Fitorreguladores e posição de explantes foliares na indução à calogênese em cerejeira-do-mato.
Ciência Rural, v.43, n.10, out, 2013.
1759
Fitorreguladores e posição de explantes foliares na indução à
calogênese em cerejeira-do-mato
Phytoregulators and leaf explants position on callus induction in cerejeira-do-mato
Diego Pascoal GolleI Lia Rejane Silveira ReinigerII ISSN 0103-8478
Ciência Rural, Santa Maria, v.43, n.10, p.1759-1762, out, 2013
Recebido 29.08.12 Aprovado 19.04.13 Devolvido pelo autor 02.08.13 CR-2012-0771.R1
RESUMO
O presente trabalho objetivou avaliar o efeito de diferentes formas de inoculação, no meio nutritivo, de explantes foliares de Eugenia involucrata DC., uma espécie fl orestal nativa com diversas potencialidades econômicas. Foram avaliadas as posições abaxial, adaxial, com e sem cortes no limbo foliar. Foi utilizado o meio de cultura MS acrescido de 10μM de ANA isolado ou das combinações, em μM, de 2,4-D e BAP: 5-5 e 5-10. A posição dos explantes afeta a calogênese e a organogênese em segmentos foliares de E. involucrata, sendo mais adequada a abaxial sem cortes na região do limbo. A associação dos reguladores de crescimento 2,4-D + BAP na concentração de 5-10μM foi mais promissora para a obtenção de calos, especialmente os nodulares, putativos à embriogênese somática.
Palavras-chave: posição de explantes, calos nodulares, organogênese, meios de cultura.
ABSTRACT
The aim of this research was to evaluate the effect of different forms of leaf explants inoculation in nutritive medium of
Eugenia involucrata DC., an native forest species with diverse economic potentials. Inoculations were evaluated at abaxial and adaxial positions, with and without cuts on the leaf surface in the MS nutritive medium supplemented with 10μM NAA or combination, in
μM, of 2.4-D and BAP: 5-5 and 5-10. The explants position inoculation affects the callus induction and organogenesis in E. involucrata leaf segments, being the abaxial the most suitable position, without cuts in the leaf surface. The combination of growth regulators 2.4-D and BAP, at concentrations of 5-10(μM) was most promising of callus obtaining, especially nodular callus, putative at somatic embryogenesis.
Key words: explants position, nodular callus, organogenesis, culture medium.
Eugenia involucrata DC. (Myrtaceae), conhecida popularmente como cerejeira-do-mato, é uma espécie nativa de vários Estados brasileiros. Possui potencial madeireiro, frutícola, paisagístico, ambiental e medicinal (BACKES & IRGANG, 2002; PAROUL, 2007). Dotada de sementes recalcitrantes, sua propagação sexuada é difi cultada, além de só atingir a idade reprodutiva entre os seis e sete anos de idade (CARVALHO, 2008). Dessa forma, justifi ca-se a necessidade de estudos relacionados à cultura de tecidos da espécie.
Calos são grupos de células em divisão, obtidas a partir do estímulo fornecido por substâncias presentes no meio nutritivo e por fatores ambientais. Com incitações específi cas, processos morfogênicos podem conduzir essas células à formação de embriões somáticos, de brotos e raízes, à propagação em grande escala e a estudos de transformação genética, em que é necessário regenerar as plantas a partir de células isoladas (MA et al., 2009; NEUMANN, et al., 2009; VENTURIERI & VENTURIERI, 2004). Esse trabalho objetivou avaliar a infl uência de diferentes formas de inoculação de explantes foliares no meio de cultura e de combinações de fi torreguladores na formação de diferentes tipos de calos em E. involucrata.
Os explantes foram isolados a partir de folhas jovens de plantas de quatro anos de idade mantidas em casa de vegetação. Após a
NOTA
-ILaboratório de Cultura de Tecidos Vegetais in vitro, Polo de Inovação Tecnológica do Alto Jacuí, Universidade de Cruz Alta
(UNICRUZ), 98020-290, Cruz Alta, RS, Brasil. E-mail: dgolle@unicruz.edu.br. Autor para correspondência.
