ELIAS SAN VITO
GLICERINA BRUTA NA ALIMENTAÇÃO DE VACAS LEITEIRAS
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Zootecnia, para obtenção do título de
VIÇOSA
ELIAS SAN VITO
GLICERINA BRUTA NA ALIMENTAÇÃO DE VACAS LEITEIRAS
Dissertação apresentada á Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Zootecnia, para obtenção do título de
Aprovada: 08 de Julho de 2010.
___________________________________ ___________________________________
Prof. Sebastião de C. Valadares Filho Prof. Marcelo de Andrade Ferreira (Co-Orientador)
___________________________________ ___________________________________ Profa. Rilene Ferreira Diniz Valadares Prof. Rogério de Paula Lana
______________________________________ Prof. José Maurício de Souza Campos
ii Aos meus pais, Ledo e Idelve e meus irmãos Ivete, Ivanor e Diego, pelo amor e carinho e principalmente por acreditar que isso seria possível.
Ao Departamento de Zootecnia da Universidade Federal de Viçosa, pela oportunidade e pelo apoio na realização do curso.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e ao Instituto Nacional de Ciência e Tecnologia (INCT) de Ciência Animal pelo financiamento do projeto.
Ao professor José Maurício de Souza Campos, pela confiança e orientação.
Aos professores Sebastião de Campos Valadares Filho e Edênio Detmann pelos importantes ensinamentos.
Aos professores Rogério de Paula Lana, Rilene Ferreira Diniz Valadares Filho e Marcelo de Andrade Ferreira pela participação na banca avaliadora.
Ao professor André Soares de Oliveira por toda a ajuda e conselhos.
Aos meus amigos Mateus Pies Gionbelli e Rafael Mezzomo, pelo companheirismo, incondicional apoio e valiosa amizade.
Ao meu amigo Edfran Stocco Granado, pela dedicação na realização do experimento.
A Kássia Armondes, pela dedicação e apoio.
A republica “Os Cavalo Deitado”, nas pessoas de Goiano, Edfran, Daniel, Hamilton, Luizão, Leo, Guella, Liminha, Rafael, por ser minha casa em Viçosa, pela amizade e companheirismo em todas as horas.
Aos amigos e colegas: João Paulo, Simone, Amanda, Ivanna, Goiano, Denise, Márcia e demais, pelo intercâmbio de conhecimento e idéias, pelo convívio, apoio e valiosa amizade.
Aos funcionários da Unidade de Ensino, Pesquisa e Extensão em Gado de Leite, do Laboratório de Nutrição Animal e da Zootecnia, pelas contribuições na realização deste trabalho.
iii ELIAS SAN VITO, filho de Ledo San Vito e Idelve Terezinha San Vito, nasceu em Maravilha, Estado de Santa Catarina, em 16 de novembro de 1985.
Em 2004, ingressou na Universidade do Estado de Santa Catarina – UDESC, onde obteve o título de Zootecnista, colando grau em 19 de julho de 2008.
iv
RESUMO...v
ABSTRACT...vii
INTRODUÇÃO...1
Material e Métodos...5
Resultados e Discussão...14
Conclusões...26
v SAN VITO, Elias, M.Sc., Universidade Federal de Viçosa, Julho de 2010.
! " "# Orientador: José Maurício de Souza Campos. Co-orientadores: Sebastião de Campos Valadares Filho e Edenio Detmann.
vii SAN VITO, Elias, M.Sc., Universidade Federal de Viçosa, July 2010. $ % "
& & % '"# Adviser: José Maurício de Souza Campos. Co-advisers: Sebastião de Campos Valadares Filho and Edenio Detmann.
1
()
As mudanças na cadeia produtiva do leite que ocorreram nas últimas décadas, aliadas ao grande potencial de crescimento desse setor, devido à disponibilidade de recursos naturais, como terra, água e infra-estrutura, elevaram a produção de leite a um lugar de destaque no agronegócio brasileiro.
O Brasil apresenta um dos menores custos de produção de leite do mundo (Carvalho, 2007), destacando-se ainda por possuir uma grande disponibilidade de produtos para utilização como ingredientes em dieta para animais. Atualmente, o crescimento da participação de biodiesel na matriz energética mundial estimulou o interesse no cultivo e processamento de oleaginosas no Brasil, criando oportunidades para a produção de ruminantes, pela possibilidade de utilização de co-produtos resultantes do processo de extração do óleo das sementes e transformação deste em biodiesel.
2 A glicerina é um triol viscoso, incolor, inodoro e higroscópio resultante, entre outros, do processo de transformação de um triglicerídeo em biodiesel, a partir de uma reação de transesterificação, na presença de um catalisador e de um álcool de cadeia curta (metanol ou etanol). Nesse processo, a glicerina, denominada glicerina bruta, contém de 75 a 85% de glicerol, sendo o restante composto por água, óleo (7 a 13%), minerais oriundo dos catalisadores (2 a 3%) e álcool (<0,5%) (Donkin & Doane, 2007). Para cada 90 m³de biodiesel produzidos são gerados 10 m³de glicerina. Com a obrigatoriedade da adição de biodiesel ao diesel de petróleo, espera-se um excedente anual de 150 mil toneladas para o ano de 2013 (B5), o que possivelmente levará a uma redução nos preços. Esse cenário indica a necessidade de viabilização comercial deste volume extra de glicerina, buscando outras aplicações. Atualmente, a glicerina purificada (com 99,5% de glicerol) é aplicada na indústria de cosméticos, saboaria e fármacos, setores incapazes de sozinhos, absorverem o volume de glicerina gerado com a produção de biodiesel (Gonçalves et al., 2006).
3 em vacas de leite de alta produção, utilizando-se baixos níveis na dieta (< 5%, base da MS), sendo amplamente documentado (Johnson et al., 1954; Fisher et al., 1971; 1973; DeFrain et al., 2004). No entanto, existem poucas informações sobre o uso como macro-ingrediente em dietas de ruminantes. Em estudo recente, pesquisadores americanos avaliaram o uso da glicerina de óleo vegetais bi-destilada (99,5% de glicerol) em dietas de vacas de leite de alta produção (37 kg/dia) e concluíram que o glicerol pode ser usado como macro-ingrediente em até 15%, base da MS da dieta, em substituição ao milho grão, sem afetar o consumo de matéria seca, a produção e a composição do leite (Donkin & Doane, 2007). Todavia, como a glicerina obtida do processo de transesterificação do óleo apresenta-se na forma bruta, com impurezas, os impactos no consumo, na digestibilidade dos nutrientes e no desempenho animal podem ser diferentes aos obtidos com a glicerina purificada, de custo mais elevado.
