadas
moléculas odoríferas presentes nos frutos como o álcool (Laska et al., 2006), e que essa molécula é utilizada como pista química para localizar e avaliar o conteúdo nutricional
de determinados frutos, uma vez que a concentração dessa substância está diretamente
relacionada com a concentração de açúcares solúveis (Dominy, 2004).
Além disso, o trabalho de Laska (1996) demonstrou que Saimiri sciureus utiliza a gustação para selecionar itens alimentares com maior conteúdo de açúcar, e desse
modo, com maior valor energético, sendo verificado que esse primata é capaz de
discriminar diferentes concentrações de sacarose (Laska, 1994). Já foi observado
também que através da gustação C. jacchus é capaz de distinguir água de uma solução de cálcio, exibindo clara preferência pela solução de cálcio (Power et al., 1999). Nesse
último trabalho, além da discriminação descrita, os autores observaram ainda, que as
fêmeas reprodutoras ingeriram uma quantidade superior da solução de cálcio em relação
aos outros indivíduos.
Além da olfação e da gustação, já foi observado que primatas podem utilizar
também a visão e o tato para analisar a qualidade dos alimentos. Em macacos aranha
mais pistas táteis para avaliar alimentos novos (Laska et al., 2007). Além disso, há trabalhos que mostram que a textura pode ser um indicador preciso do status nutricional
do fruto (Dominy, 2004). Com relação à visão, por sua vez, trabalhos como o de Riba9
Hernández et al. (2005) mostram que certos frutos maduros exibem coloração que está diretamente relacionada com a concentração de glicose.
É necessário salientar que juntamente com feedback sensorial, os animais podem utilizar feedback pós9ingesta para analisar a composição dos alimentos. C. apella, por exemplo, ao ser exposto a combinações binárias de alimentos novos apresentava
inicialmente preferência correlacionada positivamente com o conteúdo de glicose e
frutose. Posteriormente, a sua preferência foi correlacionada positivamente com o
conteúdo de energia total. Desse modo, esse primata é capaz de associar as propriedades
sensoriais do alimento a fatores pós9ingesta; essa interação, por sua vez, irá direcionar a
sua seleção a alimentos que tenham maior conteúdo energético (Visalberghi et al., 2003).
Por fim, deve ser enfatizado que a tomada de decisão no forrageio também pode
ser influenciada por outros fatores como a familiaridade do animal com o alimento, a
sazonalidade e a interação social. Já foi observado, por exemplo, que macacos rhesus adultos (Macaca mullata) discriminavam alimentos novos de alimentos conhecidos e
exibiam clara preferência por alimentos conhecidos em detrimento dos alimentos novos.
Esse padrão comportamental permanecia mesmo quando os animais eram submetidos à
privação de comida; desse modo, a neofobia alimentar exerce papel na seleção da dieta
nesse primata (Johnson, 2000). De modo similar, machos adultos de C. jacchus exibiram cautela maior do que as fêmeas adultas diante da possibilidade da ingestão de
alimentos novos (Lacerda, 2004; Lopes, 2002). Esse mesmo padrão foi verificado entre
aos selvagens. Assim, nesse primata, a neofobia é responsável também por influenciar a
competição alimentar. Já que o alimento novo pode desencadear diferentes reações nos
animais (Lopes, 2002).
Com relação a influência da sazonalidade na tomada de decisão durante o
forrageio, já foi observado que C. jacchus na estação seca pode consumir mais goma com o intuito de compensar a escassez de frutos (Castro & Araújo, 2006). Por sua vez,
com relação à influência da interação social, já foi verificado que pares de C. jacchus ao terem sua preferência alimentar artificialmente alterada (tornando o alimento preferido
impalatável),foram capazes de restabelecê9la ao interagir com indivíduos que não
passaram por esse procedimento (Queyras et al., 2000).
Discriminação Visual Simultânea de Objetos
No sentido literal, o termo discriminação refere9se à capacidade de detectar
diferenças. Os estudos de discriminação têm como objetivo analisar a habilidade animal
em responder diferentemente a itens ou locais no ambiente, assim como, determinar o
limiar sensorial do animal (O´Keefe & Nadel, 1978).
A aprendizagem discriminatória é analisada através de tarefas instrumentais nas
quais o objeto de estudo é recompensado por escolher um estímulo ao invés de outro.
Esse estímulo pode ser apresentado simultaneamente ou sucessivamente (Shettleworth,
2010; Tomasello & Call, 1997).