IINúcleo de Biotecnologia e Melhoramento, Departamento de Fitotecnia, Universidade Federal de Santa Maria (UFSM), Santa Maria, RS,
1760 Golle & Reiniger
Ciência Rural, v.43, n.10, out, 2013. coleta, as folhas receberam um pré-tratamento de
desinfestação, sendo mantidas em solução de Benlate 500® (benomyl) a 1g L-1 + sulfato de estreptomicina
a 0,1g L-1 por 30min. Em seguida, foram lavadas
com detergente comercial, água corrente (5min) e submetidas à sequência: imersão em etanol a 70% (v/v) por 30s; enxágue em água destilada; imersão em solução de hipoclorito de sódio (NaOCl) a 1,5% (v/v) com três gotas de detergente comercial por 15min; e, por fi m, triplo enxágue em água destilada estéril.
O experimento foi conduzido em delineamento inteiramente casualizado, utilizando-se esquema bifatorial 4x3, em que os níveis do fator “A” corresponderam às posições de inoculação (abaxial e adaxial, com ou sem corte no limbo foliar – seis cortes) dos explantes foliares no meio de cultura, e os níveis do fator “B” às diferentes inserções de reguladores de crescimento no meio, a saber: 10μM de ácido α-naftalenoacético (ANA), isoladamente, ou combinações entre ácido 2,4-diclorofenóxiacético (2,4-D) e 6-benzilaminopurina (BAP) nas concentrações 5-5 e 5-10 (em μM). Os explantes consistiram de fragmentos foliares de 1x1,5cm, contendo corte em toda a região de bordadura. Os níveis do fator “B” foram escolhidos de acordo com resultados observados em experimentos anteriores de indução à calogênese. Utilizaram-se 10 repetições, cada uma composta por um frasco com capacidade para 150mL, contendo 30mL de meio de cultura e dois explantes, totalizando 120 unidades experimentais e 240 explantes. O meio de cultura utilizado foi o MS (MURASHIGE e SKOOG, 1962), contendo 30g L-1
de sacarose, 100mg L-1 de mio-inositol e 7g L-1 de
ágar (pH 5,8). As parcelas permaneceram em sala de cultivo com temperatura de 25±3°C e fotoperíodo de 16h (com intensidade luminosa de 20μmol m-2 s-1)
obtido por meio de lâmpadas fl uorescentes “luz do dia”. Os frascos permaneceram 45 dias no escuro (em caixas de papel). Após, foram transferidos para a luz por 25 dias adicionais.
Ao fi nal desses 70 dias, foram avaliadas as variáveis: formação de calos, formação de calos rizogênicos, calos friáveis, calos fi rmes, calos nodulares e oxidação fenólica, todas expressas em porcentagem. Também foram atribuídas notas aos calos, a saber: 0) ausência de formação calogênica; 1) formação de calo cicatricial ou início da formação calogênica; 2) formação calogênica em toda a bordadura (ou nas regiões de cortes); 3) formação calogênica com crescimento secundário; e 4) formação calogênica com crescimento abundante. A normalidade dos erros foi avaliada pelo teste de Kolmogorov-Smirnov e a homocedasticidade pelo
teste de Bartlett. Os dados foram submetidos à análise de variância e as médias foram comparadas pelo teste de Tukey a 5% com auxílio do programa SISVAR versão 4.0 (FERREIRA, 2000).