4 Diante do exposto, é fato de que a glicerina purificada (99,5% de glicerol) pode ser usada na alimentação de vacas leiteiras, até o nível de 15% em base da matéria seca, sem que haja prejuízo no consumo de matéria seca e na produção e composição do leite. Assim, estudos sobre a viabilidade econômica e biológica do uso da glicerina bruta (84% glicerol) na alimentação de vacas leiteiras são inexistentes, o que limita o uso deste material na alimentação animal. Além disso, as informações existentes não possibilitam gerar recomendações nutricionais precisas do seu uso na alimentação de bovinos de maior potencial de produção.
5
* "
O experimento foi conduzido na Unidade de Ensino, Pesquisa e Extensão em Gado de Leite, Departamento de Zootecnia da Universidade Federal de Viçosa, no período de setembro a novembro de 2009. Foram utilizadas 12 vacas da raça Holandesa, distribuídas em três quadrados latinos 4 x 4, de acordo com o período de lactação. Os animais iniciaram o experimento com média de 70 dias de lactação e no final do experimento apresentaram menos de 150 dias de gestação. O experimento foi constituído por quatro períodos, com duração de 21 dias cada, sendo os 14 primeiros dias de adaptação às dietas e os demais para coleta de dados. Os animais foram alimentados com quatro dietas experimentais referentes a quatro proporções de substituição do milho grão (MG) pela glicerina bruta (GB): 0%; 33,3; 66,6% e 100%, na base da matéria seca (MS), que corresponderam a 0%, 7%, 14% e 21% de inclusão de GB na dieta (MS).
As dietas foram formuladas para serem isonitrogenadas, contendo 16% de proteína bruta (PB), de forma a atender as exigências nutricionais de uma vaca com 600 kg de peso corporal, produzindo 30 kg/dia de leite com 3,5% de gordura (NRC, 2001). O glúten de milho (63 %PB) e a mistura contendo nove partes de uréia e uma parte de sulfato de amônio foi utilizada para ajustar o teor de PB da dieta. Os animais receberam ração na proporção de 50:50 em volumoso e concentrado, com base da MS. Silagem de milho foi utilizada como fonte exclusiva de volumoso. A mistura de microminerais foi balanceada para atender 100% das exigências nutricionais, segundo o NRC (2001) (Tabela 1).
6 A silagem, a ração concentrada e a GB foram pesados separadamente e misturados dentro do cocho dos animais da hora do fornecimento da alimentação.
Tabela 1 – Proporção dos ingredientes nas dietas experimentais, em porcentagem na material seca
Ingredientes Níveis de substituição do milho grão pela GB
0 33,3 66,6 100
Silagem de milho 49,92 49,98 50,05 50,12
Milho grão moído 21,00 14,02 7,02 0,00
Glicerina bruta1 0,00 6,88 13,78 20,70
Farelo de soja 18,54 18,57 18,59 18,62
Farelo de trigo 9,06 8,34 7,60 6,88
Glúten de milho 0,00 0,60 1,21 1,81
Uréia/sulfato de amônio2 0,00 0,13 0,26 0,39
Mistura mineral3 1,48 1,48 1,49 1,49
1
Glicerina bruta com 84% de glierol. 2 relação 9:1; 344,60% de Calcário (Ca) ; 9% de fosfato bicálcico
(P); 1,5% de Sulfato de Mg (Mg); 1,23% de flor de enxofre (S); 20% de bicarbonato de sódio; 22% de sal (Na); 2300 ppm de Cu; 8000 ppm de Zn; 1800 ppm de Mn; 160 ppm de I; 185 ppm de Co; 40 ppm de Se.
No período de coleta, foram feitas pesagens e amostragens diárias de silagem de milho e rações concentradas fornecidas e das sobras de cada tratamento. As amostras foram armazenadas a -15ºC para posteriores análises químicas.
As fezes foram coletadas diretamente na ampola retal, uma vez ao dia, às 8:00, 10:00, 12:00, 14:00 e 16:00 horas, do 16o ao 20o dias de cada período experimental, respectivamente. As amostras diárias de fezes de cada animal, em cada período, foram armazenadas a -15oC para posterior secagem e análises químicas.
7 diárias de fezes secas ao ar de cada animal, em cada período, foram feitas amostras compostas. A composição química dos alimentos e das dietas são apresentados nas tabelas 2 e 3, respectivamente.
Tabela 2 – Composição química dos ingredientes utilizados nas dietas experimentais
Itens1 Alimentos Silagem de milho Milho grão Farelo de soja Farelo de trigo Glúten de milho Glicerina bruta
MS (%) 27,06 87,78 88,48 88,05 90,58 89,66
MO2 95,69 98,99 93,93 95,30 98,33 92,52
PB2 6,27 8,80 50,70 18,18 68,85 0,00
NNP3 43,29 10,02 10,04 18,00 6,74 0,00
NIDN3 15,45 21,27 16,43 21,82 11,08 0,00
EE2 3,05 4,12 1,58 3,31 3,33 6,94
FDNcp2 52,16 16,88 16,54 39,86 4,41 0,00
CNF2 32,40 69,16 25,11 33,95 21,74 85,58
FDAi2, 4 14,71 0,24 0,54 7,85 0,00 0,00
1
MS = Matéria seca; MO = Matéria orgânica; PB = Proteína bruta; NNP = Nitrogênio não protéico; NIDN = nitrogênio insolúvel em detergente neutro; EE = Extrato etéreo; FDNcp = Fibra em detergente
neutro corrigida para cinzas e proteína; CNF = Carboidratos não-fibrosos; FDAi = Fibra em detergente
ácido indigestível. 2 % da MS; 3 % do N total; 4 Obtido após incubação ruminal por 264 horas.