A discriminação visual simultânea de objetos utiliza pares de objetos que são
apresentados simultaneamente, sendo que o indivíduo é recompensado se escolher um
Essa tarefa é um modo de se analisar a memória de hábitos (Bachevalier et al., 1990) que está relacionada com a associação do estímulo à resposta, sendo armazenada
através da tentativa e erro (Bachevalier et al., 1990). Como essa memória está funcionalmente madura em infantes de primatas não9humanos (Bachevalier & Mishkin,
1984), é de se esperar que a tarefa de discriminação visual simultânea de objetos seja
um teste simples de ser realizado inclusive por primatas jovens. De fato, já foi
verificado que C. apella jovens (com idade variando entre 1 a 3 anos e meio) já são capazes de realizar tarefa de discriminação simultânea de objetos (Resende et al., 2003). Além disso, foi observado que infantes de macacos rhesus com três meses de idade eram capazes de realizar a discriminação visual tão rapidamente quanto adultos
(Bachevalier & Mishkin, 1984).
Apesar de apresentar desempenho inferior se comparado ao gênero Saimiri spp. e Macaca spp.(Deaner et al., 2006), C. jacchus é capaz de realizar tarefas de discriminação visual simultânea de objetos (Croft et al., 1999; Maclean et al., 2001; Ridley et al., 2001). Tem sido utilizado como modelo experimental tanto pela psicologia comparativa como pela etologia e neurociência cognitiva pelas facilidades de
manuseio e manutenção do animal (Stevenson, 1977), havendo crescente interesse no
processo de aprendizagem discriminatória por psicólogos comparativos (Hauser et al.,
2002). Assim, a escolha do sagui como objeto de estudo justifica9se como tentativa de
melhor compreender algumas particularidades relacionadas a esse processo, como a
influência de propriedades dos estímulos utilizados bem como as influências sociais em
Objetivos
Objetivo Geral
O objetivo deste trabalho foi analisar a influência do aspecto qualitativo da
recompensa sobre o desempenho na tarefa de discriminação visual simultânea em sagui
comum (Callithrix jacchus) isolados e em diferentes contextos sociais.
Objetivos específicos
Analisar a capacidade de C. jacchus em discriminar os alimentos mais energéticos dos menos energéticos (Estudo 1).
Analisar a influência da experiência anterior no desempenho da tarefa. (Estudo 2).
Hipóteses e Predições
Hipótese 1 – A preferência alimentar será influenciada pelo valor calórico dos
alimentos (Estudo 1).
Predição 1 Como a teoria do forrageio ótimo prevê que os animais selecionam
itens alimentares que proporcionem um maior retorno energético (Alcock, 2001;
Stephens et al., 2007), e como já foi observado em primatas do Novo Mundo quea preferência alimentar está correlacionada positivamente com a quantidade
Hipótese 2 – A preferência alimentar irá influenciar o desempenho na tarefa de
discriminação visual simultânea em C. jacchus (Estudo 1).
Predição 2 – Devido ao fato de que já foi observado que C. jacchus é capaz de realizar a associação da forma com alimento (Croft et al., 1999; Maclean et al., 2001; Ridley et al., 2001), esperamos que esse primata consiga associar os alimentos preferidos com a forma geométrica.
Hipótese 3 A experiência prévia com a tarefa irá influenciar no desempenho da
mesma (Estudo 2).
Predição 3.1 – Como foi verificado em C. jacchus que indivíduos que tinham experiência anterior com tarefa instrumental atuavam como produtores e ao
interagirem com indivíduos inexperientes chegavam primeiro ao aparato e
realizavam a tarefa (Caldwell & Whiten, 2003). Esperamos que os indivíduos
que já tiveram contato anterior com a tarefa apresentem latência de aproximação
inferior em relação aos indivíduos inexperientes.
Predição 3.2 – Os indivíduos que apresentarem maior latência de aproximação
A I FLUÊ CIA DA RECOMPE SA A DISCRIMI AÇÃO VISUAL
SIMULTÂ EA O SAGUI DO TUFO BRA CO (Callithrix jacchus)
Taulli Braga Lima & Fívia de Araújo Lopes
Programa de Pós9Graduação em Psicobiologia, Universidade Federal do Rio Grande do
Norte, Centro de Biociências, Caixa Postal 1511; Campus Universitário Lagoa Nova,
CEP 590789970, Natal, Rio Grande do Norte, Brasil. e9mail: [email protected].
Resumo A discriminação visual simultânea de objetos é caracterizada pela apresentação simultânea de pares de objetos; de modo que, os indivíduos devem aprender que somente um dos objetos de cada par é recompensado. Tendo em vista que ainda não foi verificado o estimulo qualitativo da recompensa na discriminação visual simultânea de objetos em primatas não9humanos, o presente trabalho teve o intuito de analisar o papel do valor calórico das recompensas no desempenho da tarefa da discriminação visual simultânea em Callithrix jacchus. Inicialmente, analisamos a capacidade desse primata em discriminar qualitativamente quatro pares de recompensas que foram distribuídas em dois grupos: grupo 1 (G1 9 constituído por pares de alimentos que apresentavam maior diferença calórica) e grupo 2 (G2 9 constituído por pares de alimentos que apresentavam menor diferença calórica). Para isso, verificamos o consumo de um dos itens alimentares de cada par. Feito isso, analisamos a capacidade de C. jacchus associar os alimentos mais energéticos com formas geométricas. Ambas as etapas do experimento totalizaram 240 sessões de observação. Na discriminação qualitativa das recompensas, foi observado que tanto machos quanto fêmeas exibiram marcante preferência pelas recompensas mais calóricas em detrimento das menos calóricas no G1; já no G2, nenhum dos sexos foi capaz de discriminar qualitativamente as diferentes recompensas. Essa incapacidade em discriminar as recompensas no G2, por sua vez, foi responsável por gerar aleatoriedade na discriminação de objetos. Desse modo, foi possível concluir que a qualidade da recompensa influencia no desempenho da aprendizagem discriminatória para esses animais.