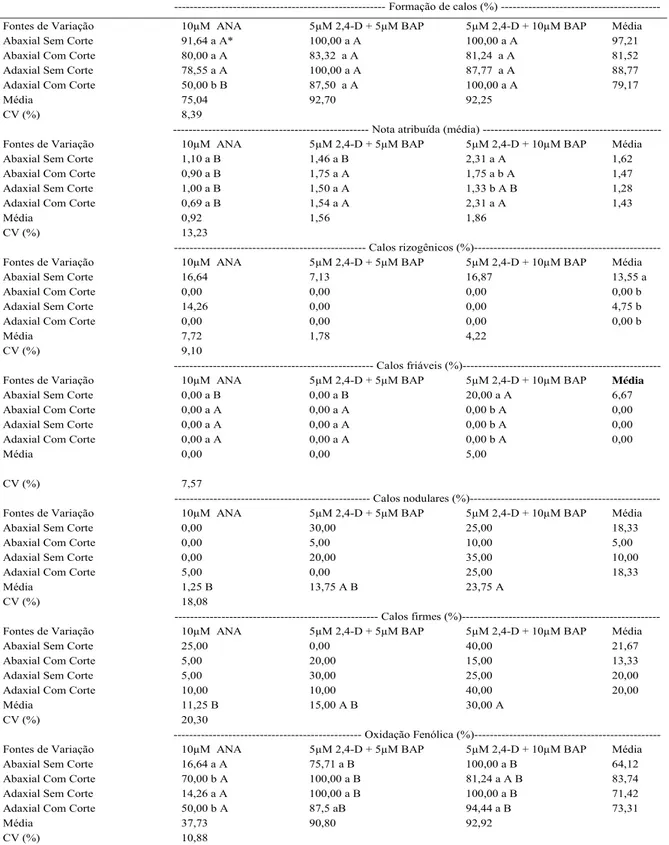
Para a variável formação de calos, observou-se interação (P=0,0030) entre os níveis dos fatores testados (Tabela 1). Com a adição de 10μM de ANA, a inoculação dos explantes na posição adaxial com cortes foi signifi cativamente inferior a todas as demais, que não diferiram entre si. Só houve distinções entre as formas de uso dos reguladores de crescimento quando os explantes foram inoculados na posição adaxial com corte. Esse resultado é contrário àquele observado em Platanus occidentalis L. (Platanaceae), em que a inoculação dos explantes na posição adaxial contendo cortes no limbo foliar foi necessária para obtenção de calos e brotações (SUN et al., 2009). Para as notas atribuídas aos calos (Tabela 1), também houve interação (P=0,0042), em que, com o uso da associação de 5μM de 2,4-D e 10μM de BAP, a inoculação, na posição abaxial sem cortes e adaxial com cortes, favoreceu a formação de calos. Poucos trabalhos com espécies lenhosas relataram a infl uência da forma de inoculação de explantes no meio nutritivo e a infl uência desse fator na calogênese. Em Juniperus oxycedrus L. (Cupressaceae), a inoculação, na posição adaxial, dos explantes no meio nutritivo, diferiu dos resultados observados para E. involucrata (GOMEZ & SEGURA, 1996); já para Litchi chinensis Sonn. (Sapindaceae), houve maior formação de calos quando a face abaxial fi cou em contato com o meio nutritivo, semelhante ao observado no presente estudo para E. involucrata (RAHARJO & LITZ, 2007).
Observaram-se diferenças de acordo com a forma de inoculação dos explantes (P=0,0001) na formação de calos rizogênicos (Tabela 1), sendo a posição abaxial sem cortes superior às demais. Possivelmente, a forma de contato das células dos tecidos com o meio nutritivo favoreceu essa resposta organogênica para a formação de raízes. Esses dados são díspares em relação aos de PIERIK (1990), que observou aspectos benéfi cos no aumento da superfície de contato dos explantes com o meio nutritivo por meio de cortes no limbo.
Fitorreguladores e posição de explantes foliares na indução à calogênese em cerejeira-do-mato.
Ciência Rural, v.43, n.10, out, 2013.
1761
Tabela 1 - Efeito da forma de inoculação de explantes foliares de Eugenia involucrata DC. e de diferentes reguladores de crescimento na formação de calos (%), média de notas atribuídas aos calos; formação de calos rizogênicos (%); de calos friáveis (%); de calos nodulares (%); de calos firmes (%); e oxidação fenólica (%) aos 70 dias de cultivo in vitro.