A quantidade total de MS fecal excretada foi estimada pela concentração de fibra em detergente ácido indigestível (FDAi), obtidas após incubação ruminal dos alimentos, sobras e fezes em sacos F57 (Ankon®) por 264 horas, segundo Casali et al. (2008).
8 foram realizadas em analisador de fibra (Ankon 220), utilizando sacos de TNT (tecido-não-tecido), com dimensões de 5 x 5 cm, mantendo-se relação média de 20 mg de MS/cm2 de tecido e 100 mL de detergente/g de amostra seca ao ar. A quantificação de nitrogênio não protéico (NNP) dos alimentos foi realizada segundo Licitra et al. (1996).
Tabela 3 – Composição química média das dietas experimentais ofertadas Itens1 Nível de substituição do milho grão pela GB (% da MS)
4
0 33,3 66,7 100
MS, % 57,79 57,91 58,02 58,14
MO2 94,60 94,17 93,75 93,32
PB2 16,03 16,07 16,14 16,18
NNP3 27,20 26,50 25,80 25,10
NIDN3 17,20 15,64 14,07 12,50
EE2 2,98 3,17 3,36 3,55
FDNcp2 36,26 34,86 33,45 32,04
CNF2 39,34 40,31 41,28 42,26
FDAi2 8,20 8,14 8,08 8,01
1
MS = Matéria seca; MO = Matéria orgânica; PB = Proteína bruta; NNP = Nitrogênio não protéico;NIDN
= nitrogênio insolúvel em detergente neutro; EE = Extrato etéreo; FDNcp = Fibra em detergente neutro
corrigida para cinzas e proteína; CNF = Carboidratos não-fibrosos; FDAi = Fibra em detergente ácido
indigestível. 2 % da MS. 3 % do N total. 4 GB = Glicerina bruta com 84% de glierol.
9 O teor de glicerol (84,41%), ácidos graxos totais (6,95%) e metanol (8,64%) da glicerina bruta foram obtidos por cromatografia gasosa em cromatógrafo a gás modelo CG 17ª marca SHIMADZU com detector FID. A amostra foi colocada em um funil de separação de 250 mL e extraída com volume de 25 mL de água. A mistura foi agitada várias vezes, sendo em seguida deixada em repouso por 1 hora. Foram formadas duas fases, em que a fase inferior foi recolhida e posteriormente injetada no cromatógrafo a gás. Foi utilizada coluna capilar carbowax (30 m x 0,25 mm). Para a separação cromatográfica, 1 µL de amostra foi injetado com auxílio de seringa de 10 µL (Hamilton®) sistema Split = 10. O gás nitrogênio foi utilizado como carreador com velocidade linear programada para 26,4 cm/s e os gases hidrogênio e ar sintético foram a chama do detector. As temperaturas do injetor e do detector foram de 280ºC e 300ºC, respectivamente. A temperatura inicial da coluna foi de 40ºC (mantida por 7 minutos), aumentando em 2ºC por minuto até atingir 80ºC e 6ºC por minuto até atingir 240ºC (mantida por 40 minutos), totalizando 93 minutos de análise. O fluxo do gás de arraste na coluna foi de 1,09 mL/minuto. A identificação dos compostos foi realizada através do tempo de retenção do padrão correspondente.
10 proteína, extrato seco total e extrato seco desengordurado, segundo métodos descritos pelo International Dairy Federation (1996); a segunda alíquota foi desproteinizada com ácido tricloroacético (10 mL de leite misturados com 5 mL de ácido tricloroacético a 25%), filtrada em papel-filtro e armazenada a –15oC para posterior análise de alantoína. A produção de leite (PL) corrigida para 3,5% de gordura (PLC) foi calculada segundo Sklan et al. (1992), pela seguinte fórmula: PLC= (0,432 + 0,1625 x % gordura do leite) x produção de leite em kg/dia.
No sétimo dia de adaptação e no final de cada período experimental foram feitas pesagens individuais dos animais para avaliar a variação de peso. O peso dos animais correspondeu à média de duas pesagens, feitas antes do fornecimento da alimentação e após a ordenha.
Amostras de sangue foram coletadas no 18o dia por punção da veia coccígea, utilizando tubos de ensaio com anticoagulante (EDTA). Imediatamente, foram centrifugadas a 2.700 x g por 20 minutos sendo então retiradas amostras de plasma que foram acondicionadas em tubos ependorf e congeladas a –15oC para posteriores análises de concentração de nitrogênio uréico. Uma segunda amostra de sangue foi coletada utilizando-se tubos contendo fluoreto de sódio, que após a coleta foram homogeneizados lentamente, armazenados em caixa de isopor com gelo moído e imediatamente enviados ao laboratório, para realização de análise de glicose.
11 alantoína (AL), ácido úrico (AU) e creatinina. Imediatamente antes das análises, as amostras de urina foram descongeladas e centrifugadas a 2.000 x por 15 minutos.
As análises de AL na urina e no leite foram feitas pelo método colorimétrico, conforme descrito por Chen & Gomes (1992). As análises de uréia foram realizadas por meio de sistema enzimático-colorimétrico pelo método urease, utilizando-se comerciais (Labtest Diagnóstica S.A.). As análises de ácido úrico na urina foram realizadas por meio do método enzimático-Trinder, utilizando-se comerciais (Labtest Diagnóstica S.A.) As análises de creatinina na urina foram realizadas por meio do método de ponto final com picrato e acidificante, utilizando-se comerciais (Labtest Diagnóstica S.A.).
O volume urinário total diário foi estimado dividindo-se as excreções urinárias diárias de creatinina pelos valores observados de concentração de creatinina na urina (Valadares Filho & Valadares, 2001). A excreção urinária diária de creatinina foi estimada a partir da proposição de 24,05 mg/kg de peso vivo (PV) de creatinina (Chizzotti et al., 2008).
A excreção total de derivados de purina (PT) foi calculada pela soma das quantidades de alantoína e ácido úrico excretados na urina e da quantidade de alantoína excretada no leite. As purinas absorvidas (PA, mmol/dia) foram calculadas a partir da excreção de PT (PT, mmol/dia), por meio da equação PA = (PT – 0,512* PV0,75)/0,85, em que 0,85 é a recuperação de purinas absorvidas como derivados de purinas (Verbic et al., 1990) e 0,512*PV0,75 a contribuição endógena para excreção de purinas obtida para vacas leiteiras (Gonzalez-Ronquillo et al., 2003).