Introdução
A formação da memória é considerada um dos mecanismos responsáveis por
alterar o comportamento (Kandel et al., 2000). Há várias teorias que a tem dissociado em dois tipos diferentes: memória explícita, que está relacionada a fatos e eventos; e
implícita que está relacionada a habilidades, hábitos e procedimentos (Corkin, 1968;
Shettleworth, 2010). A memória de hábitos “atua” de modo inconsciente e independente
da memória explícita, sendo armazenada através da tentativa e erro. Esse tipo de
memória envolve a lenta aquisição da associação do estímulo com a resposta (Bayley et al., 2005).
Um modo de se analisar a memória de hábitos é através das tarefas de
aprendizagem discriminatórias (Bachevalier et al., 1990). Tais tarefas caracterizam9se
por procedimentos nos quais os animais são recompensados por escolher um estímulo
ao invés de outro. Estes estímulos, por sua vez, podem ser sucessivos ou simultâneos
(Shettleworth, 2010; Tomasello & Call, 1997).
Têm sido empregados dois tipos de procedimentos operantes para a análise da
discriminação visual em animais: no procedimento ir/ não ir (Go/&ot Go), a resposta é recompensada na presença de um estímulo mas não na presença de outro; já o segundo
procedimento, envolve a escolha entre duas ou mais alternativas de respostas associadas
a um estímulo (Chase & Heinemann, 2001).
O paradigma da discriminação visual simultânea de objetos utiliza o segundo
procedimento, que é caracterizado pela apresentação simultânea de pares de objetos.
Para solucionar a tarefa os indivíduos têm que aprender que somente um dos estímulos
de cada par é recompensado (Deaner et al., 2006). Desse modo, o processo de discriminação simultânea requer que o indivíduo retenha a representação do estímulo e,
2003). Deve9se salientar, que quando o indivíduo é exposto a um grande número de
tarefas de discriminação é esperado que ele tenha progressiva melhora no seu
desempenho (Shettleworth, 2010, Tomasello & Call, 1997).
Há trabalhos que mostram que primatas do Novo Mundo são capazes de realizar
a tarefa de discriminação visual simultânea de objetos. Foi verificado que Cebus apella, por exemplo, é capaz de aprender tal discriminação utilizando vinte pares de estímulos.
Além disso, ele também foi capaz de reter a associação estímulo/recompensa com base
em testes únicos dentro de um intervalo de 24 horas (Resende et al., 2003).
Já foi verificado também que, além do Cebus, Callithrix jacchus é capaz de realizar tarefa de discriminação simples de objetos (Croft et al., 1999; Maclean et al., 2001; Ridley et al., 2001). Esse primata é encontrado em diversos estados brasileiros (Rylands et al., 1993), vive em grupos estáveis constituídos pelo casal reprodutor, a prole do casal e indivíduos não aparentados (Hubrecht, 1984; Silva & Cruz, 1993).
Além disso, C. jacchus tem sido utilizado como modelo tanto pela psicologia comparativa como pela etologia e neurociência cognitiva por apresentar: tamanho
corporal reduzido, o que permite fácil manuseio e manutenção (Stevenson, 1977), alta
taxa de reprodução, características dos sistemas de criação semelhantes ao dos homens,
efeitos comportamentais resultantes da administração de drogas semelhante aos dos
homens (Yamazaki & Watanabe, 2009).
São evidentes as vantagens de C. jacchus como modelo experimental e o crescente interesse no processo de aprendizagem discriminatória por psicólogos
comparativos (Hauser et al., 2002), porém até o presente momento não foi observado o aspecto qualitativo da recompensa no desempenho da discriminação visual simultânea
de objetos em primatas não9humanos. Portanto, o presente trabalho teve o intuito de
discriminação visual simultânea em C. jacchus. O nosso trabalho é baseado nas seguintes predições: 1) Como a teoria do forrageio ótimo prevê que os animais
selecionam itens alimentares que proporcionem um maior retorno energético com o
intuito de aumentar a sobrevivência e/ou o sucesso reprodutivo (Alcock, 2001; Stephens
et al., 2007), C. jacchus é capaz de discriminar recompensas, exibindo preferência pela recompensa mais energética em detrimento da menos energética; 2) A preferência pela
recompensa irá guiar a tomada de decisão do animal na tarefa de discriminação visual
simultânea de objetos; desse modo, é esperado que C. jacchus selecione a forma que está associada com o alimento de maior qualidade (mais energético).
