Programa de Pós-graduação em Psicobiologia
Natal/RN 2007
Carolina Herédia Tosi SINCRONISMO COMPORTAMENTAL EM BOTO CINZA (Sotalia guianensis)
CAROLINA HERÉDIA TOSI
SINCRONISMO COMPORTAMENTAL EM DO BOTO-CINZA (Sotalia guianensis)
Dissertação apresentada ao programa de pós-graduação em Psicobiologia no Departamento de Fisiologia da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para a obtenção do título de Mestre.
Orientadora: Profa. Dra. Renata Gonçalves Ferreira
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Tosi, Carolina Herédia.
Sincronismo comportamental em Boto cinza (Sotalia guianensis) / Carolina Herédia Tosi. – Natal, RN, 2007.
103 f. : il.
Orientadora : Renata Gonçalves Ferreira.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1.Comportamento animal – Dissertação. 2. Sincronismo comportamental – Dissertação. 3. Boto cinza (Sotalia guianensis) - Dissertação. I. Ferreira, Renata Gonçalves. II. Título.
Título: Sincronismo comportamental em boto-cinza (Sotalia guianensis)
Autor: Carolina Herédia Tosi
Data: 28 de setembro de 2007
Banca Examinadora:
__________________________________________________________________ Prof. Dr. Paulo César Simões-Lopes (UFSC)
__________________________________________________________________ Prof. Dr. Flávio José de Lima Silva (DECB-UERN)
“Muito estudar não basta para ensinar a compreender”
AGRADECIMENTOS
A minha família que sempre me apoiou na busca de realizar meus sonhos (mesmo quando pareciam impossíveis!!) e que muito contribui para meu crescimento profissional e pessoal. Valeu Grande Família, amo vocês!!
Ao meu amor Fagner Magalhães por ter me apresentado o mundo dos cetáceos e por fazer parte da minha vida. Obrigada por toda atenção, amor e cuidado dedicados e por estar sempre comigo, caminhando juntos em busca de novas conquistas. Te amo muito!!!
A Prof. Dra. Renata Gonçalves Ferreira pela orientação e compreensão em momentos difíceis e de decisão ao longo do meu mestrado. Obrigada pela dedicação, principalmente nesta fase final do nosso trabalho!!
Ao Projeto Pequenos Cetáceos (PPC) pelo suporte logístico antes e durante minha coleta de dados e por estar me proporcionando novas experiências profissionais. A amiga mais brasileira do que argentina Rosana Garri, que se tornou mais do que uma “hermana”, longe de casa somos uma a família da outra, juntas desde o início nas horas boas e ruins (e olha que não foram poucas!!), sempre unidas lutando por um mesmo ideal. Obrigada por todo carinho e amizade, e nunca esqueça que sou sua amiga-filha hein!!!
A alguns amigos que contribuíram para a realização das minhas coleta de dados, ofereço meu carinho e agradecimentos mais sinceros, a Kelly Pansard, Bruno Maggi, Rosana Garri, Liisa Havukainen, Karla e Bruno Colaço, ao amigo colombiano Maurício Carrasquilla e em especial aos amigos Emmanuel Moralez (Manú) pela companhia e injeções de ânimo nas intermináveis horas de ponto e a Fernando Roberto e sua filha Rafaela pela amizade de terem ficado “boiando” horas no mar como parte da minha coleta.
Ao amigo Lídio França, pelo carinho, atenção e amizade durante toda minha jornada, por estar sempre disponível a ajudar e a quem devo grande parte do meu conhecimento em campo. Valeu por tudo!!!
Ao piloto Zé e a Seu Zé Vicente (Pipa) pela grande simpatia e boa vontade, disponibilizando sua lancha para a finalização das minhas coletas.
Aos amigos de turma, e em especial a Marina Dal Poggetto e Fernanda Cutrim pelo companheirismo e pelas bebidas e comilanças periódicas (vou sentir saudades!!), a Dell, Thomas, Diego, Garcia e Ricardo pela nossa inesquecível semana em Nísia (rimos muito!!!)
A Paul e Michele, amigos queridos, obrigada por tudo!! Se não fosse por vocês eu seria uma sem teto em Natal!!
A João Carlos Borges e Jociere Vergara que me impulsionaram para o mestrado, obrigada pela confiança!!
SUMÁRIO
Páginas RESUMO
ABSTRACT INTRODUÇÃO Vida em grupo
Sincronismo de atividades (estado comportamental) Fatores que influenciam o sincronismo de atividades Sincronismo de movimento (evento comportamental) Custos e benefícios do sincronismo
Sincronismo em cetáceos
Efeito das embarcações sobre os cetáceos
ix x 1 2 4 5 8 9 10 13 OBJETIVOS Objetivo geral Objetivos específicos 17 18 18 METODOLOGIA GERAL
Espécie em estudo
Descrição da área de estudo Coleta de dados
Definições adotadas Procedimento de análise Análise estatística 21 22 23 24 28 30 35 RESULTADOS E DISCUSSÃO
1) Determinação do padrão comportamental dos botos cinza 1.1. Resultados
1.2. Discussão
2) Influência de curto prazo das embarcações no comportamento geral e no sincronismo dos golfinhos
2.1. Resultados 2.2. Discussão
3) Caracterização da dinâmica ecológica do sincronismo comportamental (sincronismo de atividades) dos golfinhos
3.1. Resultados 3.2. Discussão
4) Caracterização da dinâmica social do sincronismo comportamental (sincronismo de respiração e eventos) dos golfinhos
RESUMO
Sincronismo é a efetivação ao mesmo tempo de um determinado comportamento por dois ou mais indivíduos. Nos cetáceos, os comportamentos realizados de forma sincrônica são pouco estudados, e somente a partir de 2000 verifica-se um maior esforço na cetologia para investigação deste sincronismo comportamental. O sincronismo em cetáceos tem sido relatado para uma variedade de contextos incluindo alimentação, socialização, cuidada parental, enquanto descansam, viajam e brincam, durante a afiliação, no comportamento sexual e possivelmente enquanto ensinam. O sincronismo de comportamento é influenciado por variáveis ambientais e pode modificar-se em resposta a situações estressantes como o tráfego de embarcações. Este estudo visa caracterizar o sincronismo de comportamento efetivado pelo boto cinza (Sotalia guianensis) na praia de Pipa/RN. Para tanto foram
realizadas um total de 414h de observação durante o ano de 2006, com a varredura como método de registro. Para a análise algumas etapas e objetivos específicos foram determinados. Primeiramente, foi determinado o padrão de atividades dos grupos que freqüentaram as enseadas na época do estudo, verificando possíveis diferenças correlacionadas à enseada e ao período do ano. As análises mostraram que ocorreu maior forrageio no período após as chuvas, mas não houve modificação no número de indivíduos ou coesão entre os mesmos. Em seguida, analisamos a influência do turismo de observação de golfinho sobre o comportamento geral (orçamento de atividades e coesão entre os indivíduos) e sincrônico dos animais. Foi verificado que não houve influência do barco no comportamento dos golfinhos. Por fim, testamos sete hipóteses acerca da dinâmica ecológica e social do sincronismo comportamental. Destacou-se a influência de fatores externos (ambientais) na apresentação do sincronismo, a ausência de influência da faixa etária, e a exibição mais seletiva (individualizada) de respiração sincrônica em forrageio e socialização, dado o menor número de indivíduos que apresentam este sincronismo respiratório.
ABSTRACT
Synchronism is the efetivation of the same behavior at the same time by two or more individuals. In cetology only from 2000 we verify an increasing of efforts for studies of synchronic behaviors. The synchronism in cetaceans was reported for a variety of contexts, including feeding, socializing, parental care, while resting, traveling and play, during afilliation, in sexual behaviour and possible while teaching. The synchronism in behavior is influenced by environmental factors and can be modified during stress situations such as the presence of boats. The aim of this study is to explore behavior synchronism presented by the estuarine dolphin (Sotalia guianensis) in Pipa, Rio Grande do Norte state, Northeastern Brazil. A total of 414
hours of observation during the year 2006, using scan sampling method, from a land based sighting was conducted. Analyses were conducted in four steps. Firstly, we determined the pattern of activity of the groups within the bay, testing for the influence of bay configuration before and after rains periods on the activity of animals. Analyses show that forage occurs more frequently after the rainy period, but there was no change in the number of individuals or spatial cohesions between them. Secondly, we analysed the influence of turism boat traffic on the behavior of animals. We verified no changes on the bahavior of animals during the presence of boats. Finally, we tested seven hypotheses regarding the ecological and social dynamics of synchronic behavior. Environmental aspects were considered the main influence for the synchronism, no influences of age, and more selective exihibition of synchronic breathing in feeding and social displays were observed during the study period.