12 0,83, a digestibilidade das purinas microbianas e 0,116, a relação N-purina:N total nas bactérias (Chen & Gomes, 1992).
O balanço de compostos nitrogenados (BN) foi obtido pela diferença entre o total de nitrogênio ingerido (Ning) e o total de nitrogênio excretado nas fezes (N-fezes), na urina (N-urina) e no leite (N-leite). A avaliação do nitrogênio total nas fezes e na urina foi feita segundo técnica descrita em Silva & Queiroz (2002).
A orçamentação parcial relacionada ao custo da alimentação (X) e ao retorno com a venda do leite (Y) foi realizada através das relações retorno absoluto (Y-X) e retorno relativo (Y)/(X), em que X foi obtido pela multiplicação do valor por kg de matéria seca da dieta (A) pelo consumo de matéria seca em kg por dia (B). Enquanto que Y foi calculado multiplicando-se o valor por litro de leite (C) pela produção de leite em kg por dia (D).
As variáveis foram analisadas segundo o modelo estatístico: Y ijkl = µ + Qi + Tj + (P/Q)ik + (V/Q)il + QxTij + eijkl, sendo:
Y ijkl = observação na vaca 1, no período k, submetida ao tratamento j, no
quadrado latino i; µ = constante geral;
Qi = efeito do quadrado latino i, sendo i = 1, 2 e 3; Tj = efeito do tratamento j, sendo j = 1, 2, 3 e 4;
14
" " " ""
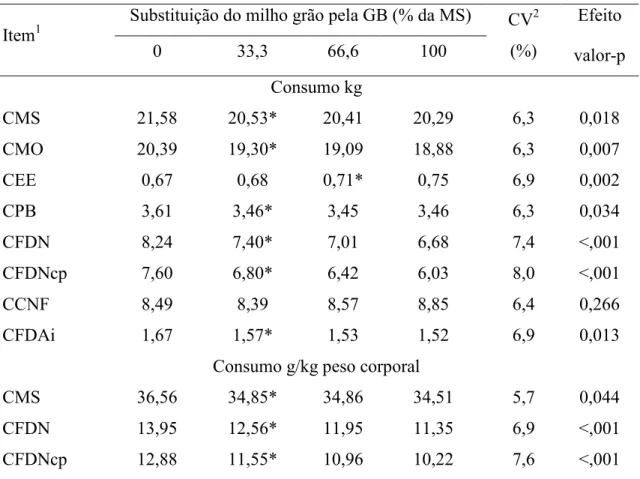
O consumo de MS foi reduzido (P<0,05) a partir de 33,3% de substituição de MG por GB na dieta. Os consumos de MO, PB, FDN, FDNcp e FDAi seguiram a mesma tendência, reduzindo (P<0,05) a partir de 33,3% de substituição, possivelmente, devido à redução do consumo de MS e da redução do teor fibra na dieta (Tabela 4). O consumo de EE aumentou (P<0,05) a partir do nível de 66% de substituição, comportamento que pode ser atribuído ao fato de que o teor dietético total de EE aumentou à medida que se elevou o nível de GB da dieta (Tabela 3). Todavia não houve efeito significativo (P>0,05) para o consumo de CNF.
Consumo de MS, FDN e FDNcp, expressos em g/kg de peso corporal, apresentaram redução a partir do nível de 33,3% de inclusão de GB em substituição ao MG nas dietas, acompanhando a tendência observada para o consumo em kg/dia (Tabela 4).
Neste contexto, pode-se dizer que, apesar da boa aceitação da GB pelos animais, a redução no consumo de MS pode estar associado ao teor de impurezas contido na GB. Dentre esses contaminantes, pode-se destacar o metanol, que na GB utilizada neste trabalho girou em torno de 8%, sendo que níveis elevados de metanol são de particular preocupação, devendo seu teor na glicerina bruta ser inferior a 0,5% (Donkin & Doane, 2007).
15 energia (NRC, 2001). Evidências empíricas para o metabólico mostra que o potencial produtivo do animal, que afeta sua capacidade de utilizar os nutrientes, interage com o equilíbrio dos nutrientes absorvidos para regular o consumo (Illius & Jessop, 1996). Todavia esse efeito de redução do consumo não foi verificado quando o glicerol foi incluído em substituição ao milho grão em dietas de vacas em lactação nos níveis 3,6% e 15% da MS (Khalil et al., 1997 e Donkin & Doane, 2007), respectivamente.
Tabela 4 – Consumos diários de constituintes em função do nível de glicerina bruta na dieta
Item1
Substituição do milho grão pela GB (% da MS) CV2 (%)
Efeito valor-p
0 33,3 66,6 100
Consumo kg
CMS 21,58 20,53* 20,41 20,29 6,3 0,018
CMO 20,39 19,30* 19,09 18,88 6,3 0,007
CEE 0,67 0,68 0,71* 0,75 6,9 0,002
CPB 3,61 3,46* 3,45 3,46 6,3 0,034
CFDN 8,24 7,40* 7,01 6,68 7,4 <,001
CFDNcp 7,60 6,80* 6,42 6,03 8,0 <,001
CCNF 8,49 8,39 8,57 8,85 6,4 0,266
CFDAi 1,67 1,57* 1,53 1,52 6,9 0,013
Consumo g/kg peso corporal
CMS 36,56 34,85* 34,86 34,51 5,7 0,044
CFDN 13,95 12,56* 11,95 11,35 6,9 <,001
CFDNcp 12,88 11,55* 10,96 10,22 7,6 <,001
Médias seguidas por (*) indicam o nível de inclusão a partir do qual se observa diferença em relação ao tratamento controle (nível zero) pelo teste de Williams (P<0,05).
1
MS = Matéria seca; MO = Matéria orgânica; PB = Proteína bruta; EE = Extrato etéreo; FDN = Fibra em detergente neutro; FDNcp = Fibra em detergente neutro corrigida para cinzas e proteína; CNF =
16 Apesar da digestibilidade da MS, MO, PB, CNF e o teor de NDT (% da MS) terem aumentado (P<0,05) a partir de 66,6% de substituição do MG pela GB, os consumos de MS, MO digeridos e de NDT não foram influenciados significativamente (P>0,05) pela substituição do MG por GB nas dietas. A digestibilidade do EE aumentou significativamente (P<0,05) a partir do nível de 33,3% de substituição, influenciando o consumo de EE digestível, que aumentou (P<0,05) a partir do mesmo nível. Enquanto que o consumo de componentes digeridos da PB reduziu (P<0,05) a partir do nível de 33,3% de substituição do MG pela GB da dieta (Tabela 5).