Métodos
Local de Estudo e Animais
Realizamos o experimento no Núcleo de Primatologia da Universidade Federal
do Rio Grande do Norte (Natal, RN, Brasil). Utilizamos vinte animais adultos, dez
machos e dez fêmeas. Esses animais ficaram isolados durante todo o tempo; e não
somente durante a realização do experimento. Além disso, esses saguis nunca
participaram de nenhum experimento anterior. Mantivemos os saguis em gaiolas
individuais medindo 2 metros de altura por 2 de comprimento e 1 metro de largura.
Cada gaiola continha plataformas, galhos e uma caixa ninho. Alimentamos os animais
duas vezes ao dia com uma mistura de proteínas no início da manhã e frutas frescas no
início da tarde, com água ad libitum, sendo os mesmos mantidos em condições naturais de iluminação, temperatura e umidade do ar. Observamos as fêmeas durante o mês de
EstímuloAlimentar
Utilizamos itens alimentares conhecidos pelos animais, alguns dos quais
oferecíamos com maior frequência rotineiramente aos animais (todos os dias), a saber:
banana pacovã (Musa spp.), mamão papaia (Carica papaya L.) e melancia (Citrullus vulgaris Schrad.). Já outros itens alimentares, apesar de também serem conhecidos pelos animais, eram oferecidos com menor frequência(uma vez a cada duas semanas):
maçã argentina (Malus domestica), abacaxi (Ananas comosus), manga Tomy Atkims (Mangifera indica L.), melão (Cucumis melo L.) e uva passa (Vitis vinifera).
Para a realização do experimento, pareamos os alimentos em combinações,
compondo dois grupos: grupo 1 (G1), constituído dos pareamentos banana x abacaxi/
passa x melão; e grupo 2 (G2), formado pelos pares maçã x mamão/ manga x melancia.
A formação desses grupos foi baseada na diferença calórica entre os alimentos mais
energéticos e menos energéticos, de modo que, o G1 apresentou maior diferença
calórica entre os alimentos quando comparados aos pares de alimentos que compuseram
o G2 (Tabela 1). Cada item alimentar de cada pareamento apresentava o mesmo
tamanho.
Tabela 1: Combinações dos alimentos utilizados em cada sessão acompanhados de seu valor energético. ALIME TOS
MAIS E ERGÉTICOS
(+)
VALOR a E ERGÉTICO
ALIME TOS ME OS E ERGÉTICOS
( )
VALOR a E ERGÉTICO
G1 BANANA
b
78kcal X ABACAXI b 48kcal
UVA PASSA c 299kcal X MELÃO b 29kcal
G2 MACÃ
b 63kcal X MAMÃO b 40kcal
MANGAb 51kcal X MELÂNCIA b 33kcal
a
O valor energético dos alimentos foi calculado para 100 g de alimento. Esse valor foi obtido a partir dos teores em proteínas, lipídios e glicídios utilizando os coeficientes específicos que levam em consideração o calor de combustão e a digestibilidade (Lima et al., 2006)
b
Valor calórico obtido na Tabela Brasileira de Composição de Alimentos9TACO (Lima et al.,2006).
c
Procedimento
O aparato utilizado no experimento era composto por uma base de PVC
medindo 28 x 11,5 x 1,0cm, presa com arame a um suporte de plástico medindo 30 x 15
x 0,3cm (Figuras 1a e 1b). Nessa base havia dois poços (Figura 1a), distantes 14,5 cm:
um no formato de um triângulo, medindo 3,5 x 3,5 x 3,0 x 1cm e outro no formato de
um quadrado, medindo 3 x 3 x 1cm. No interior desses poços encaixamos duas formas
geométricas de plástico opacas (Figura 1b).
(a) (b)
Figura 1: Aparato experimental utilizado no experimento.(a) Alimentos presente no poço do aparato (b) Alimentos presentes no interior das formas geométricas tridimencionais de plástico. Neste exemplo, no quadrado foi colocado manga e no triângulo melancia (Fotos de Taulli Braga Lima).
Para a apresentação dos alimentos aos animais, fixamos esse aparato no portão
da gaiola do animal, a uma altura de aproximadamente 1,10m do chão com arame; e a
sessão de observação era iniciada quando o aparato era preso no lado interno da porta da
gaiola. O término ocorria após totalizar dez minutos ou quando havia o consumo de um
dos itens alimentares (o que ocorresse primeiro). Caso o consumo ocorresse dentro dos
dez minutos, consideraríamos a observação concluída, havendo a reposição dos novos
O experimento teve três fases: Adaptação, Experimentação 1 e Experimentação
2.