INTRODUÇÃO
Vida em Grupo
Uma pressão seletiva é uma condição particular do ambiente que beneficia indivíduos com certas características (benefícios medidos em termos de fitness ou
propagação gênica), mas que prejudica (dificulta a propagação gênica) aqueles sem tal característica. Assim, segundo a perspectiva evolutiva, comportamentos em geral, e a vida em grupo, mais especificamente, são vistos como adaptações em resposta a características particulares do ambiente (Byrne, 1995).
Diversas vantagens para a vida em grupo já foram enumeradas. A vantagem mais discutida para a vida em grupo é o incremento da proteção contra predadores (Hamilton, 1971; Alcock, 1993; Krebs & Davies, 1996). Este incremento pode ocorrer de quatro formas. A primeira consiste na habilidade de detectar predadores: vários olhos são melhores do que um. Por exemplo, no estudo de Bertram (1980,
apud Krebs & Davies, 1996) em bandos de avestruzes, o autor constatou que
quando num grupo, cada indivíduo gastava uma proporção menor do seu tempo vigiando do que quando sozinho. Ademais, a vigilância total do grupo (proporção do tempo com pelo menos uma ave vigiando) aumentava ligeiramente com o tamanho do grupo. Portanto, cada ave do bando tem mais tempo para se alimentar e desfrutar de maior percepção quanto a aproximação de predadores. Os avestruzes levantam suas cabeças a intervalos aleatórios, o que torna impossível para um predador prever quanto tempo ele tem para avançar sem ser detectado por sua vítima.
Já no efeito de confusão, os indivíduos posicionados no meio de um grupo podem estar mais seguros do que aqueles da borda. Segundo Hamilton (1971), se o predador captura as vítimas das extremidades, cada membro do grupo deveria procurar uma posição central, escondendo-se atrás de outros. Há evidências de que os predadores sofrem um tipo de confusão quando atacam um grupo denso de presas (Neill & Cullen, 1974) e isto pode fornecer uma explicação do por quê os predadores direcionam os ataques para a borda do grupo. Uma quarta proteção contra predadores é a defesa cooperativa. Indivíduos de um mesmo grupo podem atacar conjuntamente um predador, como é o caso do mobbing em pássaros (Lee,
1994).
Se somarmos estes quatro efeitos, pode ser vantajoso para o predador concentrar seus ataques a grupos pequenos e aos indivíduos solitários. Sendo assim, não apenas viver em grupo é vantajoso, mas o tamanho e a posição do animal dentro deste também influenciam (Alcock, 1993).
Outra vantagem seletiva da vida em grupo pode ser a colaboração na exploração de recurso alimentar, seja por aumento da probabilidade de encontrar ou capturar alimento, seja na defesa cooperativa de fontes alimentares. Segundo Krebs & Davies (1996), para espécies sociais que se alimentam de sementes e frutos, as chances de encontrarem bons locais para alimentação são muito maiores do que um indivíduo solitário, e uma vez que o lugar tenha sido encontrado, haverá abundância de alimento pelo menos por algum tempo. Em se tratando de predadores que capturam presas móveis, indivíduos em um grupo podem ser capazes de capturar uma presa difícil para um indivíduo sozinho subjugar, ou porque a presa é grande demais, ou porque é muito ágil. Quando as presas estão agrupadas, os predadores que caçam coletivamente podem ser melhor sucedidos em isolar uma vítima de seus companheiros e, subsequentemente, persegui-la até subjugá-la.
Lee (1994) discute ainda outras vantagens da vida em grupo, tais como: cuidado com a prole (ex. aves), termorregulação (ex. pingüins e morcegos) e troca de informação (ex. primatas).
na competição por recursos, tanto para fins reprodutivos quanto alimentares. Esta pode apresentar-se sob a forma de competição direta (conflitos e agressões) ou indireta (acesso anterior ao recurso). Um exemplo de competição indireta é verificado em peixes, onde os indivíduos na frente do cardume capturam as presas privando aqueles da retaguarda (Alcock, 1993; Krebs & Davies, 1996).
Uma segunda desvantagem seria o aumento da evidência em relação a predadores, tendo em vista que a detectabilidade de animais em grupos por predadores é maior do que a de indivíduos solitários. Outros custos seriam relativos ao aumento de doenças e transmissão parasitária como resultado da proximidade. O risco da infidelidade com vizinhos (muito comum em aves) e o canibalismo, também podem ser citados. (Hamilton,1971; Taylor,1979; Wrangham et al., 1993;
Lee, 1994; Côté & Poulin,1995; Valenzuela et al., 2000).
Outras duas dificuldades podem ainda ser destacadas: manter o contato com outros para permanecer integrado com o grupo, e estabelecer coordenações físicas e sincronismo com outros membros (Lammers et al., 2003).
Sincronismo de atividades (estado comportamental)
Apesar dos custos e benefícios da vida em grupo serem extensivamente investigados (Pulliam & Caraco, 1984; Emlen, 1991), relativamente pouca atenção tem sido dada à necessidade dos membros dos grupos para sincronizarem suas atividades, e aos custos proximais que isto representa em termos de orçamento de atividades (Conradt, 1997 e 1998).
atividades (Cotê et al., 1997; Conradt & Roper, 2000; Ruckstuhl & Neuhaus, 2001;
Boyd & Bandi, 2002; Michelena et al., 2006).
O sincronismo, entretanto, possui certa complexidade pois, segundo Conradt & Roper (2000) para que a sincronização ocorra é preciso que os membros do grupo permaneçam no mesmo local e na mesma hora. Um possível mecanismo para o sincronismo é a facilitação social. Neste caso, o comportamento de um indivíduo pode induzir o mesmo comportamento em outros, podendo desta forma gerar o sincronismo, aumentando os benefícios da vida em grupo e permitindo que este fique mais coeso. Coté et al. (1997) usam como suporte à esta sugestão o fato de
que o sincronismo é maior no início de um surto de atividades que no final (i.e. ocorreria facilitação social para iniciar a atividade, mas a interrupção desta dependeria de variáveis internas individuais).
O sincronismo resultante da facilitação social é uma fina escala do sincronismo resultante dos estímulos ambientais e deste modo aumenta a coesão de grupos sociais (Clayton, 1978). Michelena (2006) em um experimento com ovelhas, verificou que o nível de sincronia observado entre grupos ocupando áreas adjacentes mas que não tinham contato visual foi próximo de zero, enquanto o nível de sincronia entre grupos com contato visual foi alto.
Fatores que Influenciam o sincronismo de atividades
- Variáveis Ambientais
diferentes áreas, que ocorreu devido a um mesmo fator ambiental (Schaub et al.,
2005).
Fatores externos podem também levar a sincronismos de história de vida. Para organismos que apresentam um ciclo de vida complexo, as pressões de predação podem levar ao sincronismo em períodos de vida vulneráveis. Por exemplo, o sincronismo na reprodução (peixes, anfíbios, répteis, aves e mamíferos) e na dispersão larval, que pode ocorrer em invertebrados marinhos, relacionam-se aos benefícios antipredatórios (De Vito, 2003).
Estes fatores ambientais, entretanto, tendem a influenciar todos os organismos de uma determinada população ou ter moldado os representantes das espécies (em nível distal) de uma forma mais ou menos homogênea. Existem, por outro lado, variações entre os indivíduos que influenciam a dinâmica do comportamento sincrônico num nível mais proximal, e a dificuldade em sincronizar torna-se maior quão mais díspares forem os componentes do grupo social.
Variáveis Internas
- Tamanho corpóreo e idade
A dinâmica fisiológica dos indivíduos (ex. padrão hormonal, necessidades alimentares e sociais) varia no decorrer da história de vida. Uma importante variável que influencia o sincronismo dos indivíduos é a faixa etária. Estudos em diversos mamíferos (primatas e ungulados principalmente) mostram que indivíduos de mesma faixa etária tendem a apresentar determinados comportamentos na mesma hora do dia. Observações em grupos de Saguinus sp., Cebus sp. e outros
primatas mostram que infantes e juvenis tendem a ter longos surtos de brincadeiras, enquanto os adultos dormem ou forrageiam (Lee, 1983; Smuts et al., 1987; Boinsky
& Garber, 2002). Em ungulados, grupos de subadultos tendem a sincronizar e otimizar seu orçamento de atividades, alternando entre surtos de alimentação e surtos de interações competitivas (Gross et al., 1995; Cransac et al., 1998). Este é
Ruckstuhl (1999) e Ruckstuhl & Festa-Bianchet (2001) destacam que a dificuldade de sincronização devido ao tamanho corpóreo não se deve somente à idade. A segregação por sexo tem sido observada em algumas espécies de ungulados sociais. Conradt (1998) sugere que a sincronização de atividades é particularmente custosa em grupos que consistem de membros dimórficos. Mais especificamente, ela sugere que é freqüente a ocorrência de segregação social intersexual em espécies sexualmente dimórficas (e.g. ungulados, Clutton-Brock et al., 1982, macrópodes, Newsome,1980, primatas, Boinski, 1994, pinípedes, Kovacs et al., 1990, cetáceos, Brown et al., 1995 e proboscídea, Sukamar & Gadgil, 1988).