Embora sem haver significância (P>0,05) da digestibilidade do FDNcp, em termos absolutos, observa-se uma redução de 6,20 unidades percentuais no nível de 100% de substituição do MG pela GB em relação ao nível zero, que pode estar relacionado com a redução significativa (P<0,05) do consumo de componentes digeridos do FDNcp a partir de 33,3% de substituição (Tabela 5).
O menor consumo de componentes digeridos apresentado pela FDNcp pode ser atribuído ao aumento da proporção de CNF na dieta e ao menor tempo de permanência da digesta no ambiente ruminal, visto que a degradação da FDNcp é lenta e altamente influenciada pelo tempo de contato com as enzimas bacterianas.
A redução do teor de fibra decorrente da inclusão de GB nas dietas e conseqüente aumento do CNF (Tabela 3) podem estar relacionados com o maior teor de NDT (% da MS) com o aumento da GB, embora o consumo do NDT não ter se apresentado significativo (P>0,05) com a inclusão de GB nas dietas.
17 e a digestibilidade ruminal, síntese microbiana ruminal e digestibilidade total dos nutrientes em novilhos.
Tabela 5 – Coeficientes de digestibilidade total, teores de nutrientes digestíveis totais (NDT) e consumo de componentes digeridos em função dos níveis de substituição de milho grão pela glicerina bruta com 85% de glicerol nas dietas
Itens Substituição do milho grão pela GB (% da MS) CV%¹ Efeito valor-p
0 33,3 66,6 100
%
CDMS 61,00 62,05 63,93* 66,51 3,5 <,001
CDMO 62,37 63,39 65,12* 67,54 3,5 <,001
CDEE 42,64 59,55* 67,71 78,96 27,9 0,008
CDPB 65,94 65,84 68,82* 72,10 3,7 <,001
CDFDNcp 48,07 45,95 43,36 41,87 12 0,144
CDCNF 75,35 76,99 79,93* 82,45 3,8 <,001
NDT(% da MS) 60,80 62,39 64,24* 66,81 4,7 0,001
kg/dia
CMSD 13,15 12,68 13,06 13,78 6,4 0,050
CMOD 12,70 12,17 12,44 12,02 6,4 0,141
CPBD 2,37 2,27* 2,37 2,54 5,5 0,002
CEED 0,28 0,41* 0,48 0,60 26,8 <,001
CFDNcpD 3,62 3,09* 2,81 2,60 15 0,003
CCNFD 6,46 6,42 6,85* 7,31 7,6 0,004
CNDT 13,11 12,74 13,12 13,83 6,9 0,070
Médias seguidas por (*) indicam o nível de inclusão a partir do qual se observa diferença em relação ao tratamento controle (nível zero) pelo teste de Williams (P<0,05).
¹
Coeficiente de variação.
18 milho pode ser substituído por glicerol sem ajustes para o teor de energia, embora, o valor energético do glicerol bruto seja susceptível de ser menor do que o glicerol puro e deve ser ajustado para o conteúdo energético de impurezas.
A adição de fontes lipídicas em dietas de ruminantes pode acarretar em redução no consumo de MS (Vargas et al., 2002). No entanto, o aumento do teor de EE nas dietas decorrentes da elevação dos níveis de GB, bem como o aumento do consumo de EE digestível (P<0,05) não influenciou o nível energético das dietas, assim como reportado por Osborne et al. (2009), que não encontraram efeito significativo da suplementação energética sobre o consumo de energia de vacas leiteiras.
Na Tabela 6 encontram-se os valores observados e exigências diárias de proteína e energia para vacas lactantes com peso corporal médio de 600 kg, produção média diária de 30 kg/dia, com 3,5% de gordura e com 24 semanas de lactação, segundo o NRC (2001).
Talela 6 – Valores observados e exigências de proteína bruta (PB) e nutrientes digestíveis totais (NDT) segundo o NRC (2001), de vacas lactantes com 600 kg de peso corporal, com 24 semanas de lactação, produzindo 30 kg/dia com 3,5% de gordura
Itens Exigências Substituição do milho grão pela GB (% da MS) 1
0 33,3 66,6 100
PB (kg/dia) 3,30 3,61 3,46 3,45 3,46
Diferença 0,31 0,16 0,15 0,16
NDT (kg/dia) 12,50 13,11 12,74 13,12 13,83
Diferença 0,61 0,24 0,62 1,33
1
GB = glicerina bruta (84 % glicerol).
19 ocorrida no presente experimento, não foi causada por uma deficiência energética para os animais.
A produção de leite (PL) e PL corrigida para 3,5% de gordura (PLC) apresentaram redução (P<0,05) a partir do nível de 66,6% de substituição do MG pela GB, enquanto que não houve efeito (P>0,05) dos níveis de GB nas dietas sobre a composição do leite, relação entre o teor de proteína e gordura do leite e eficiência alimentar dos animais (Tabela 7).
Tabela 7 – Produção de leite, composição do leite e eficiência na produção em função do nível de substituição do milho grão por glicerina bruta na dieta
Item Substituição do MG pela GB (% da MS)² CV²
(%)
Efeito valor-p
0 33,3 66,7 100
Leite (kg/d) 31,14 29,77 28,42* 28,14 7,3 0,017
Leite com 3,5% gordura (kg/d) 31,05 31,09 29,40* 27,97 9,1 0,045
Lactose (%) 4,09 4,08 4,00 4,05 4,1 0,469
Proteína bruta (%) 3,14 3,21 3,24 3,22 5 0,552
Gordura (%) 3,55 3,86 3,84 3,49 11 0,089
Proteína:gordura 0,90 0,86 0,86 0,92 10,6 0,278
Extrato seco total (%) 11,92 12,29 12,23 11,93 3,3 0,091 Extrato seco desengordurado (%) 8,36 8,43 8,39 8,44 2,9 0,961
PLC/CMS¹ 1,46 1,52 1,50 1,37 12,4 0,168
Médias seguidas por (*) indicam o nível de inclusão a partir do qual se observa diferença em relação ao tratamento
controle (nível zero) pelo teste de Williams (P<0,05); 1 Produção diária de leite com 3,5% de gordura/consumo
diário de MS. 2 MG = milho grão, GB = glicerina bruta (84% de glicerol); Coeficiente de variação (CV).