Adaptação: consistiu em apresentar somente a base para os animais. Essa fase foi
realizada com o intuito de habituar os animais ao experimentador e ao aparato em si. Ao
contrário das outras duas fases, essa não utilizou alimento. Durante esse processo foram
realizadas quatro sessões com a duração máxima de cinco minutos tanto no período da
manhã (entre 10:00911:00h) quanto no período da tarde (entre 15:00916:00h).
Experimentação 1: esta fase teve o intuito de analisar a capacidade de discriminação
entre recompensas de alta (mais energética) e baixa qualidade (menos energética).
Realizamos esse processo através de um teste de escolha entre duas alternativas
(apresentamos pares de itens alimentares aos animais e registramos qual item alimentar
o animal consumiu primeiro). Realizamos as observações no período da manhã, entre
07:40 e 09:30 horas, antes da primeira refeição diária dos animais. Utilizamos quatro
combinações diferentes de alimentos (Tabela1). Nessa fase, apresentamos os pares de
alimentos tanto no poço do aparato; quanto no interior de formas geométricas
tridimensionais de plástico (que encaixamos sobre os respectivos poços mantendo a face
aberta dessas formas voltada para cima) (Figura 1b). Convencionamos colocar no
quadrado o alimento mais energético e no triângulo o menos energético. Aleatorizamos
o posicionamento das formas para evitar vícios de resposta em se mantendo uma
apresentação sempre na mesma posição. Consideramos cumprida a fase de
Experimentação 1 caso o animal, após oito sessões consecutivas, apresentasse uma
eficiência de 90% na aquisição de qualquer item alimentar disponível.
Experimentação 2: nesta fase apresentamos as formas geométricas tridimensionais
alimento, o mesmo teria que remover uma das formas geométricas de um dos poços.
Assim como na fase de Experimentação 1, aleatorizamos a posição das formas
geométricas tridimensionais. Nesta etapa, pretendeu9se analisar a capacidade do sagui
em associar as características do objeto (quadrado) com a qualidade da recompensa (que
seria o alimento com maior valor calórico). Realizamos um total de oito sessões de
observação para cada animal; mantendo o procedimento adotado para a observação
nessa fase semelhante ao da fase de Experimentação 1.
No processo de Experimentação 1, nos realizamos um total de 160 sessões de
observação; já na Experimentação 2, realizamos um total de 80 sessões. Desse modo, a
fase de Experimentação 1 juntamente com a Experimentação 2 totalizou 240 sessões de
observação; de modo que, realizamos 60 sessões para cada uma das quatro combinações
de alimentos. Tanto no período de Experimentação 1 como no de Experimentação 2, o
animal tinha contato visual com o experimentador que ficava parado na frente da gaiola
e do animal. Com o intuito de evitar fornecimento de qualquer pista para o animal
durante a resolução da tarefa, o experimentador fixava o olhar em algum ponto do
aparato durante a tomada de decisão do animal.
Análise Estatística
Para realizar a análise da preferência alimentar em cada uma das fases do
experimento, bem como a análise do desempenho na tarefa de discriminação visual,
utilizamos o qui9quadrado de Pearson 9 χ2. Para todas as análises o nível de significância
Resultados
Consumo
Fêmeas
Ao se analisar o consumo das fêmeas, nos observamos, na Experimentação 1,
76 eventos de consumo e 4 eventos sem consumo no G1; no G2 nos verificamos 69
eventos de consumo e 11 eventos sem consumo.Já na Experimentação 2, nos
verificamos 37 eventos de consumo e 3 sem consumo no G1; no G2, nos observamos 36
eventos de consumo e 4 sem consumo.
Além disso, nos observamos que as fêmeas consumiram mais
significativamente alimentos mais energéticos em detrimento dos alimentos menos
energéticos no G1 tanto na Experimentação 1 (χ2= 53,895, df = 1, p<0,001) quanto na
Experimentação 2 (χ2= 16,892, df =1, p<0,001) (Figura 2). Já no G2, as fêmeas
consumiram os alimentos menos energéticos mais significativamente do que os
alimentos mais energéticos na Experimentação 1 (χ2=10,565, df=1, p=0,001); na
Experimentação 2 não observamos diferença significativa no consumo entre os itens
alimentares (χ2= 1,000, df=1, p=0,317) (Figura 2).
(a) (b)
Machos
No que se refere à análise dos machos, verificamos na Experimentação 1 71
eventos de consumo e 9 sem consumo no G1; no G2, observamos 62 eventos de
consumo e 18 sem consumo. Na Experimentação 2, verificamos 34 eventos de consumo
e 6 sem consumo no G1; no G2, observamos 31 eventos de consumo e 9 sem consumo.