Segundo a autora, isto pode ser explicado pelo fato de sexos diferentes terem ritmos de atividades distintos (incompatibilidade em orçamento de atividades), causando uma maior freqüência de separação em grupos compostos por indivíduos de ambos os sexos do que em grupos homogêneos (Ruckstuhl, 1998). Em algumas espécies de ungulados, como também em macacos e baleias que vivem em grupos, a segregação sexual entre machos e fêmeas é bastante comum (Ruckstuhl & Neuhaus, 2000).
Conradt & Roper (2000) constataram que se os custos para sincronizar as atividades começam a ser altos para uma categoria de idade e sexo específica, então o grupo se separa. De fato, animais em grupos compostos por mesmo tamanho corporal são realmente mais sincronizados em suas atividades e mais coesos do que indivíduos em grupos de tamanho corporal diferente (Ruckstuhl, 1998 e 1999). - Estado reprodutivo
alo-amamentação (Maestripieri, 1994; Manson, 1999). Por outro lado, Barrett et al.
(2000) discutem que, devido à falta de sincronia reprodutiva observada entre fêmeas de babuínos (Theropithecus gelada), as fêmeas preferem os machos como
parceiros de catação e aliança do que outras fêmeas, pois estas não seriam aliadas confiáveis (“reliable partners”) durante os períodos de amamentação e prenhez.
Sincronismo de movimento (evento comportamental)
No homem, o sincronismo pode aparecer como uma coordenação social, a qual é caracterizada por comportamentos verbais e não verbais, seguindo uma homogeneidade, onde os indivíduos dão, recebem e direcionam a atenção para um objeto ou para uma atividade específica, a fim de evitar a confusão com movimentos simultâneos, mas diferentes (Fellner, 2000). Esta coordenação é bastante observada em interações entre pais e filhos, ocorrendo com uma maior freqüência na infância (Feldman & Greenbaum, 1997), facilitando as trocas verbais e servindo como uma ferramenta de comunicação não verbal dos sinais de interesse do próprio indivíduo (Bernieri et al., 1988).
Recentemente, Hagen & Bryant (2003) sugeriam que o sincronismo em músicas e danças ritualísticas em grupos humanos servem como um sinal de cooperação e coordenação entre os indivíduos. Isto funcionaria como um sinal de aliança não apenas para os indivíduos do mesmo grupo, quanto para os indivíduos de grupos competidores. Ou seja, a sincronia na dança sinalizaria uma maior capacidade competitiva.
Em outros animais o sincronismo de movimento, apesar de pouco descrito, também tem forte componente social. Machos que precisam exibir-se para que as fêmeas os escolham, freqüentemente exibem comportamentos sincrônicos em áreas específicas, como é o caso da emissão luminosa sincrônica observada em vaga-lumes, coros de vocalizações de sapos, exibições em leks de pássaros e disputas
O sincronismo de movimento também é relacionado a capacidades cognitivas e aprendizagem. Caldwell & Whiten (2004), em estudo com saguis (Callithrix jacchus) discutem que a imitação é a apresentação de movimentos
sincrônicos por dois indivíduos, e que para que esta faculdade ter evoluído é necessário haver antes um sincronismo de atividades entre os membros do grupos, num ambiente social de não agressividade. Neste sentido, o aspecto social do sincronismo estaria contido dentro do aspecto ecológico.
Custos e Benefícios do sincronismo
Com base no descrito acima, verifica-se que os custos da sincronização podem influenciar a decisão do indivíduo em permanecer no grupo, o que por conseqüência, pode afetar a estabilidade e a composição do grupo, comprometendo por fim sua organização social (Conradt & Roper 2000). Estes custos apresentam uma alta particularidade para cada indivíduo do grupo, pois incluem membros de classes diferentes (sexo, tamanho e idade), sendo assim, a distribuição ótima do tempo de várias atividades é possivelmente distinta entre tais classes (Gompper, 1996 apud Conradt & Roper, 2000). Desta maneira, a sincronização de atividades
comportamentais é potencialmente custosa, pois requer que o indivíduo do grupo concilie seu próprio orçamento de atividades para igualar-se ao comportamento dos companheiros (Conradt & Roper, 2000).
Os benefícios, por sua vez, são mais investigados do que os custos. Em pelo menos dois filos, os artrópodes: insetos (Buck, 1988) e cordados: peixes (Breder, 1967; Partridge, 1982), aves (Greenberg, 2000) e mamíferos (Fellner, 2000; Ruckstuhl, 1999; Byrne, 2000), o comportamento sincrônico foi descrito e muitas funções deste comportamento podem ser propostas, estando todas relacionadas às vantagens da vida em grupo: o aumento da vigilância, a evitação de predadores e a comunicação (Fellner, 2000).
cooperativas. Grafe (2003) verificou que a vocalização sincronizada de sapos dificultava sua localização pelos predadores, funcionando também como um atrativo para as fêmeas, tratando-se de uma competição acústica para atrair o recurso (fêmea) reprodutivo. Segundo Ruckstuhl & Neuhaus (2001), o sincronismo de atividades é relativamente alto em grupos de forrageio, possivelmente para manter a coesão do grupo.
O ato da cooperação é um mecanismo que pode otimizar a energia investida de predadores individuais e as vantagens do sincronismo são mais evidentes nas estratégias de caça cooperativa. Gales et al. (2004) em seu estudo com foca
caranguejeira (Lobodon carcinophagus) observaram que as focas nadavam e
mergulhavam sincronicamente por períodos de tempo, providenciando evidências de benefícios mútuos (cooperação) em interações durante cada mergulho para capturar suas presas.
Em pingüins, o comportamento sincrônico com outros co-específicos também tem sido sugerido como uma função que aumenta a eficiência de forrageio (Ainley, 1972; Siegfried et al., 1975; Wilsonet al., 1986; Trembley & Cherel, 1999 apud Takahashi et al., 2004). De acordo com estudos de Takahashi et al. (2004),
pistas auditivas são importantes para os pingüins (Pygoscelis adeliae) aumentarem
o sincronismo no tempo de mergulho enquanto eles estão submersos no mar de gelo, pois além do risco de predação ser reduzido quando os indivíduos iniciam os mergulhos sincronicamente, através do efeito de diluição, aumentam a vigilância contra predadores.
Sincronismo em cetáceos
Apesar de anedoticamente bastante conhecido, atualmente estudos científicos relacionados ao sincronismo nos cetáceos são escassos, necessitando-se a intensificação de pesquisas voltadas para este comportamento específico.
Lammers & Au (2003) estudando direcionalidade nas vocalizações de golfinho rotador (Stenella longirostris) sugerem que esta tenha a função de
manter um indivíduo em contato com outros, como parte do desafio para permanecer integrado ao grupo. Outra função do sincronismo é proposta por Fellner (2000) como uma estratégia antipredatória, ajudando a proteger os golfinhos, através do efeito de diluição e confusão. Estes efeitos foram também mostrados por Hastie et al. (2003), onde grupos de golfinhos nariz de garrafa
(Tursiops truncatus) apresentavam uma respiração sincrônica em resposta a
presença de barcos, provavelmente por considerarem estes barcos como uma ameaça.
Analisando as atividades diárias dos animais, Fellner (2000), Hastie et al.
(2003) e Connor et al. (2006), observaram que o sincronismo tem sido usado em
uma variedade de contextos do comportamento nos cetáceos, incluindo alimentação, socialização, cuidado parental, enquanto viajam e brincam, em resposta a situações estressantes, durante a afiliação, no comportamento sexual e possivelmente enquanto ensinam.
À semelhança do que ocorre em cardumes de peixes, durante as viagens e deslocamentos, a sincronia em cetáceos pode ser utilizada para aumentar a eficiência hidrodinâmica dos grupos de cetáceos, resultando em uma maior conservação de energia, isto foi sugerido por Norris & Schlt (1988, apud Connor et al., 2006).
O comportamento realizado de forma sincrônica também pode funcionar como um agente que acentua a detecção de presa e predador. De acordo com Norris & Dohl (1980) os grupos de golfinhos movendo-se sincronicamente atuam de forma singular, onde os organismos são super sensitivos a um sistema de integração sensorial (SIS), e por ele ocorre a rápida transmissão de informações entre os membros do grupo, facilitando o reconhecimento do objeto de interesse (presa ou predador).
Simões-Lopes et al. (1998) estudando T. truncatus registrou a interação na
comportamental de predação entre cetáceos, mostrando que o sincronismo de comportamento também pode atuar como uma estratégia de caça cooperativa.
Gnone et al. (2001), por exemplo, apontam o sincronismo como um forte
indicativo do comportamento de descanso, os autores ainda sugerem que o sincronismo tem a função de conservar a energia e manter a vigilância nos grupos de golfinhos.