--- Formação de calos (%)
---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 91,64 a A* 100,00 a A 100,00 a A 97,21
Abaxial Com Corte 80,00 a A 83,32 a A 81,24 a A 81,52
Adaxial Sem Corte 78,55 a A 100,00 a A 87,77 a A 88,77
Adaxial Com Corte 50,00 b B 87,50 a A 100,00 a A 79,17
Média 75,04 92,70 92,25
CV (%) 8,39
---- Nota atribuída (média) ---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 1,10 a B 1,46 a B 2,31 a A 1,62
Abaxial Com Corte 0,90 a B 1,75 a A 1,75 a b A 1,47
Adaxial Sem Corte 1,00 a B 1,50 a A 1,33 b A B 1,28
Adaxial Com Corte 0,69 a B 1,54 a A 2,31 a A 1,43
Média 0,92 1,56 1,86
CV (%) 13,23
--- Calos rizogênicos (%)---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 16,64 7,13 16,87 13,55 a
Abaxial Com Corte 0,00 0,00 0,00 0,00 b
Adaxial Sem Corte 14,26 0,00 0,00 4,75 b
Adaxial Com Corte 0,00 0,00 0,00 0,00 b
Média 7,72 1,78 4,22
CV (%) 9,10
--- Calos friáveis (%)---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 0,00 a B 0,00 a B 20,00 a A 6,67
Abaxial Com Corte 0,00 a A 0,00 a A 0,00 b A 0,00
Adaxial Sem Corte 0,00 a A 0,00 a A 0,00 b A 0,00
Adaxial Com Corte 0,00 a A 0,00 a A 0,00 b A 0,00
Média 0,00 0,00 5,00
CV (%) 7,57
--- Calos nodulares (%)---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 0,00 30,00 25,00 18,33
Abaxial Com Corte 0,00 5,00 10,00 5,00
Adaxial Sem Corte 0,00 20,00 35,00 10,00
Adaxial Com Corte 5,00 0,00 25,00 18,33
Média 1,25 B 13,75 A B 23,75 A
CV (%) 18,08
--- Calos firmes (%)---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 25,00 0,00 40,00 21,67
Abaxial Com Corte 5,00 20,00 15,00 13,33
Adaxial Sem Corte 5,00 30,00 25,00 20,00
Adaxial Com Corte 10,00 10,00 40,00 20,00
Média 11,25 B 15,00 A B 30,00 A
CV (%) 20,30
--- Oxidação Fenólica (%)---Fontes de Variação 10μM ANA 5μM 2,4-D + 5μM BAP 5μM 2,4-D + 10μM BAP Média
Abaxial Sem Corte 16,64 a A 75,71 a B 100,00 a B 64,12
Abaxial Com Corte 70,00 b A 100,00 a B 81,24 a A B 83,74
Adaxial Sem Corte 14,26 a A 100,00 a B 100,00 a B 71,42
Adaxial Com Corte 50,00 b A 87,5 aB 94,44 a B 73,31
Média 37,73 90,80 92,92
CV (%) 10,88
1762 Golle & Reiniger
Ciência Rural, v.43, n.10, out, 2013. (P=0,0168), em que a média da combinação 5μM
de 2,4-D + 10μM de BAP foi superior a 10μM de ANA, mas que não diferiu da combinação 5μM de 2,4-D + 5μM de BAP. A formação de calos nodulares, por sua vez, foi afetada apenas pelos diferentes usos de fi torreguladores (P=0,0006). Esses calos, putativos à embriogênese, tiveram sua maior ocorrência com o uso das combinações de 2,4-D e BAP na concentração, em μM, de 5-10. A oxidação fenólica apresentou interação signifi cativa (P=0,0001) entre os níveis dos fatores (Tabela 1). Houve redução nas taxas oxidativas dos explantes sem cortes quando se utilizou o fi torregulador ANA, corroborando os registros de ANDRADE et al. (2000) e LEDO et al. (2002), que relatam o aumento no índice de fenóis devido às injúrias. Além disso, sabe-se que a atividade da fenilalanina amoniliase (PAL), enzima que atua no metabolismo de compostos fenólicos, é aumentada pela agressão aos tecidos (TAIZ & ZEIGER, 2009).
A melhor forma de inoculação de explantes foliares no meio nutritivo é na posição abaxial em contato com o meio, sem cortes no limbo foliar, o que favorece a obtenção de diversos tipos de calos em E. involucrata, e o uso da combinação de 2,4-D e BAP 5-10 favorece o surgimento de calos putativos à embriogênese somática.