20 A redução na síntese de N microbiano ruminal pode ter contribuído para redução da produção, por ocasionar reduções no fluxo de aminoácidos na glândula mamária. A proteína α-lactoalbumina, sintetizada na glândula mamária a partir de aminoácidos livres absorvidos na circulação sanguínea, apresenta papel essencial na atuação catalítica do complexo enzimático lactose sintase (Fonseca, 1995). Neste sentido, reduções no fluxo de aminoácidos de origem microbiana possivelmente reduziriam a síntese de componentes do leite na glândula mamária (Gennadij et al., 2000)
Donkin & Doane (2007), não encontraram efeito significativo (P>0,10) na produção e composição do leite, em vacas alimentadas com níveis de até 15% de glicerol em base da MS, adicionado em substituição ao MG nas dietas. Trabalhos onde menores níveis de inclusão de glicerol (<10% na MS) foram utilizados também não encontraram diferença significativa sobre a produção de leite (Khalil et al., 1997, DeFrain et al., 2004 e Al Bodarski al., 2005). Todavia, foram utilizados baixos níveis de inclusão de glicerol no desenvolvimento desses trabalhos, e o glicerol utilizado se apresentava na forma pura (99,5% de glicerol), diferindo da GB (84% de glicerol) utilizada no presente trabalho.
Não houve efeito sobre a variação de peso corporal, apresentando valores médios de 0,68, 0,38, 0,70 e 0,47 kg/dia, para as proporções de 0%, 33,3%, 66,6% e 100% de substituição do MG pela GB, respectivamente.
21 80% dos aminoácidos absorvidos no intestino delgado em ruminantes (Storm & Ørskov, 1983).
A quantidade total de proteína microbiana no intestino delgado depende da disponibilidade de nutrientes e da eficiência de uso desses nutrientes pelas bactérias ruminais. Segundo Russel et al. (1992), variações na síntese microbiana são associadas a mudanças no suprimento de energia e proteína ao animal hospedeiro, sendo que o fornecimento de energia é o primeiro fator limitando o crescimento microbiano ruminal (Valadares Filho et al., 2010).
Apesar da substituição do MG pela GB não afetar a disponibilidade ruminal de proteína bruta, essa substituição pode ter ocasionado redução da disponibilidade de energia e de esqueletos de carbono para a síntese de PBmic nas dietas com maiores níveis de inclusão de glicerina. Segundo Krehbiel (2008), o glicerol pode tomar três destinos no rúmen, incluindo passagem (13%), fermentação (44%) e absorção (43%), sendo que, mais da metade do glicerol consumido não é fermentado no rúmen, desta forma haveria uma redução de substrato para a síntese microbiana nas dietas com glicerina quando comparado ao milho grão.
22 A concentração de glicose sanguínea (mg/dL) não apresentou efeito significativo (P>0,05) com a inclusão de GB (Tabela 8). Segundo Allen et al. (2005), o propionato é o principal precursor de glicose para os ruminantes, no entanto o possível aumento da produção de propionato decorrente da fermentação do glicerol, não influencio significativamente (P>0,05) os níveis de glicose sanguínea, podendo-se concluir assim, que os níveis energéticos das dietas se mantiveram constantes para os animais.
A concentração de nitrogênio uréico no plasma (NUP) expressa em mg/dL, não apresentou resposta significativa (P>0,05) com a inclusão de GB nas dietas. Os níveis de concentração de nitrogênio no plasma acompanharam a tendência das demais variáveis relacionadas ao perfil de nutrição protéica, sendo que embora o coeficiente de digestibilidade da PB tenha aumentado significativamente (P<0,05) enquanto que o consumo de PB digestível apresentou uma redução significativa (P<0,05) em relação à inclusão de GB nas dietas (Tabela 5), os níveis de PB das dietas se mantiveram constante (Tabela 3). Porém, os valores de NUP encontram-se dentro da faixa considerada adequada de balanceamento de energia e proteína, de 10 a 17 mg/dL (Broderick, 1995; Moore & Varga, 1996; Jonker et al., 1998; Ferguson, 2001).
23 substituição de MG por GB na dieta não afetou significativamente (P>0,05) o balanço de nitrogênio (BN) expresso em g/dia e porcentagem do Ningerido (Tabela 8).
Contudo, salienta-se que não ocorreu valor negativo médio para o BN em nenhuma das proporções de substituição do MG pela GB, indicando que o consumo de N permitiu atender as exigências de N dos animais.
Os resultados deste estudo indicam claramente que GB é um ingrediente valioso para a alimentação de vacas leiteiras em lactação. A GB pode ser incluída como um macro ingrediente em dietas para vacas em lactação, e apesar de influenciar o consumo e produção de leite a inclusão do GB em substituição ao milho é uma estratégia Tabela 8 – Síntese e eficiência de síntese de nitrogênio microbiano ruminal (Nmic), teor de
glicose sanguínea e indicadores de metabolismo de nitrogênio em função do nível de substituição do milho grão pela glicerina bruta na dieta
Item¹ Substituição do milho grão pela GB (% da MS) CV² (%)
Efeito valor-p
0 33,3 66,6 100
Nmic, g/dia 310,12 265,58* 271,81 270,94 19,7 0,040
gPBmic/kgNDT g/dia 145,90 125,82* 124,70 108,16 22,0 0,016
NUP, mg/dL 17,07 15,54 14,85 15,16 11,3 0,099
Glicose Sanguínea mg/dL 60,27 57,41 54,08 58,00 14,9 0,129 Ningerido, g/dia 579,06 553,88* 553,37 554,57 6,3 0,034
Nurina g/dia 173,60 195,89 200,03 219,80 22,5 0,459
Nleite, g/dia 153,06 147,82 139,48* 145,37 6,1 <,001
Nfezes, g/dia 190,69 192,24 169,46* 151,30 11,0 0,001
BN, g/dia 61,08 17,92 46,80 43,59 30,3 0,281
BN/Ningerido % 10,06 3,06 8,63 7,38 6,7 0,278
Nleite/Ningerido % 26,35 26,82 25,18* 25,86 6,49 0,028
24 alternativa para formulação de dietas para vacas lactantes quando o milho não possui um preço favorável.