No G1 os machos consumiram alimentos mais energéticos mais
significativamente do que os alimentos menos energéticos tanto na Experimentação 1
(χ2= 26,042, df=1, p<0,001) quanto na Experimentação 2 (χ2= 11,765, df=1, p<0,001)
(Figura 3); no G2, por sua vez, não observamos diferença significativa entre o consumo
dos itens mais e menos calóricos na Experimentação 1 (χ2= 0,581, df =1, p<0,486) e
Experimentação 2 (χ2= 1,581, df=1, p=0,209) (Figura 3).
(a) (b)
Discriminação Visual
Realizamos um total de oitenta sessões: na metade dessas sessões foram
utilizados o G1 e na outra metade o G2. Durante essas sessões, verificamos os seguintes
eventos: remoção da forma seguido pelo consumo do alimento ou remoção da forma
sem consumo do alimento presente no poço.
Fêmeas Grupo 1
Durante as sessões no G1, nos verificamos que as fêmeas removeram um total
de 40 formas; de modo que 26 foram removidas em eventos seguidos de consumo e 14
em eventos remoção sem consumo do alimento. A escolha das formas ficou dentro da
aleatoriedade, visto que não observamos diferença significativa na remoção entre o
quadrado e o triângulo (quadrado: 55%; triângulo: 45%; χ2= 0,400, df =1, p=0,517).
Entretanto, ao se considerar os eventos em que houve remoção seguida de consumo
observamos que as fêmeas removeram o quadrado mais frequentemente do que o
triângulo (quadrado: 80,8%; triângulo: 19,2 %; χ2= 9,84, df=1, p=0,002) (Figura 4); já
ao se analisar eventos de remoção sem consumo no G1, verificamos que as fêmeas
removeram o triângulo mais frequentemente do que o quadrado (quadrado: 7%;
Figura 4. Remoção das formas pelas fêmeas no G1. Após essa remoção verificamos tanto eventos seguidos (esquerda) ou não de consumo (direita).* O quadrado foi removido mais frequentemente do que o triângulo quando houve consumo. ** O triângulo foi removido mais frequentemente do que o quadrado quando não houve consumo.
Fêmeas Grupo 2
No G2 nos observamos apenas um evento onde não houve remoção das formas.
Desse modo, foram removidas um total de 39 formas geométricas; de modo que 20
foram removidas em eventos seguidos de consumo e 19 em eventos remoção sem
consumo do alimento. Assim como foi verificado no G1, não observamos diferença
significativa na remoção entre quadrado e triângulo no G2 (quadrado: 56,4%; triângulo:
43,6%; χ2= 0,641, df=1 p=0,423). Além disso, também não observamos remoção
diferencial das formas nem nas situações em que verificamos o consumo do alimento
presente no poço (quadrado: 45%; triângulo: 55%; χ2= 0,200, df=1, p=0,655), nem nos
eventos em que houve remoção da forma sem o consumo do alimento presente no poço
Machos Grupo 1
No G1, nos observamos um total de 37 eventos de remoção das formas e apenas
3 sem remoção. Desses 37 eventos, 24 formas foram removidas em eventos seguidos
de consumo e 13 em eventos sem consumo. Assim como no caso das fêmeas, não
verificamos diferença significativa na remoção das formas no G1 pelos machos
(quadrado: 54,1%; triângulo: 45,9%; χ2= 0,243, df=1, p=0,622). Porém, em eventos
onde a remoção foi seguida do consumo do alimento presente no poço observamos que
os machos removeram o quadrado mais frequentemente do que o triângulo (quadrado:
70,8% triângulo: 29,2%; χ2=4,167, df=1, p=0,041) (Figura 5). Já ao se analisar os
eventos de remoção sem consumo do alimento, verificamos que os machos removeram
o triângulo mais frequentemente do que o quadrado (quadrado: 23%; triângulo: 77%;
χ2= 4,216, df=1, p=0,040) (Figura 5).
Figura 5. Remoção das formas pelos machos no G1. Após essa remoção verificamos tanto eventos seguidos (esquerda) ou não de consumo (direita). * O quadrado foi removido mais frequentemente do que o triângulo quando houve consumo. ** O triângulo foi removido mais frequentemente do que o quadrado quando não houve consumo.
Machos Grupo 2
No G2, nos observamos um total de 39 eventos de remoção das formas e apenas
um evento onde não foi observado remoção. Desses 39 eventos de remoção, 27 formas
foram removidas em eventos seguidos de consumo e 12 em eventos sem consumo.
Assim como no G1, o comportamento dos machos tendeu à aleatoriedade, visto que,
não houve remoção diferencial das formas (quadrado: 56,41%; triângulo: 43,59%; χ2=
0,641, df=1, p=0,564). Assim como no caso das fêmeas, não observamos diferença
significativa na remoção das formas em situações com consumo (quadrado: 63%;
triângulo: 37%; χ2= 1,815, df=1, p=0,178) e sem consumo (quadrado: 41,66%;
triângulo: 58,33%; χ2(1)= 0,333, df=1, p=0,423).