O sincronismo em golfinhos também está presente em contexto social. Conforme Mann et al. (2000), animais que vivem em grupo como os golfinhos,
expressam afiliação por proximidade, contato físico e movimentos sincrônicos. Sendo assim, a autora sugere que, dentro do contexto social, esta pode ser uma das mais importantes funções do sincronismo de movimento, funcionando também como um método de anunciar ou manter a relação hierárquica dentro do grupo.
Mann & Smuts (1999) e Fellner (2000) relacionam o sincronismo ao cuidado parental. O filhote no seu primeiro mês de vida apresenta um elevado sincronismo respiratório com sua mãe e permanece na maior parte do tempo em uma posição chamada de “echelon”. Este posicionamento e o sincronismo atuam como uma vantagem hidrodinâmica para os filhotes, pois estes são carregados ou impulsionados pelas pressões de ondas criadas pelo deslocamento de suas mães. A partir do segundo mês de vida este sincronismo começa a decair.
A função social do sincronismo é mais detalhada nos trabalhos de Connor et al. (2001; 2005; 2006; 2007). Este autor sugere que o sincronismo de respiração
tenha uma função importante nas interações afiliativas, principalmente nas alianças de primeira ordem entre machos. Nestas, foi verificado uma alta freqüência de respiração sincrônica quando havia presença de fêmeas ou de outra aliança de primeira ordem. Mais especificamente, Connor et al. (2006) sugere que o
melhor aliança durante situações de conflito. Neste caso, o sincronismo de respiração poderia ser também observado em outras situações, como na caça-cooperativa (como é o caso das focas caranguejeiras e pingüins acima citado).
Por fim, o sincronismo em cetáceos também pode atuar como um mecanismo de aprendizagem para indivíduos imaturos, provavelmente influenciando a comunicação através da imitação. Isto foi observado por Tayler & Saayman (1973) em cativeiro, onde golfinhos nariz de garrafa imitavam espontaneamente coespecíficos.
Cabe ressaltar, portanto, que no sincronismo de atividades pode ou não ocorrer sincronismo de movimento e respiração (por exemplo, grupos com mais de 100 indivíduos em deslocamento ou forrageio, mas não necessariamente em movimentos perfeitamente sincrônicos ou em respiração sincrônica). A exceção refere-se aos estudos sobre socialização, em que o sincronismo de respiração e movimento é contabilizado. Desta forma, estudos do sincronismo de atividades referem-se a uma abordagem mais ecológica, acerca das vantagens da vida em grupo, vantagens hidrodinâmicas, estratégias de caça cooperativa, ou competição entre indivíduos. Já os estudos sobre sincronismo de respiração e movimento voltam-se para uma análise de comunicação social e cognição.
Efeito das embarcações sobre os cetáceos
Muitas populações costeiras de cetáceos têm áreas de uso próximas a centros urbanos, por exemplo, golfinho nariz de garrafa (Tursiops aduncus) que ocorre em
Shark bay, Austrália; T. truncatus em Sarasota bay na Flórida; golfinho de hector (Cephalorhynchus hectori) em Porpoise bay, Nova Zelândia; golfinho de flancos
brancos do atlântico (Lagenorhynchus acutus) na Nova Inglaterra e S. longirostris
na ilha de Fernando de Noronha, Brasil. A ampla variedade de atividades humanas realizadas nestas áreas podem resultar em sérias injurias e mortalidade aos cetáceos (Hastie et al., 2003; Van Parijs & Corkeron, 2001), como por exemplo atividades
Embora nem todas as atividades antropogênicas causem impactos adversos, os veículos motorizados, em particular, são potencialmente uma fonte de perturbação principalmente para os cetáceos costeiros (Lemon et al., 2006;
Nowaceket al., 2001).
As baleias e golfinhos são animais carismáticos e que sempre atraíram a curiosidade humana. Em virtude disto, sua popularidade tem resultado em um rápido crescimento do turismo de avistagem (Whale and Dolphin-Watching) por todo mundo. Na estimativa mais recente, com dados até 1998, Hoyt (2001) encontrou que o turismo marinho de avistagem de cetáceos tem crescido consideravelmente nos últimos 40 anos e está presente em pelo menos 87 países. Esta indústria gera em torno de um bilhão de dólares por ano, transformando-se em um grande potencial econômico, e servindo de plataforma para a pesquisa e para fins educativos. Os benefícios do turismo baseado na natureza incluem trocas potenciais na atitude das pessoas em relação ao ambiente (Orams, 1995) e geram melhorias consideráveis para as comunidades locais (Duffus & Dearden, 1993; Hoyt, 2001). Porém um dos grandes desafios do turismo marinho é a proteção e conservação do habitat e da vida selvagem enquanto se administram as necessidades do turista (Giannecchini, 1993).
Apesar do inegável benefício econômico e educacional e do seu acelerado crescimento, pouco se conhece sobre os efeitos do turismo de observação no comportamento dos cetáceos (Constantine et al., 2004), principalmente aqueles
causados pelas embarcações motorizadas, que são muito utilizadas pela indústria do turismo. Esforços estão sendo direcionados para a quantificação dos potenciais impactos de embarcações turísticas na população de cetáceos de vida livre tanto a curto, quanto a longo prazo.
Efeitos diretos como injúrias físicas são fáceis de identificar, porém a maior dificuldade está na avaliação dos efeitos indiretos, como o impacto do contato acústico ou visual e no potencial reprodutivo dos indivíduos (Nowacek et al.,
- evitação da embarcação (Watkins, 1986; Janik & Thompson, 1996; Moore & Clarke, 2002),
- aumento na velocidade de natação (Nowacek et al., 2001; Moore &
Clarke, 2002; Jahoda et al., 2003),
- mudança na direção da natação (Nowacek et al., 2001; Lemon et al.,
2006),
- alteração na composição do grupo (Bejder et al., 1999),
- diminuição da distância inter animal (Nowacek et al., 2001),
- alteração do padrão respiratório (Janik & Thompson, 1996; Jahoda et al.,
2003),
- mudanças na vocalização (Lesage et al., 1999),
- alterações do padrão de atividades de superfície (Janik & Thompson, 1996; Richardson & Würsig, 1997)
- alteração no sincronismo na respiração (Hastie et al., 2003).
Hastieet al., (2003), buscou quantificar o padrão de respiração sincrônica de
uma população de T. truncatus na Escócia. Os autores investigaram possíveis
funções do sincronismo referentes a padrões de alimentação e presença de filhotes. Também foi investigado se o tráfego de embarcações afetava a respiração sincrônica dos golfinhos. Os resultados apontaram que a maioria dos golfinhos apresentou um padrão respiratório randômico e que apenas um terço dos grupos amostrados mostrou um sincronismo respiratório. Não foi verificada nenhuma alteração no comportamento referente a atividades de forrageio, entretanto, a sincronia foi negativamente relacionada à presença de filhotes no grupo e positivamente relacionada à presença de embarcações na área de estudo.
É importante também salientar que nem todas as mudanças comportamentais são negativas e que espécies diferentes não reagem da mesma maneira quando expostas ao mesmo impacto (Bejder et al., 1999). O comportamento chamado de
um menor gasto de energia metabólica, na maior parte do tempo sua cauda mantém-se imóvel e o golfinho corrige a direção apenas com movimentos de cabeça e nadadeiras peitorais” (Simões-Lopes, 2005). Este comportamento é freqüentemente observado em T. truncatus (William et al., 1992).
*
No Brasil, trabalhos voltados para o sincronismo comportamental de mamíferos são escassos. Estudos sobre o sincronismo em cetáceos, e em específico para a espécie boto cinza (Sotalia guianensis) são inexistentes. Sendo assim, o
objetivo desta pesquisa é analisar a dinâmica do sincronismo de comportamento realizado por grupos de S. guianensis que freqüentam as águas costeiras do litoral
OBJETIVOS
Objetivo geral
Caracterizar o sincronismo de comportamento realizado pelo boto cinza (Sotalia guianensis) na praia de Pipa/RN, explorando sua dinâmica ecológica e
social.
Objetivos específicos:
1) Determinar o padrão comportamental dos botos cinza, verificando possíveis diferenças comportamentais quanto às enseadas ocupadas e o período do ano (período antes das chuvas- período 1 e período após as chuvas- período 2). A população alvo deste estudo foi caracterizada quanto:
- ao padrão de atividade diária (i.e. orçamento de atividades);
- ao número e faixa etária dos indivíduos presentes na área de estudo; - a distância entre os indivíduos (Índice de coesão).
2) Analisar possíveis influências de curto prazo das embarcações no comportamento geral e no sincronismo dos golfinhos. Isto foi verificado quanto: - ao orçamento de atividades;
- à distância existente entre os indivíduos; - ao sincronismo de atividades;
- à respiração sincrônica.
