Iranian Journal of Basic Medical Sciences
ijbms.mums.ac.ir
Squid ink polysaccharide reduces cyclophosphamide‐induced
testicular damage via Nrf /ARE activation pathway in mice
Xiaoyan Le , Ping Luo , Yipeng Gu , Yexing Tao , (uazhong Liu *
College of Science, Guangdong Ocean University, Zhanjiang, China
A R T ) C L E ) N F O A B S T R A C T Article type:
Short communication Objective(s):via restoration of antioxidant ability in our previous investigations. This study investigated roles of Cyclophosphamide CP toxicity on testis was hampered by squid ink polysaccharide S)P Nrf /ARE signal pathway in testis of treated mice.
Materials and Methods: Male Kunming mice were employed to undergo treatment with S)P and/or CP.
Protein levels of Nrf , keap‐ , histone deacetylase (DAC , quinone oxidoreductase NQO‐ , and heme oxygenase (O‐ and phosphorylation level of protein kinase C PKC in testis were evaluated by Western blotting.
Results: Data showed that S)P elevated expressions of NQO‐ and (O‐ genes, two downstream target
molecules of Nrf , via activating Nrf to play preventive roles on CP‐treated testis, and further discovered that upstream regulators of Nrf , keap‐ , (DAC , and PKC, were concerned with the regulation of Nrf .
Conclusion: These results suggest that S)P could effectively weaken CP‐associated testicular damage
via Nrf /ARE signal pathway. Article history:
Received: Oct , Accepted: May ,
Keywords: Cyclophosphamide Mice
Nrf /ARE Squid )nk Polysaccharides Testis
►Please cite this article as:
Le X, Luo P, Gu Y, Tao Y, Liu (. Squid ink polysaccharide reduces cyclophosphamide‐induced testicular damage via Nrf /ARE activation pathway in mice. )ran J Basic Med Sci ; : ‐
Introduction
Chemotherapy is known to be a critical remedy for tumors, and cyclophosphamide CP is used as a common chemotherapeutic drug for cancer and immunosuppression for nephritic disorders and lupus erythematosus, while negative roles of CP on male reproductive functions have been widely recognized for many years, which result in dose reduction of antineoplastic drugs and leads to a decline of chemotherapeutic effects ‐ . (ence, weakening toxic damage of CP on normal tissues and increasing dose of CP for elevation of therapeutic effects in patients have been taken as purposes for developing cytoprotectors to be used as assistant drugs for chemotherapy.
)n recent years we proved squid ink polysaccha‐ rides S)P , a type of glycosaminoglycan with a unique structure ‐[ GlcA ‐ GalNAc ‐ ‐Fuc ]n–
, , to have antioxidative activities and
chemoprotective roles invitro and/or invivo ‐ .
Our previous investigations revealed that the chemoprotective activities of S)P were observed in
liver, lung, heart, kidney, marrow, and testis of model animals exposed to CP through improving
antioxidant abilities of the organs , which implies
that S)P can prevent testis from CP‐induced oxidative stress damage.
Nrf NF‐E ‐related factor is an important
transcription factor that binds to AREs that are
important gene regulatory elements of many phase Τ
drug‐metabolizing/detoxification enzymes as well as cellular defensive enzymes and regulates anti‐oxidative stress and plays a concernful role in eliminating intracellular superfluous reactive oxygen species ROS
and improves antioxidant ability . Some bioactive
substances have been proved to activate Nrf to relieve or prevent diseases correlated to oxidative stress, such
as astaxanthin and curcumin , . (owever, it
is still unknown whether S)P impairs CP‐caused testicular damage through activating Nrf to upregulate
expression of antioxidative enzyme genes and phase Τ
enzyme genes subjected to maintaining redox equilibrium. )n this study, mice were employed to
prove the hypothesis invivo.
Materials
and
Methods
PreparationofSIP
Live squids purchased from a local aquatic products market were sacrificed to harvest fresh ink sacs that were then stored at ‐ for future use.
According to our previous methods , ink collected
from sacs thawed at and was suspended with p( . PBS, and was then ground and ultrasonically treated. The resultant ink solution was stored at
for hr and was then centrifuged at g for
hr at . The supernatant was subjected to
enzymolysis with % papain in PBS p( . at for hr, and was then mixed with a / volume liquid mixture of chloroform and n‐butanol v/v, / followed by stirring for min on a magnetic
stirrer plate. After centrifugation at g for
min, the supernatant was re‐digested with papain, the digestion process was performed twice. S)P in the resulting supernatant was precipitated with four volumes of absolute alcohol, and was subjected to freeze‐drying in a vacuum. Solid powder S)P was stored at before use.
