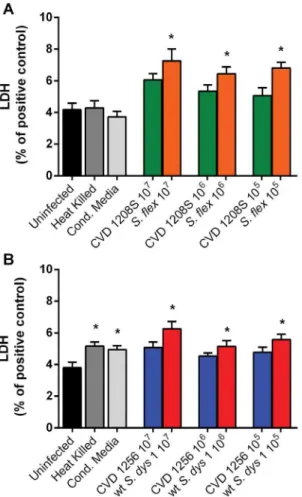
Effect of wild-type Shigella species and attenuated Shigella vaccine candidates on small intestinal barrier function, antigen trafficking, and cytokine release.
Texto
Imagem
Documentos relacionados
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
This log must identify the roles of any sub-investigator and the person(s) who will be delegated other study- related tasks; such as CRF/EDC entry. Any changes to
O caso do bairro de São Francisco, na cidade de Niterói, estado do Rio de Janeiro, trouxe conclusões e reflexões interessantes sobre que pontos levam a
The objective of this study was to evaluate the effect of oxidized soybean oils on the growth performance, metabolic oxidative status and intestinal barrier function of
A cross-sectional study adopting an ex post facto design was conducted with a non-probability sample of 4,759 students of both sexes with an average age of 18.4 years and using
En este trabajo, se valora la presencia de ideación suicida a partir de variables predictoras como la sintomatología depresiva, impulsividad y autoestima en adolescentes de
Considerando o cenário de crescimento da população idosa, as mudanças fisiológicas do processo de envelhecimento e as consequências da perda de peso para o estado de saúde, o