Discussão
Dividimos o presente trabalho em dois momentos de avaliação: no primeiro
analisamos a capacidade de C. jacchus em discriminar diferentes recompensas, tomando como referência o consumo de itens alimentares com diferentes valores energéticos. No
segundo, analisamos o aspecto qualitativo da recompensa no desempenho da
aprendizagem discriminatória, levando em consideração a interação do animal com uma
forma geométrica específica e o consumo do item alimentar associado a ela.
O padrão de escolha entre recompensas pode ser feito de acordo com o valor
recompensa). Deve9se salientar, entretanto, que a preferência por recompensa é relativa
e irá depender de outras alternativas viáveis (Hassani et al., 2001; Tremblay & Schultz, 1999; Cromwell et al., 2005). No presente trabalho, inferimos o valor motivacional das recompensas através da preferência alimentar, tendo como base o consumo dos itens
alimentares. Observamos que quando havia maior diferença quanto ao valor calórico
entre os itens alimentares (G1) os animais apresentaram preferência pelos alimentos
mais calóricos em detrimento dos alimentos menos calóricos. Desse modo, nesse grupo,
os animais foram capazes de discriminar qualitativamente as diferentes recompensas
apresentadas. Esse resultado se assemelha aos de trabalhos que ilustram que em
primatas do Novo Mundo a preferência alimentar está diretamente relacionada com o
conteúdo energético (Laska et al., 2000; Laska, 2001).
Já no G2, os animais não foram capazes de avaliar qualitativamente as diferentes
recompensas apresentadas. Evidenciamos isso pela ausência da preferência clara entre
alimentos mais e menos calóricos pelos machos e pela preferência por alimentos menos
calóricos pelas fêmeas. Uma possível explicação para isso estaria no fato de que C. jacchus sendo espécie gomívora9insetívora (Garber, 1992; Sussman & Kinzey, 1984), diferentemente de primatas frugívoros, não utiliza carboidratos solúveis (principal fonte
energética presente nos frutos) como principal recurso energético metabolizável. Desse
modo, C. jacchus poderia não apresentar limiar sensorial capaz de detectar diferenças sutis de concentração entre os carboidratos solúveis presentes em diferentes frutos de
modo eficiente.
De fato, o grau de frugivoria de uma espécie pode afetar sua habilidade em
discriminar diferentes intensidades de açúcares presentes nos alimentos (Laska et al., 1999). Tal dificuldade em detectar as concentrações entre os carboidratos solúveis
para o G2. No entanto, durante a Experimentação 1, as fêmeas exibiram preferência
pelos alimentos menos energéticos. É possível que elas, diferentemente dos machos,
possam ter sido influenciadas, inicialmente, por aspectos mecânicos para tomar sua
decisão. Foram observadas, por exemplo, situações em que as fêmeas apenas mordiam
parte da maçã e não a consumiam. Esse comportamento poderia ter sido realizado com
o intuito de analisar a resistência à punção do fruto. Já foi observado que macacos
aranha (Ateles paniscus) consomem frutos que apresentam resistência de punção do pericarpo menor do que os frutos que são consumidos pelos cuxiús (Chiropotes satanas); assim, o padrão de escolha do alimento é afetado pela rigidez da fruta entre essas duas espécies de primatas (Kinzey & Norconk, 1990). Além disso, observamos
que algumas fêmeas descartavam porções da manga ricas em fibras. Há trabalhos que
mostram que pistas táteis são utilizadas por primatas para analisar se alimento novo
necessita de alta demanda energética e investimento temporal para ser processado
(Laska et al., 2007). Assim, a textura pode ser um indicador do status nutricional do fruto. Além dos aspectos mecânicos e de características do alimento, as fêmeas não
sendo capazes de, inicialmente, selecionar o mais energético, podem ter utilizado a
familiaridade para selecionar os alimentos presentes no G2. Apesar de todos os
alimentos serem conhecidos pelos animais, não oferecemos a maçã e a manga com a
mesma frequência que o mamão e a melancia em nossa colônia.
Outro aspecto que observamos nas fêmeas com relação ao consumo de alimentos
presentes no G2, foi o fato de que com o decorrer das sessões elas exibiram gradual
mudança na preferência alimentar. Na Experimentação 1, as fêmeas exibiram
preferência por alimentos menos calóricos; já na Experimentação 2, elas não mostraram
preferência pelos diferentes alimentos que foram apresentados. Uma provável
associar as características dos alimentos com baixa frequência de oferta com os seus
efeitos pós9ingesta . De fato, já foi observado que o feedback pós9ingesta tem um papel importante na seleção de alimentos em alguns primatas, inclusive na seleção de
alimentos novos. Foi verificado, por exemplo, que após a familiarização com alimentos
novos, macacos prego utilizaram além do feedback sensorial, feedback pós9ingesta para selecionar alimentos mais calóricos (Visalberghi et al., 2003).
