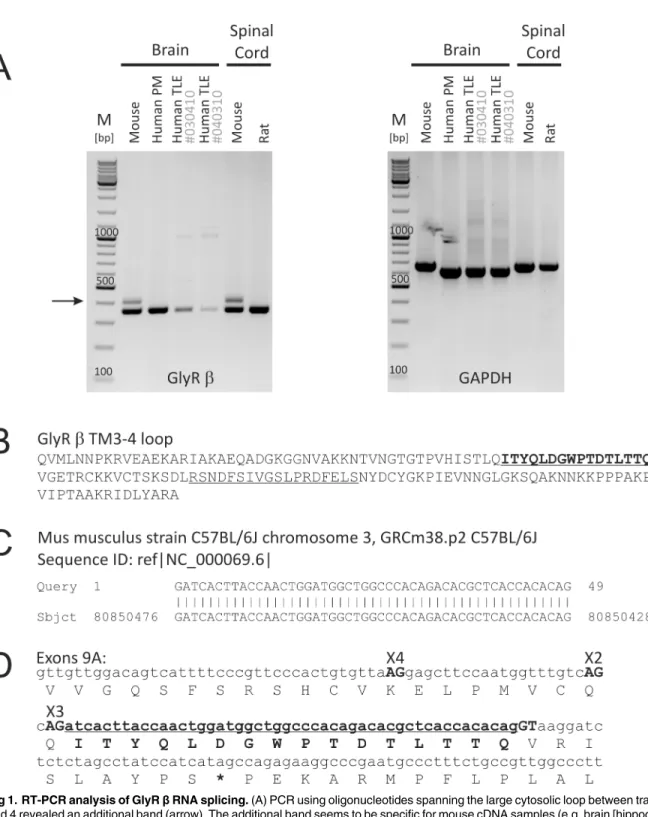
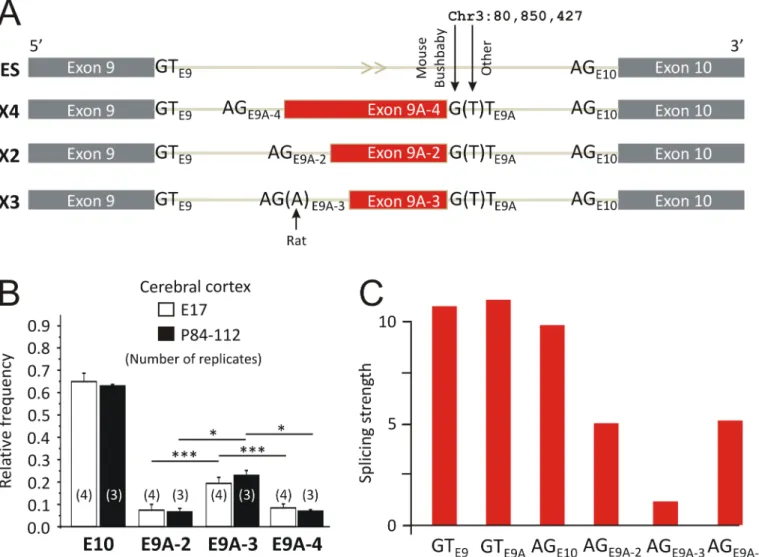
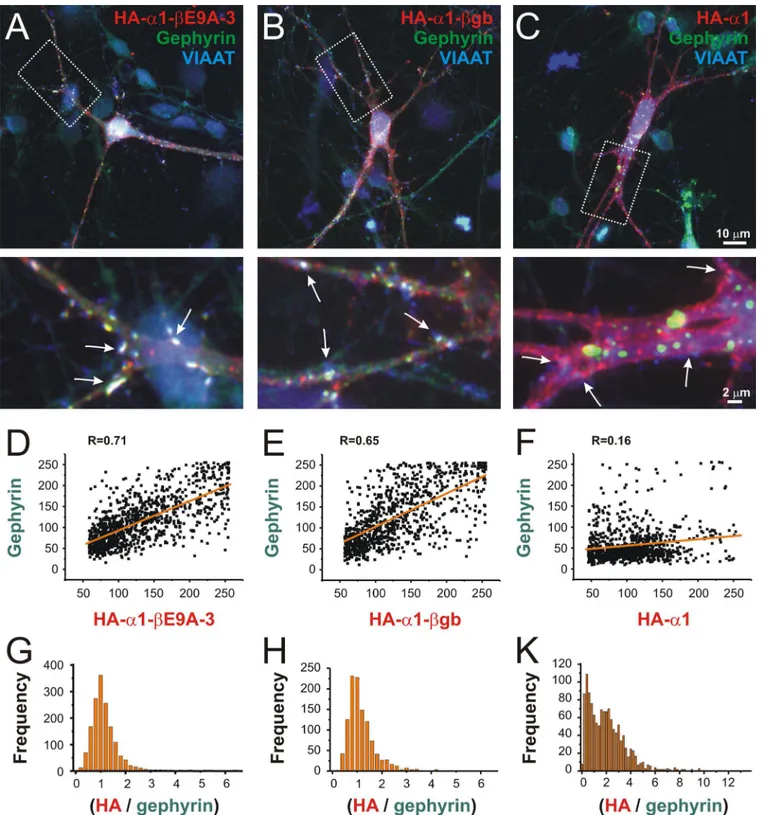
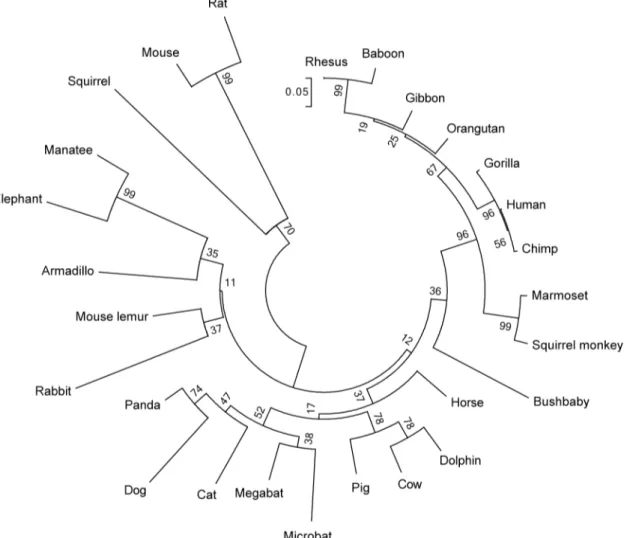
Identification of a new genomic hot spot of evolutionary diversification of protein function.
Texto
Imagem
Documentos relacionados
We also determined the critical strain rate (CSR), understood as the tangent of the inclination angle between the tangent to the crack development curve and the crack development
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Christian: - Então... Vai me contar o que aconteceu? Jack: - Dois caras agrediram Marc Silverman. C: - Agrediram Marc Silverman? Mas não agrediram você. C: - Fiz cirurgia em um
Here we review the origin of domesticated Oryza sativa from wild progenitors, the ecological and genomic diversity of the Oryza genus, and the stress tolerance variation observed
With the aim of evaluating the SRAP (sequence-related amplified polymorphism) technique for linkage mapping in Pisum sativum L., a F 2 mapping population.. derived from an
São diversos os fundamentos desde o âmbito filosófico até o âmbito puramente social que permitem a classificação da liberdade de expressão como um direito fundamental
O exame Holter da Piguita teve como ritmo de base um ritmo sinusal, mas eram também frequentes os episódios de arritmia sinusal respiratória bem como os de
de sólidos solúveis totais, índice de refração, pH e atividade de água, durante a etapa de infusão do álcool de cereais com a banana, determinando o melhor parâmetro e tempo