3) Caracterizar a dinâmica ecológica do sincronismo comportamental (sincronismo de atividades) dos golfinhos. Para tanto foi verificado:
- o estado comportamental em que ocorre a maior porcentagem de sincronismo (freqüência de sincronismo);
- a duração do sincronismo nos estados comportamentais;
- o número de indivíduos sincrônicos por estado comportamental;
- a distância existente entre os golfinhos em sincronismo de atividade (índice de sincronismo);
Com relação à dinâmica ecológica do sincronismo testou-se as seguintes hipóteses:
Hipótese 1: Fatores externos (ex. presença de presas ou proteção na área, sazonalidade) tem ação preponderante no sincronismo de comportamento dos indivíduos
Predição: Uma grande proporção de animais presentes na enseada devem efetuar o mesmo comportamento ao mesmo tempo, e estes comportamentos devem variar segundo as condições ambientais locais. Inversamente, se fatores externos não são influência preponderante na efetivação dos comportamentos, então deve haver uma baixa proporção de indivíduos realizando o mesmo comportamento ao mesmo tempo, isto é, os animais devem diferir no comportamento apresentado, seguindo seu próprio orçamento de atividades.
Hipótese 2: O sincronismo de atividades facilita a coesão do grupo.
Predição: O sincronismo será maior em grupos de indivíduos com menor distância entre si. Inversamente, se o sincronismo não facilita a coesão do grupo, então o sincronismo de atividades irá manter o mesmo valor independente da distância entre os indivíduos.
Hipótese 3: Similaridade de necessidades energéticas aumentam o sincronismo de atividades.
Predição: Grupos compostos somente por adultos terão maior índice de sincronismo que grupos formados por adultos e imaturos.
Hipótese 4: O sincronismo de atividades tem função de cuidado e ensino.
4) Caracterizar a dinâmica social do sincronismo comportamental (sincronismo de respiração e eventos) dos golfinhos quanto a:
- o estado comportamental em que ocorreu o maior número de respiração sincrônica (sincronismo em respiração);
- faixa etária em sincronismo de respiração; - duração da respiração sincrônica;
- o número de indivíduos com respiração sincrônica; - simetria ou assimetria da respiração sincrônica; - freqüência de eventos sincrônicos.
Com relação à dinâmica social do sincronismo testou-se as seguintes hipóteses:
Hipótese 5: O sincronismo de respiração e eventos é uma demonstração social. Predição: Deve haver alta proporção de sincronismo de respiração e eventos durante o estado de socialização.
Hipótese 6: O sincronismo de respiração reflete necessidade de oxigênio para ações simultâneas.
Predição: Será observado um alto índice de sincronismo no estado de forrageio e deslocamento.
Hipótese 7: O sincronismo de respiração tem função de cuidado e ensino.
Espécie em estudo
Até recentemente dois ecótipos eram aceitos para a espécie Sotalia fluviatilis: um marinho, popularmente conhecido como boto cinza, e um fluvial,
conhecido como tucuxi. Porém, estudos recentes de Monteiro-Filho et al. (2002) e
Cunha et al. (2005), comprovam por análises morfométricas e genéticas que o
gênero Sotalia deve ser desmembrado em duas espécies. A espécie fluvial, Sotalia fluviatilis (Gervais, 1853), que é endêmica da bacia do rio Amazonas, ocorrendo
desde sua desembocadura até os rios Putumayo e Ucayalli no Peru (Borobia et al.,
1991; Silva & Best; 1994, 1996). E a espécie marinha, Sotalia guianensis (van
Bénéden, 1864) (figura 1), que apresenta uma distribuição tipicamente costeira e contínua na costa atlântica tropical e subtropical da América do Sul e Central cujos limites são: ao Sul, na baía norte de Santa Catarina (27°, 35’S; 49°, 34’W) (Simões Lopes, 1988) e, ao Norte, até Honduras (15°, 58’N; 85°, 42’W) (Silva & Best, 1996).
Figura 1. Boto cinza (Sotalia guianensis).
De acordo com Silva & Best (1996), apesar de relativamente pequeno, animais do gênero Sotalia são robustos e de nadadeiras grandes. Seu comprimento máximo
é de 2,06m e o comprimento médio é de 1,7m. Sua coloração varia, sendo cinza escuro na região dorsal e nas nadadeiras, e na região ventral a coloração é mais clara, chegando a ser rosada ou cinza claro.
S. guianensis é comumente encontrado em estuários e desembocaduras de
rios, ou baías próximas onde as águas são geralmente turvas (Borobia et al., 1991;
Flores, 1999; Cunha et al., 1999; Santos et al., 2000; Simão et al., 2000), porém no
nordeste do Brasil este golfinho também pode ocupar águas mais abertas ou mesmo oceânicas (Monteiro-Neto et al., 2004; Valle & Melo, 2006).
Segundo Silva & Best (1996), o boto cinza apresenta uma estrutura social caracterizada por grupos sociais pequenos (1-30 indivíduos), porém estes grupos são menores em áreas consideradas pobres em recurso alimentar, como é o caso da Baía de Guanabara (bastante poluída) no Rio de janeiro, onde o tamanho de grupo varia em torno de sete animais (Azevedo et al., 2005). Alguns trabalhos indicam que o tamanho e a estrutura do grupo variam de acordo com a hora do dia e o tipo de atividade (Silva & Best, 1996) e geralmente os golfinhos nadam em grupos coesos com os indivíduos muito próximos uns dos outros, o que sugere um estreito vínculo social para a espécie, principalmente entre fêmea e filhote (Hetzel & Lodi, 1993).
De acordo com IBAMA (2001), o boto cinza é uma espécie ainda pouco conhecida, cuja maioria dos dados biológicos foram coletados oportunisticamente ou coletados em áreas restritas. Grande parte dos estudos comportamentais para está espécie, tem focado em populações que habitam estuários, bocas de rios (e.g. Geise et al., 1999; Cremer et al., 2004; Azevedo et al., 2005) e baías protegidas
(Lodi, 2003; Daura-Jorge et al., 2004; 2005; 2007; Wedekin et al., 2007). Essa
limitada informação da história natural e dos dados demográficos ao longo da sua distribuição é que levou a espécie à classificação de “dados deficientes”.
Descrição da área de estudo
pela presença de um curral pesqueiro na enseada do Curral e por um número mais elevado de banhistas na enseada do Madeiro (observação direta).
Figura 2. Área de estudo próxima a desembocadura da lagoa de Guaraíras. Foto: Renato Matos
Figuras 3: Praia do Curral (a) e Praia do Madeiro (b). Fotos: Carolina Tosi
Coleta de dados
- Material
A coleta de dados contou com o auxílio de binóculos Bushnell 10x50, de um relógio digital e de duas planilhas padronizadas (ver anexos).
a b Enseada do Curral
Enseada do Madeiro
- Procedimento de coleta de dados
Os dados foram coletados a partir de dois pontos fixos, um localizado na enseada do Curral (1ª área) (figura 4) e o outro na enseada do Madeiro (2ª área) (figura 5). A coleta constou de seis horas de observações diárias no período de fevereiro a dezembro de 2006. As observações foram realizadas apenas no período seco, não tendo sido realizadas coletas em junho, julho e agosto, período em que as chuvas são mais intensas, o que prejudicava a visualização do observador em relação aos comportamentos que estavam sendo efetivados, a verificação do número de indivíduos presentes na área e a determinação da faixa etária destes golfinhos. A coleta de dados foi realizada por um pesquisador treinado, e eventualmente, por dois pesquisadores, permitindo checagem periódica da acuidade do observador principal.
Na enseada do Curral foi realizado um total de 26 dias de observação, sendo 20 dias efetivos de coletas, no período de fevereiro a maio, o que gerou 147,82 hs de esforço amostral e 47,21 hs de esforço efetivo, o que corresponde a 31,9% do tempo com os animais presentes na área. Na enseada do Madeiro foram realizados 50 dias de observação, sendo 13 dias de coleta nos meses de fevereiro a maio, e 37 dias de coleta nos meses de setembro a dezembro. O esforço amostral total na enseada do madeiro foi de 267,08 hs e o efetivo de 149,91 hs (56,1%), destas 74,38 hs de esforço amostral e 39,45 hs de esforço efetivo foi no período antes das chuvas (período 1), e 192,7 hs de esforço amostral e 110,46 hs de esforço efetivo foi no período após as chuvas (período 2).
A metodologia de coleta de dados foi varredura (scan sampling segundo Altmann, 1974), sendo realizados registros instantâneos a intervalos de tempo de 2 minutos. A cada intervalo de 2 minutos eram registrados em planilhas específicas1:
- o estado comportamental de cada indivíduo avistado dentro da enseada, definidos como: forrageio, deslocamento, socialização e descanso (ver definições adotas abaixo).