Na Tabela 9 são apresentados dados de uma orçamentação parcial da produção de leite em função dos níveis de substituição do MG pela GB nas dietas, onde são levados em consideração apenas gastos com alimentação, e receita provinda da venda de leite. Nessa demonstração foram usados dados de consumo e produção do presente experimento e valores médios de mercado. Os preços dos ingredientes em reais por kg (R$/kg) foram: 0,10; 0,33; 0,30; 0,57; 0,28; 1,30; 1,20; e 0,90 para silagem de milho, milho grão, glicerina bruta, farelo de soja, farelo de trigo, glúten de milho, uréia e mistura mineral, respectivamente.
¹Retorno relativo com o preço da glicerina bruta a R$ 0,18/kg.
O retorno relativo, encontrado dividindo-se o ganho por dia pelo gasto com alimentação pode ser considerado constante até o nível de 33% de substituição do MG pela GB na dieta, sendo que ocorreu uma diminuição de apenas R$0,02. No entanto, Tabela 9 – Orçamentação parcial da produção de leite em função dos níveis de substituição
do milho grão pela GB na dieta
Item Níveis de substituição de milho por glicerina bruta (% da MS)
0% 33% 66% 100%
R$/kg de MS 0,43 0,43 0,44 0,44
Consumo de MS kg/dia 21,58 20,53 20,41 20,29
Gasto com alimentação 9,17 8,83 8,89 8,94
R$/litro de leite 0,60 0,60 0,60 0,60
Produção de Leite L/dia 31,14 29,77 28,42 28,14
Ganho R$/dia 18,68 17,86 17,05 16,88
Retorno absoluto 9,51 9,03 8,16 7,95
Retorno relativo 2,04 2,02 1,92 1,89
25 fazendo-se uma simulação com o preço da GB a R$ 0,18/kg, encontramos um valor constante para o retorno relativo para todos os níveis de substituição do MG pela GB.
O aumento da produção de biodiesel, resultando na oferta de GB, combinada com a concorrência pela utilização do milho por outras espécies animais, pode justificar o uso de GB na alimentação de gado. Embora existam questões em relação à composição da GB, sua utilização pode se tornar viável em até 33,3% de inclusão em substituição ao MG com base na MS. Devendo-se tomar cuidado com os níveis de metanol contidos na GB, que podem ser prejudiciais a saúde dos animais.
26
"+ "
27
,
ABUGHAZALEH, A.A.; EL-NOR, S.A.; BABU, R. The effect of replacing corn with glycerol on rumen fermentation and fiber digestibility. - & , v.86, E-Suppl.2, p.474, 2008.
ALLEN, M. S., BRADFORD, B. J.Metabolic regulation of feed intake in cattle: a conceltual model. - & #87: 3317-3334, 2009.
ALLEN, M. S., BRADFORD, B. J., HARVATINE, K. J. The cow as a model to study food intake regulation. ! '" & . 25:523-547, 2005.
BERGNER, H., KIJORA, C., CERESNAKOVA, Z. SZAKACS, J. 1995. In vitro studies on glycerol transformation by rumen microorganisms. .# . . 48:245-256, 1995
AL BODARSKI, R., WERTELECKI, T. BOMMER, F., GOSIEWSKI, S. The changes of metabolic status and lactation performance in dairy cows under feeding TMR with glycerin (glycerol) supplement at periparturient period# - & / ".
$ ! " ", Animal Husbandry, 8:1- 9, 2005
BRODERICK, G.A. Use of milk urea as an indicator of nitrogen utilization in lactating dairy cows. USDA. Agriculture Research Service. US Dairy Forage Research Center, 1995. " . "0122p.
CARVALHO, G. R. Leite: Por que olhar para o Brasil? In: MilkPoint. 2007. Disponível em: www. http://milhpoint.com.br.
CASALI, A.O.; DETMANN, E.; VALADARES FILHO, S.C. et al. Influência do tempo de incubação e do tamanho de partículas sobre os teores de compostos indigestíveis em alimentos e fezes bovinas obtidos por procedimentos ! " "
1 , v.37, n.2, p.335-342, 2008.
CHEN, X.B.; GOMES, M.J. Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives - an overview of technical details.
, 2 . Rowett Research Institute.
Aberdeen, UK. (occasional publication). 1992. 21p.
28 CHIZZOTTI, M.L., VALADARES FILHO, S.C., VALADARES, R.F.D. et al. Consumo, digestibilidade e excreção de uréia e derivados de purinas em vacas de diferentes níveis de produção de leite. ! " " 1 . v.36, n.1, p.138-146, 2007 DEFRAIN, J.M.; HIPPEN, A.R.; KALSCHEUR, K.F. et al. Feeding glycerol to transition
dairy cows: Effects on blood metabolites and lactation performance. - & %
, v.87, p.4195-4206, 2004.
DONKIN, S.S.; DOANE, P. % " & $ % ". In: Tri-State Dairy Conference, p.97-103, 2007. Disponível em: www. http://tristatedairy.osu.edu/proceedings.htm.
DROUILLARD, J.S. Glycerin as a feed for ruminants: using glycerin in high-concentrate diets. - & , v.86, E-Suppl.2, p.392, 2008.
FERGUSON, J.D. Milk urea nitrogen. & 2 . / ! %, 2001, http://cahpwww.vet.upenn.edu/mun/mun_info.html (10-01-2009).
FISHER, L.J.; ERFLE, J.D.; LODGE, G.A. et al. Effects of propylene glycol or glycerol supplementation of the diet of dairy cows on feed intake, milk yield and composition, and incidence of ketosis. - & 0 v.53, p.289–296, 1973.
FISHER, L.J.; ERFLE, J.D.; SAUER, F.D. Preliminary evaluation of the addition of glucogenic materials to the rations of lactating cows. - &
0v.51, p.721–727, 1971.