REFERÊNCIAS
ANDRADE, M.W. et al. Micropropagação de aroeira (Myracrodruon urundeuva Fr. Allemao). Ciência e Agrotecnologia, Lavras, v.24, n.1, p.174 - 180, 2000.
BACKES, A.; IRGANG, B. Árvores do Sul: guia de identifi cação e interesse ecológico. Porto Alegre: Pallotti, 2002. 275p.
CARVALHO, P.E. Espécies arbóreas brasileiras. Brasília: Embrapa Informação Tecnológicas; Colombo: Embrapa Florestas, 2008. v.3, 593p.
FERREIRA, D.F. Análises estatísticas por meio do Sisvar para Windows versão 4.0. In: REUNIÃO ANUAL DA REGIÃO BRASILEIRA DA SOCIEDADE INTERNACIONAL DE BIOMETRIA, 45., 2000, São Carlos, SP. Anais... São Carlos: Universidade Federal de São Carlos, 2000. p.255-258.
GOMEZ, M.P.; SEGURA, J. Morphogenesis in leaf and single-cell cultures of mature Juniperus oxycedrus. Tree Physiology, Victoria, v.16, n.8, p.681-686, 1996. Disponível em: <http://
treephys.oxfordjournals.org/content/16/8/681.full.pdf>. Acesso em: 23 jul. 2012. doi:10.1093/treephys/15.9.625.
LEDO, A.S. et al. Explantes de cupuaçuzeiro submetidos a diferentes condições de cultura in vitro. Revista Brasileira de Fruticultura, Jaboticabal, v.24, n.3, p.604-607, 2002. Disponível em: <http:// dx.doi.org/10.1590/S0100-29452002000300005>. Acesso em: 16 abr. 2012. doi:10.1590/S0100-29452002000300005.
MA, X.Y. et al. Leaf callus induction and suspension culture establishment in lychee (Litchi chinensis Sonn.) cv. ‘Huaizhi’.
Acta Physiologiae Plantarum, Kraków, v.31, n.2, p.401-405, 2009. Disponível em: <http://dx.doi.org/ 10.1007/s11738-008-0223-x.> Acesso em: 22 ago. 2012. doi: 10.1007/s11738-008-0223-x.
MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum, Copenhagen, v.15, p.473-49, 1962.
NEUMANN, K.H. et al. Plant cell and tissue culture– a toll in biotechnology: basics and application. Berlin: Springer Verlang, 2009. 333p.
PAROUL, N. et al. Avaliação da composição química do óleo volátil de cerejeira (Eugenia involucrataDC). In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE QUÍMICA, 30., 2007, Águas de Lindóia. Anais... Águas de Lindóia: Sociedade Brasileira de Química, 2007. p.1-1. Disponível em: <http://sec.sbq.org.br/ cdrom/30ra/resumos/T1853-1.pdf>. Acesso em: 23 mar. 2011.
PIERIK, R.L.M. Cultivo in vitro de las plantas superiores. Madrid: Mundi-Prensa, 1990. 326p.
RAHARJO, S.H.T.; LITZ, R.E. Somatic embryogenesis and plant regeneration of litchi (Litchi chinensis Sonn.) from leaves of mature phase trees. Plant Cell, Tissue and Organ Culture, Dordrecht, v.89, n.2-3, p.113-119, 2007. Disponível em: <http:// www.springerlink.com/content/64185kp7047052mp/>. Acesso em: 28 ago. 2012. doi:10.1007/s11240-007-9219-2.
SUN, Y. et al. Adventitious bud regeneration from leaf explants of Platanus occidetalis L. ande genetic stability assessment.
Acta Physiologiae Plantarum, Kraków, v.31, n.1, p.33-41, 2009. Disponível em: <http://dx.doi.org/ DOI: 008-0196-9>. Acesso em: 13 jul. 2012. doi: DOI: 10.1007/s11738-008-0196-9.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 4ed. Porto Alegre: Artmed, 2009. 819p.
VENTURIERI, G.A.; VENTURIERI, G.C. Calogênese do híbrido