- número total de indivíduos na enseada;
- a faixa etária dos indivíduos (ver definições adotas abaixo); - a distância existente entre os indivíduos;
- se os indivíduos respiravam sincronicamente ou não; - se ocorreu algum evento comportamental sincrônico; - presença de embarcações.
Além disso, eventos comportamentais (i.e. comportamentos de curtíssima duração. Ver definições adotadas abaixo) foram registrados ad libitum. Como tanto
o tamanho do grupo quanto a área de observação eram pequenas, as freqüências dos eventos foram comparadas nos diferentes períodos assumindo-se a premissa de que tais “notas de campo” refletem a probabilidade de ocorrência dos eventos nos diferentes períodos e pelas diferentes classes etárias e de sexo (para uma discussão sobre análises com dados coletados ad libitum ver Lehner, 1996, pag. 196).
Para a coleta da distância existente entre os indivíduos, formulou-se uma planilha específica, onde a distância poderia ser anotada em quatro categorias: de 0 a 20 m, de 20 a 40m, de 40 a 80m e acima de 80m. As distâncias foram determinadas com parâmetros visuais (pontos específicos: bóias que delimitam a enseada) na área de estudo, e medidas com o auxílio de cordas metrificadas.
Para a verificação da influência das embarcações sobre o comportamento sincrônico dos indivíduos, foi anotado, independente dos intervalos de registro, o
1 Neste ponto cabe uma explicação metodológica. O estudo do comportamento por varreduras apresenta a
questão da dependência temporal entre os comportamentos, ou seja, a probabilidade de um comportamento ser registrado no instante t+1 depende do comportamento registrado no instante t. Em estudo anterior com uma população de S. guianensis em área 20 km mais ao sul da área do presente estudo, Queiroz (2006)
momento da entrada e da saída dos barcos na enseada, quantificando também a duração de permanência destes na área.
Figura 4: Ponto de observação na Praia do Curral.
Figura 5: Ponto de observação na Praia do Madeiro.
Ponto de observação
Foto: Carolina Tosi
Ponto de observação
Definições adotadas - Faixa etária
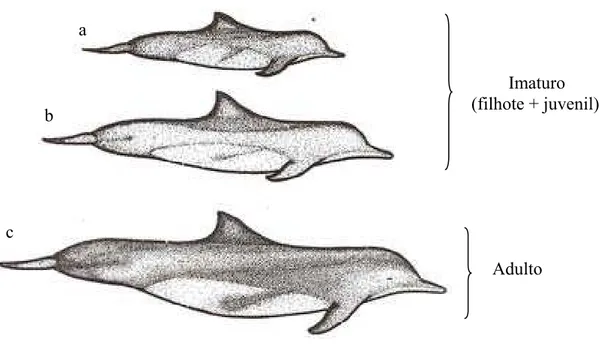
Adulto:Indivíduo de comprimento total com cerca de 2 m (figura 6).
Imaturo: É caracterizado pela soma de juvenis e filhotes, indivíduos estes com tamanho corporal inferior ao adulto (animais atingindo até 2/3 do tamanho do adulto), geralmente com coloração mais clara (figura 6).
Indeterminado: Esta categoria corresponde aos animais em que a faixa etária não pôde ser definida pelo pesquisador.
Figura 6. Determinação da faixa etária: a- filhote, b- juvenil, c- adulto. Ilustração: Lídio França
- Estados comportamentais
Forrageio
Natação sem direção fixa sendo periodicamente interrompida por eventos de perseguição, giros 360° (para definição consultar Nascimento, 2002) e bote na presa podendo este ter sucesso ou não (Norris & Dohl, 1980), podem estar inseridos neste estado, o padrão de varredura (procura de alimento), e de arrastão 1 e 2 (encurralamento das presas na enseada), conforme descrito por Nascimento (2002).
Adulto Imaturo (filhote + juvenil) a
b
Deslocamento
É caracterizado como a natação que apresenta uma direção definida resultando na mudança de posição do animal de uma determinada área para outra. De acordo com Nascimento (2002), a atividade pode ocorrer com velocidades distintas, podendo variar entre o deslocamento lento, rápido, em velocidade, com saltos (o que equivale ao porpoising) ou torpedos.
Socialização
Caracteriza-se por uma natação na superfície, entre dois ou mais indivíduos (adultos e/ou imaturos), e com constante contato físico, que pode incluir fricções, perseguições, elevação de partes do corpo, inspeção genital, agressão e displays. De acordo com a literatura a socialização pode envolver aspectos de brincadeira e cópula (Mann & Smuts, 1999; Nascimento, 2002; Lusseau, 2003; Constantine et al., 2004).
Descanso
De acordo com Gnone et al. (2001), Constantine et al. (2004) e Lusseau
(2003), durante a natação de descanso os indivíduos deslocam-se lentamente seguindo uma trajetória regular, subindo a superfície somente para respirar. Este comportamento pode ser realizado por um indivíduo solitário, mas freqüentemente dois ou mais animais tendem a descansar juntos, apresentando-se altamente agrupados (menos do que um corpo de distância entre eles), e nadando lado a lado. - Eventos comportamentais
Neste trabalho os eventos comportamentais utilizados foram: saltos, surf, espiar (cf. Simões-Lopes, 2005: spyhope), perseguição, giro 360º e bote. Para maiores definições consultar Nascimeto (2002) e Monteiro et al. (2006).
- Sincronismo de atividades
comportamental (Forrageio, Deslocamento, Socialização e Descanso) por dois ou mais indivíduos, independente da distância existente entre eles.
- Respiração sincrônica
A respiração sincrônica caracteriza-se pela subida a superfície para respirar realizada ao mesmo tempo (ou num intervalo de até 3 segundos) por dois ou mais indivíduos, que distavam até 20 metros entre si. Para ser considerada sincrônica, pelo menos dois eventos consecutivos de respiração simultânea devem ocorrer entre dois ou mais indivíduos e estes devem estar engajados no mesmo estado comportamental. Apenas um evento de respiração simultânea foi considerado como resultado do acaso.
-Sincronismo de eventos
Neste trabalho o sincronismo de eventos foi considerado como a realização simultânea de qualquer evento comportamental (saltos, surf, espiar, perseguição, giro 360°, bote), por indivíduos que distavam até 20 metros entre si e estavam engajados no mesmo estado comportamental.
Procedimento de análise
As análises foram realizadas utilizando-se dados diários padronizados. Esta padronização foi feita tanto a partir da media diária (para estados comportamentais e análises de sincronismo) ou da moda diária (para número de indivíduos).
1) Determinação do padrão comportamental dos botos cinza
- Orçamento de atividades
O orçamento de atividades foi analisado de uma forma geral (com todos os indivíduos presentes na área de estudo) e separando-se os momentos (registros) em que havia um grupo (2 indivíduos ou mais) -orçamento de atividades em grupo, dos momentos em que havia um indivíduo solitário nas enseada - orçamento de atividades de indivíduo solitário.
- Número de indivíduos na enseada
A análise do número geral de indivíduos dentro da enseada foi realizada determinando-se a moda, o número máximo e o número mínimo observado em todos os dias de coleta de dados. Estes valores foram organizados utilizando o programa excel, representando-se o número de registros em que um número determinado de indivíduos (1 ind; 2 ind; 3 ind, etc...) estavam presente. Além do valor total, foram encontrados os valores diários da moda, do número máximo e mínimo de indivíduos em cada estado comportamental (deslocamento, socialização, descanso e forrageio).
- Faixa etária
Para a análise da faixa etária, encontrou-se a porcentagem por registros de cada categoria etária (adulto, imaturo e indeterminado), ou seja, em cada intervalo de registro dividiu-se o número de indivíduos correspondente a cada categoria etária pelo número total de indivíduos registrados. Em seguida, a média diária destas proporções foi calculada.
Esta análise foi refinada, calculando-se a porcentagem média diária das faixas etárias considerando apenas o número de indivíduos engajados em cada um dos quatro estados comportamentais.
- Distância entre os indivíduos (Índice de coesão)
a 80m e acima de 80m) em cada intervalo de registro. Para o cálculo do índice foi feita uma divisão do número de indivíduos presente em cada categoria de distância pelo número total de indivíduos presentes em cada registro de observação. O índice de coesão consiste na média diária destes valores e representa a proximidade entre os indivíduos dentro da enseada, independente do sincronismo.
2) Influência de curto prazo das embarcações no comportamento geral e no sincronismo dos golfinhos
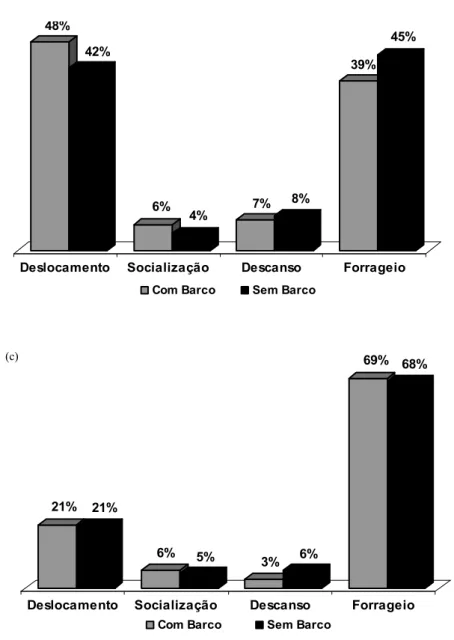
Para a segunda análise foram realizadas as mesmas análises descritas acima separando-se os dados com e sem barcos, em cada condição (enseada do curral, enseada do madeiro 1 e enseada do madeiro 2).