FONSECA, F. A. Fisiologia da Lactação. Centro de Ciências Agrárias. Departamento de Zootecnia. Viçosa-MG: UFV. 1995. 137p.
GENNADIJ, C. C.; DANFAER, A.; CANT, J.P. Simulation analysis of substrate utilization in the mammary gland of lactating cows. - & % " ., n.67, p171-188,2000.
GONÇALVES, V.L.C.; PINTO, B.P.; MUSGUEIRA, L.C. et al. Biogasolina: Produção de ésteres da glicerina. In: I Congresso da Rede Brasileira de Tecnologia de Biodisel, 2006, Brasília. "###Brasília: MCT/ABIPTI, 2006. v.2. p.14 -19.
GONZÁLEZ-RONQUILLO, M.; BALCELLS, J.; GUADA, J.A.; et al. Purine derivative excretion in dairy cows: Endogenous excretion and the effect of exogenous nucleic acid supply. - & % , v.86, n.4, p.1282-1291, 2003.
29 HESS, B.W.; LAKE, S.L.; GUNTER, S.A. Using glycerin as a supplement for forage-fed
ruminants. - & , v.86, E-Suppl.2, p.392-393, 2008.
IDF – INTERNATIONAL DAIRY FEDERATION. 4. 3# &
3& 0 5 " & . 5 & 6 & 6
" ". Bruxelas: 1996. 12p. (IDF Standard 141 B).
ILLIUS, A. W. and JESSOP, N. S. Metabolic constraints on voluntary intake in ruminants.
- & # 74: 3052-3062, 1996.
JOHNSON, M.M.; PETERS, J.P. Technical Note: An improved method to quantify nonesterified fatty acids in bovine plasma. - & % , v.71, p.753-756, 1993.
JOHNSON, R.B. The treatment of ketosis with glycerol and propylene glycol.
7 . v.44, p.6–21, 1954.
JONKER, J.S.; KOHN, R.A.; ERDMAM, R.A. Using milk urea nitrogen to predict nitrogen excretion and utilization efficiency in lactating dairy cows. - & %
, v.81, p.2681-2692, 1998.
KHALILI, H., VARVIKKO, T., TOIVONEM, V., HISSA, K., SUVITIE, M. The effects of added glycerol or unprotected free fatty acids or a combination of the two on silage intake, milk production, rúmen fermentation and diet digestibility in cows given grass silage based diets. $ #Finland. 6:349–362, 1997.
KREHBIEL, C.R. Ruminal and physiological metabolism of glycerin. Symposium: Ruminant Nutrition: Glycerin as a Feed for Ruminants, - & . Vol. 86, p-392, 2008.
LICITRA, G.; HERNANDEZ, T.M.; VAN SOEST, P.J. Standardization of procedures for
nitrogen fractionation of ruminant feeds. . $%, v.57,
n.4, p.347-358, 1996.
LIN, E. C. C. Glycerol utilization and its regulation in mammals. Annu. Rev. . . 46:765–795, 1977.
MERTENS, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: collaborative study. - &
, v.85, p.1217-1240, 2002.
MOORE, D.A.; VARGA, G. BUN and MUN: Urea nitrogen testing in dairy cattle.
5 $ 7 %, v.18, n.6, p.712-721, 1996.
NATIONAL RESEARCH COUNCIL - NRC. 8 " & % . 7.
30 OSBORNE, V. R., OGONGO, N, E.,CANT, J. P. SWANSON, K. C. McBRIDE. Effects of supplementing glycerol and soybean oil in drinking water on feed and water intake, energy balance, and production performance of periparturient dairy cows. - &
% , 92: 698-707, 2009
RÉMIND, B., SOUDAYR, E., JOUANY, J. P. In vitro and in vivo fermentation of glycerol by rumen microbes. . Technol. 41:121–132, 1993 RUSSELL, J. P., O’CONNOR, C, D., FOX, D. G. et al. A net carbohydrate and protein
system for evaluating cattle diets. I. Ruminal fermentation. - &
. 70: 3562-3577, 1992.
SCHRÖDER, A., SÜDEKUM, K. H. Glycerol as a by-product of biodiesel production in diets for ruminants. In New Horizons for an Old Crop. Proc. 10th Int. 5 "
$ #, Canberra, Australia. N. Wratten and P. A. Salisbury, ed.241: 26–29,1999 SILVA, D.J.; QUEIROZ, A.C. 9 " ": * " 8 ; " <$ ".
3.ed. Viçosa: UFV, 2002. 235p.
SKLAN, D.; ASHKENAZI, R.; BRAUN, A. et al. Fatty acids, calcium soaps of fatty acids and cottonseeds fed to high yielding cows. - & % , v.75, p.2463-2472, 1992.
SNIFFEN, C.J., O’CONNOR J.D., VAN SOEST, P.J. et al. A net carbohydrate and protein system for evaluating cattle diets: carbohydrate and protein availability. - &
0v.70, n.12, p.3562-3577, 1992.
STATISTICAL ANALISYS SYSTEM - SAS. = " >" $ . v.2, 4.ed. Cary: 1989. 846p.
STORM, E.; ØRSKOV, E. R. The nutritive value of rumen microorganisms in ruminant. 1. Large-scale isolation and chemical composition of rumen microorganisms. ".
- & , v.50, p. 463-470, 1983.
VARGAS, L. H., LANA, R. P., JHAM, G. N. et al. Adição de lipidios na ração de vacas leiteiras: parâmetros fermentativos ruminais, produção e composição do leite. ! "
" 1 , V.31, n.1, p.522-529, 2002.
VALADARES FILHO, S.C.; et al.. ? $@ " " A ; " 5 " A " 6 # 2.ed. – Viçosa : UFV, DZO, 2010, 30-31p.
31 VERBIC, J.; CHEN, X.B.; MACLEOD, N.A. et al. Excretion of purine derivatives by ruminants. Effect of microbial nucleic acid infusion on purine derivative excretion by steers. - $ , v.114, n.3, p.243-248, 1990.
WEISS, W.P Energy prediction equations for ruminant feeds. In: CORNELL NUTRITION CONFERENCE FOR FEED MANUFACTURERS, 61., 1999. Proceeding… Ithaca: Cornell University, p. 176-185. 1999.