Após a discriminação qualitativa das recompensas por C. jacchus, analisamos o papel das mesmas no desempenho da discriminação visual simultânea. Observamos que
o desempenho tanto dos machos quanto das fêmeas de C. jacchus, em ambos os grupos, tendeu a aleatoriedade. Isso provavelmente deve9se ao fato de que, diferentemente do
G1, os animais não foram capazes de discriminar qualitativamente as diferentes
recompensas no G2. Devido a essa incapacidade de discriminação no G2, as diferentes
recompensas presentes nesse grupo apresentariam o mesmo valor para C. jacchus; consequentemente, nesse grupo, não foi observado remoção diferencial para nenhuma
das duas formas. Além disso, como as sessões do G2 foram intercaladas com as do G1,
é provável que o desempenho nesse último grupo tenha sido influenciado pelas
recompensas do G2. Devido a isso, provavelmente, os animais criaram falsa expectativa
a respeito do valor da recompensa que estava no triângulo; que por sua vez, levou a uma
maior remoção dessa forma no G1 (contribuindo para a aleatoriedade dentro desse
grupo, no que se refere à remoção da forma geométrica). Essa falsa expectativa foi
evidenciada pela ausência do consumo dos alimentos presentes no triângulo no G1, após
a remoção da forma (mostrando que o valor da recompensa presente ali não era o que o
animal esperava). De fato, já foi evidenciado que macaco rhesus (Macaca mulatta) durante a discriminação visual simultânea de objetos é capaz de estabelecer memória
permitindo o animal antecipar testes posteriores, o que, consequentemente, irá guiar a
tomada de decisão do animal (Murray & Gaffan, 2006).
Com base no que foi exposto, vimos que a recompensa exerce influência no
desempenho da aprendizagem discriminatória. Um fato que ilustra isso é que em testes
de resposta espacial com retardo, primatas apresentaram taxa de erro menor quando
recebiam recompensa mais preferida em detrimento da menos preferida; essa diferença
no desempenho, por sua vez, indicava que o primata criava expectativa sobre a
recompensa que estava por vir (Hassani et al., 2001). De maneira similar, após a resposta correta em tarefa com retardo, macacos mostraram hesitação e
“desapontamento” quando a recompensa foi substituída (sem o macaco perceber), pelo
experimentador, por outra menos preferida; isso, por sua vez, ilustra a evidência de
fatores representativos de aspectos qualitativos da recompensa (Tinklepaugh, 1928).
Além disso, em aves também já foi observado que a recompensa influencia a
aprendizagem discriminatória uma vez que em relação a recompensas menores pombos
aprendem discriminação de cores mais rápido quando recebem recompensas maiores
(Rose et al., 2009).
Concluindo, o trabalho mostrou que Callithrix jacchus utiliza o valor calórico como critério para selecionar alimentos. Entretanto, esses primatas não foram capazes
de fazer essa discriminação qualitativa quando os pares de alimentos apresentados
possuíam pequena diferença calórica. Nessa situação, o critério utilizado pelas fêmeas
para selecionar os diferentes alimentos apresentaram duas fases distintas: inicialmente
elas utilizaram informações mecânicas vindas dos mesmos e/ou a familiaridade;
de decisão (Doya, 2008). Além disso, observamos que aspectos qualitativos das
recompensas influenciam o desempenho da discriminação visual simultânea de objetos.
Agradecimentos Agradecemos aos funcionários do Núcleo de Primatologia da
Universidade Federal do Rio Grande do Norte (UFRN) por terem proporcionado
cuidados adequados aos animais que utilizamos nesse estudo. Somos gratos também aos
pesquisadores e chefes do Núcleo de Primatologia Daniel Marques de Almeida Pessoa e
Maria Teresa da Silva Mota pela concessão dos animais para a realização do projeto. A
CAPES pelo apoio financeiro que auxiliou na execução desse projeto. Aos
pesquisadores Wallisen Tadashi Hattori e Fellipe Nalon Castro pelos conselhos
estatísticos. A pesquisadora Maria de Fátima Arruda cujos conselhos auxiliaram no
aperfeiçoamento do presente trabalho. Agradecemos também aos revisores por seus
comentários perspicazes que contribuíram para melhorar o manuscrito original.
Referências bibliográficas
Alcock, J. (2001). Animal Behavior an Evolutionary Approach.Massachutts: Sinauer. Bachevalier, J., Brickson, M., Hagger, C., & Mishkin, M. (1990). Age and sex
differences in the effects of selective temporal lobe lesion on the formation of visual
discrimination habits in rhesus monkeys (Macaca mulatta). Behavioral &euroscience, 104,8859889.
Bayley, P. J., Franscino, J. C., & Squirre, L. R. (2005). Robust habit learning in the