3) Caracterização da dinâmica ecológica do sincronismo comportamental (sincronismo de atividades) dos golfinhos
A análise do sincronismo foi realizada a partir de uma análise mais geral, sendo progressivamente refinada. Em todas as análises foram excluídas os registros em que havia apenas um individuo na área de estudo.
Foram realizadas cinco análises para caracterização do sincronismo de atividades:
- Freqüência de sincronismo nos estados comportamentais
mesmo tempo. Ao dividirmos os dois valores teremos o percentual de sincronismo dentro do total de cada atividade.
- Duração do sincronismo nos estados comportamentais
Para quantificar-se a duração do sincronismo em cada estado comportamental, contou-se o número de registros sem interrupção, em que dois ou mais indivíduos estavam em sincronia de atividade comportamental.
- Número de golfinhos em sincronismo de estado comportamental
O número de indivíduos que estavam sincrônicos em cada estado comportamental foi dividido pelo total de indivíduos presentes em cada registro. A média diária destas porcentagens foi calculada. Os intervalos de registro que apresentavam somente um indivíduo ou que apresentavam apenas um indivíduo por categoria comportamental foram descartados, para evitar que estes valores percentuais causassem uma interferência na análise da porcentagem média diária.
- Indivíduos sincrônicos por distância (índice de sincronismo)
Este cálculo foi semelhante ao realizado para o índice de coesão (descrito acima), sendo que apenas o número de indivíduos que apresentavam estados comportamentais sincrônicos foi considerado (chegando-se ao índice de sincronismo). O objetivo foi verificar se existe um maior sincronismo com o aumento da coesão entre os indivíduos.
- Faixa etária em sincronismo de estado comportamental
4) Caracterização da dinâmica social do sincronismo comportamental (sincronismo de respiração e eventos) dos golfinhos
Foram realizadas seis análises nesta etapa. Em todas as análises foram excluídos s registros em que havia apenas um individuo na área de estudo.
- Sincronismo em respiração
Nesta análise, foi atribuído o valor um (1) ao registro em que dois ou mais indivíduos estivessem engajados na mesma categoria comportamental (e distando até 20 metros entre si) em cada estado comportamental, ou valor zero (0) em caso contrário. Este total correspondeu às Possíveis Janelas Sincrônicas por estado comportamental. Em seguida foi atribuído valor 1 ao registro em que foi verificado, além do estado comportamental sincrônico, a respiração sincrônica, ou o valor zero caso contrário. Este total diário foi utilizado para verificar a proporção de respiração sincrônica dentro do sincronismo em cada estado comportamental por dia. Para se chegar à porcentagem de sincronismo em respiração, dividiram-se as janelas sincrônicas (registros em que ocorreu a respiração sincrônica) pelas possíveis janelas sincrônicas de cada categoria comportamental.
- Faixa etária em sincronismo de respiração
O cálculo da faixa etária em sincronismo de respiração é semelhante ao da faixa etária em sincronismo de atividade, para tal, utilizou-se três pares de categorias (AA-adulto e adulto, AI- adulto e imaturo e II- imaturo e imaturo) contando-se o número de vezes em que os indivíduos estavam agrupados em sincronia de atividade e com a respiração sincrônica.
- Número de golfinhos com respiração sincrônica
- Duração da respiração sincrônica
A quantificação da duração da respiração sincrônica é similar à da duração do sincronismo de atividade: em cada estado comportamental contou-se o número de registros sem interrupção, em que dois ou mais indivíduos estavam em sincronia de respiração.
- Simetria ou assimetria da respiração sincrônica
Utilizou-se um surto (seqüência) de registros em que havia dois ou mais indivíduos engajados no mesmo estado comportamental e em que era possível a ocorrência de respiração sincrônica. Este surto foi dividido em início da atividade e fim da atividade, sendo desta forma contabilizada a porcentagem de respiração sincrônica que ocorria no início e no final de cada surto de sincronismo por estado comportamental. Vale ressaltar que esta porcentagem era retirada de uma outra planilha já analisada de respiração sincrônica.
- Freqüência de eventos sincrônicos
Para o cálculo da freqüência de ocorrência de eventos sincrônicos, realizou-se uma contagem dentro de cada estado comportamental (deslocamento, socialização e forrageio), sem discriminar o tipo de evento, apenas o quantificando dentro de uma das três categorias comportamentais. A categoria descanso não foi utilizada nesta análise, pelo fato de não ter sido coletado nenhum evento comportamental que pudesse caracterizar o descanso.
Análise estatística
RESULTADOS E DISCUSSÃO
1. Determinação do padrão comportamental dos botos cinza
1.1. Resultados
- Orçamento de atividades
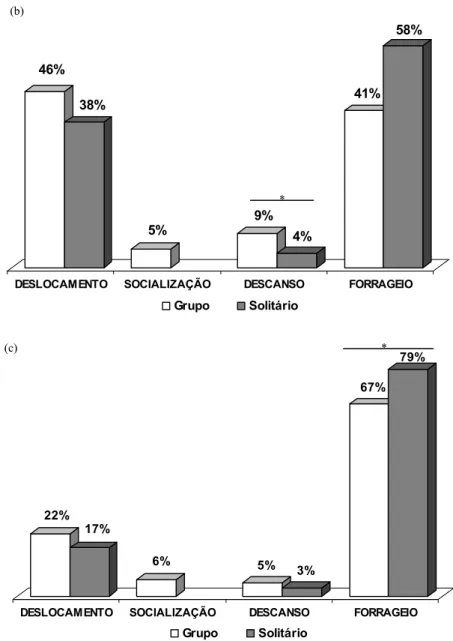
Enseada do Curral - As análises indicam que o orçamento de atividades de indivíduos solitários foi diferente em alguns aspectos do orçamento de atividades dos indivíduos em grupo. Indivíduos solitários empregaram 62% do tempo em forrageio, 37% em deslocamento, e 1% em descanso, não sendo possível haver socialização. Quando em grupo os indivíduos empregaram 41% do tempo em forrageio, 50% em deslocamento, 3% em descanso e 6% em socialização (figura 7a). Estatisticamente a diferença não foi significativa entre o orçamento de atividades de indivíduo solitário e em grupo no deslocamento (t= 2,006; n= 18 e p= 0,061) e no descanso (z= -1,483; n= 18 e p= 0,138), porém a diferença foi significativa para o forrageio (t= -3,176; n= 18 e p< 0,05), ou seja, indivíduos solitários forrageiam mais que indivíduos em grupo.
Enseada do Madeiro 1 – Semelhante à enseada do curral, os orçamentos de atividades dos indivíduos solitários e em grupo também se apresentaram de forma distintas em alguns aspectos. Quando sozinhos na enseada, os animais empregam 58% do tempo em forrageio, 38% em deslocamento e 4% em descanso. Quando em grupo, os animais forrageiam por menos tempo (41%) e deslocam mais (46%), ocorrendo um aumento no comportamento de descanso (9%), e socialização em 5% do tempo. (figura 7b). Entretanto, a única diferença significativa foi no comportamento de descanso (z= -2,490; n= 13 e p< 0,05).
Comparação entre áreas e períodos: Análises indicaram que não existe diferença significativa entre os comportamentos apresentados nas duas enseadas na época antes da chuva. Desta forma, se recalcularmos o orçamento de atividades em grupo unindo os dados das duas áreas temos: deslocamento: 48%; forrageio: 41%; descanso: 4,9%; socialização: 5,3%.
No período após as chuvas há uma mudança no orçamento de atividades: os indivíduos forrageiam significativamente mais tanto solitariamente (madeiro 1 = 58%; madeiro 2= 79%; t= -3,897; n= 13 e p< 0,05) quanto em grupo (madeiro 1 = 40,1%; madeiro 2= 67%; t= -3,865; n= 13 p< 0,05), e se deslocam significativamente menos (madeiro 1 solitário: 38,2%; madeiro 2 solitário: 17,5%; t= 2,999, n= 13; p< 0,05; madeiro 1 grupo: 45,6%; madeiro 2 grupo: 21,8%; t= 3,740; n= 13 e p< 0,05). Verifica-se uma queda no comportamento de descanso, tanto solitário (3,2%) quanto em grupo (4,9%) e um leve aumento na socialização (5,7%). Entretanto, estas diferenças não foram significativas.
Quando separamos o comportamento de forrageio em seus estados componentes de procura e forrageio efetivo (bote e perseguição), verificamos que o forrageio efetivo não apresentou variação de freqüência nos dois períodos, mas a procura foi significativamente maior no período após as chuvas que no período antes das chuvas em ambas as áreas (forrageio madeiro1: 11%; forrageio madeiro 2: 13%; t= -1,669; n= 32 e p= 0,099; procura madeiro1: 30%; procura madeiro2: 54%; t= -5,501; n= 32; p< 0,05).
50%
37%
6%
3% 1%
41% 62%
DESLOCAMENTO SOCIALIZAÇÃO DESCANSO FORRAGEIO Grupo Solitário
Figura 7. Orçamento de atividades em grupo e de indivíduo solitário na enseada do curral (a), na enseada do madeiro 1 (b) e na enseada do madeiro 2 (c). * = diferença significativa.
- Número de indivíduos na enseada
A moda do número de indivíduos presentes em ambas as áreas durante todo o período de estudo foi 2, o número mínimo foi 1 e o número máximo foi 11. Este padrão foi semelhante para cada enseada, e nos dois períodos (1 e 2) (tabela 1). O número modal de indivíduos não se altera quando consideramos cada estado comportamental separadamente.
46%
38%
5% 9% 4%
41% 58%
DESLOCAMENTO SOCIALIZAÇÃO DESCANSO FORRAGEIO Grupo Solitário
(b)
*
22% 17%
6% 5%
3%
67% 79%
DESLOCAMENTO SOCIALIZAÇÃO DESCANSO FORRAGEIO Grupo Solitário
- Faixa etária
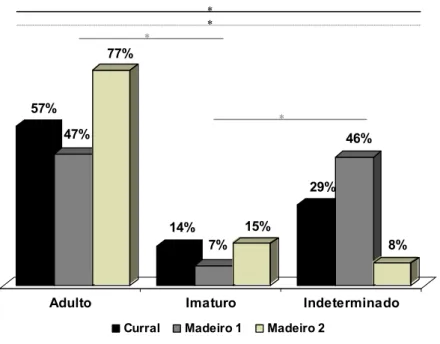
Enseada do Curral – Os resultados indicaram que a maioria dos golfinhos presente na área de estudo são adultos. Estatisticamente a diferença foi significativa entre todas as categorias de faixa etária: adulto e imaturo (t= 6,849; n= 20 e p< 0,05), adulto e indeterminado (t= 3,279; n= 20 e p< 0,05) e imaturo e indeterminado com t= -3,334; n= 20 e p< 0,05 (figura 8).
Enseada do Madeiro 1 – A estatística apontou haver diferença significativa entre as categorias de faixa etária, adulto e imaturo (t= -6,071; n= 13 e p< 0,05) e entre imaturo e indeterminado (t= -8,017; n= 13 e p< 0,05), já entre as categorias imaturo e indeterminado não houve diferença significativa (t= -0,033; n= 13 e p= 0,974) (figura 8).
Enseada do Madeiro 2 – No período após as chuvas verificou-se um aumento no número de adultos e imaturos, e uma diminuição no número de indivíduos com faixa etária indeterminada. Todas as diferenças foram significativas (adulto-imaturo: t= 20,653; n= 37; p< 0,05; adulto-indeterminado: t= 22,863; n= 37; p< 0,05; imaturo-indeterminado: t= 3,111; n= 37 e p< 0,05) (figura 8).
Comparação entre áreas e períodos: Análises indicaram que ocorreram variações significativas no número de indivíduos de cada faixa etária tanto entre as áreas quanto entre os períodos do ano. Houve uma presença maior de indivíduos com faixa etária indeterminada na enseada do madeiro que na enseada do curral (t= -3,075; n= 13 e p< 0,05). No período após as chuvas ocorreram mais adultos e menos indivíduos com faixa etária indeterminada. (t= -3,933; n= 13 e p< 0,05; t= 4,889; n= 13 e p< 0,05, respectivamente).
GERAL DESLOCAMENTO SOCIALIZAÇÃO DESCANSO FORRAGEIO
Moda Máx. Min. Moda Máx. Min. Moda Máx. Min. Moda Máx. Min. Moda Máx. Min.
Curral 2 10 1 2 9 1 3 10 2 2 4 1 2 6 1
Madeiro 1 2 10 1 2 10 1 2 10 2 2 7 1 2 10 1
Madeiro 2 2 11 1 2 9 1 2 11 2 2 6 1 2 7 1
Figura 8. Porcentagem de faixas etárias presentes nas áreas e períodos de estudo. * = diferença significativa.
- Distância entre os indivíduos (Índice de coesão)
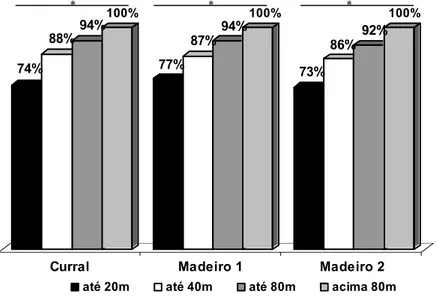
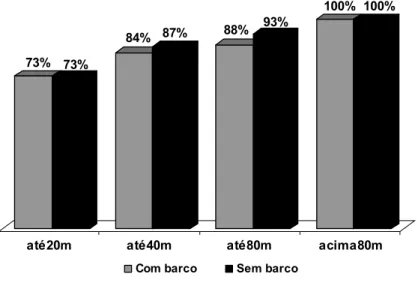
Enseada do Curral – Para o índice de coesão existente entre os golfinhos verificamos que uma média de 74% dos indivíduos presentes na enseada por dia ficam próximos, distando até 20 metros entre si. Este índice sobe para 88% na categoria de distância até 40 metros, e 94% na categoria até 80 metros (figura 9). Estatisticamente a diferença foi significativa entre todas as categorias de distância: até 20 metros e até 40 metros (z= -3,920; n= 20 e p< 0,05), até 20 metros e até 80 metros (z= 3,920; n= 20 e p< 0,05), até 20 metros e acima de 80 metros (z= -3,920; n= 20 e p< 0,05), até 40 metros e até 80 metros (z= -3,823; n= 20 e p< 0,05), até 40 metros e acima de 80 metros (z= -3,823; n= 20 e p< 0,05) e até 80 metros e acima de 80 metros (z= -3,296; n= 20 e p< 0,05 - Vale lembrar que as distancias são cumulativas)
Enseada do Madeiro 1 – O índice de coesão apresentou-se estatisticamente diferente entre todas as categorias de distância: até 20 metros e até 40 metros (t= -10,093; n= 13 e p< 0,05), até 20 metros e até 80 metros (t= -9,367; n= 13 e p< 0,05), até 40 metros e até 80 metros (t= -6,456; n= 13 e p< 0,05) (figura 9).
Enseada do Madeiro 2 – As diferenças foram estatisticamente significativas: até 20 metros e até 40 metros (t= -13,155; n= 37 e p< 0,05), até 20 metros e até 80 metros (t= -14,421 n= 37 p< 0,05), até 40 metros e até 80 metros (t= -9,028; n= 37 e p< 0,05) (figura 9).
57%
47% 77%
14% 7%
15%
29% 46%
8%
Adulto Imaturo Indeterminado
Curral Madeiro 1 Madeiro 2
Comparação entre áreas e períodos: O padrão de espaçamento entre os indivíduos permanece o mesmo, independente da área ou do período (figura 9).
Figura 9. Categorias de distância existente entre os golfinhos nas áreas e períodos de estudo. * = diferença significativa.
1.2. Discussão
Os resultados mostraram uma diferença clara no padrão de atividades dos indivíduos nas situações em que eles estavam solitários ou em grupo. Indivíduos solitários passavam mais tempo forrageando (60 a 80% do tempo) do que indivíduos em grupo (40 a 70% do tempo). Já os indivíduos em grupo realizavam mais deslocamentos e descanso (20 a 45% e 2,5 a 8%, respectivamente) que indivíduos solitários (17 a 38%). Estas diferenças foram significativas em uma ou nas duas áreas, em um ou nos dois períodos estudados.
Esta diferença remete às discussões acerca das vantagens e desvantagens da vida em grupo, e sobre a ocorrência ou não de estratégias de caça coletiva em cetáceos (Neumann & Orams, 2005). A maior proporção de forrageio solitário pode ser um indicativo de evitação de competição por alimento, em que os indivíduos estejam separando o período de suas atividades alimentares de forma a evitar competição com conspecíficos como visto em ungulados (Coté et al., 1997). Por
outro lado, o menor forrageio em grupo pode ser indicativo de maior eficiência no 74%
88%94% 100%
77%
87%94% 100%
73%
86% 92% 100%
Curral Madeiro 1 Madeiro 2
até 20m até 40m até 80m acima 80m