Animalexperimentalscheme
Following habituation for week, sexually mature male Kunming mice purchased from the Experimental Animal Centre of Guangdong Medical College were allocated to four groups ten mice per group : a control group, a CP‐treated group, a S)P‐ treated group, and a co‐treated group S)P and CP . The S)P dose was mg/kg body weight, once a day for a continuous ten week period, and the CP dose was mg/kg body weight, once a week again for a continuous ten week period .
Westernblottinganalysis
Testes were collected from mice that were sacrificed by breaking neck vertebrae and were quickly
cleaned with ice‐cold normal saline. Testis was minced and homogenated in ice‐cold normal saline with a glass homogenizer. Testis homogenate was centrifugated
at rpm for min at . The harvested
supernatant was denatured in protein sample buffer for min in boiling water. After SDS‐PAGE, protein was transferred to nitrocellulose membrane and then probed with monoclonal antibody, against Nrf
Cell Signaling , ‐actin Cell Signaling , keap‐ Cell Signaling , (DAC Cell Signaling , p‐PKC Cell Signaling , NQO‐ abcam , or (O‐ abcam , which would be captured by the secondary antibody conjugated with horseradish peroxidase Santa Cruz . The membrane was visualized with SuperSignal West Pico chemiluminescence detection system Pierce . These protein levels were normalized with ‐actin.
Statisticalanalysis
Value is expressed as mean±standard error. Statistical analysis was performed using ANOVA by JMP . software. Differences were separated by Duncan’s multiple range test. Significance was
considered at P< . or P< . .
Results
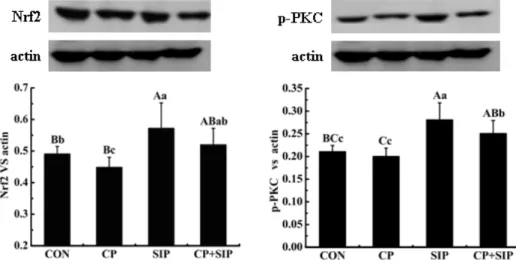
To determine roles of Nrf in mice testis treated with S)P and/or CP, in this study we detected the expression level of Nrf gene in testis and the results were presented in Figure . The data showed that CP obviously decreased protein content of Nrf in testes of model mice. Our data declared apparently an insignificant result that PKC was not phosphorylated in testes of mice exposed to CP. (owever in S)P‐ treated mice degree of PKC phosphorylation was markedly improved, meanwhile expression level of Nrf gene was increased sharply in testes, the similar positive effects of S)P were observed in testes of CP‐ administered mice.
Figure1. Contents of Nrf protein and activation level of protein kinase C PKC in testes of mice. Values with different lowercase
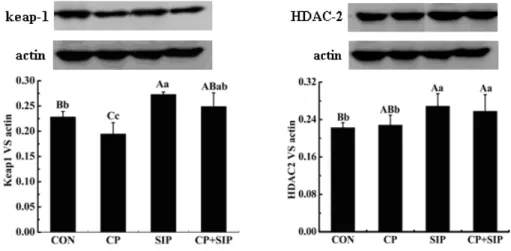
Figure2. Contents of keap‐ and histone deacetylase (DAC proteins in testes of mice. Values with different lowercase superscripts
mean significant difference P< . , those with different capital letters are extremely different P< .
)n comparison with CP‐treated mice, a marked increase of expression of keap gene in testes from S)P‐exposed and co‐treated mice. For a regulatory enzyme that can stabilize Nrf protein through deacetylation, expression content of (DAC gene was not affected by CP in mice testes, but obvious increases of the protein content of (DAC were observed under stimulation of S)P that is presented in mice of both S)P‐ and co‐treated groups Figure . As shown in Figure , data showed that CP did not obviously change expression level of (O‐ gene but sharply declined the content of NQO‐ protein in testes, and that S)P not only markedly improved contents of (O‐ and NQO‐ in normal mice, also significantly increased levels of the two proteins in CP‐treated mice.
Discussion
Acrolein, a cyclophosphamide’s metabolite induces body to produce superfluous ROS that cause tissues to
undergo oxidative stress and/or apoptosis . To
avoid the damages inducedby CPorotheroxidative
stressors, cells develop a series of signal regulative mechanisms to keep intracellular redox equilibrium , Nrf /ARE is one of their important pathways.
Under normal conditions, Nrf is kept fairly low level in cytoplasm through binding to keap or
rapid degradation. (owever stimulation by drugs or other oxidative stressors can elevate dissociative content of Nrf , release from keap or up‐regulating expression of Nrf gene, which enters the nucleus to bind ARE of the downstream genes and then to
stimulate expression of the genes, such as phase Τ
enzyme genes, antioxidant enzyme genes and protein kinase genes, which results in immediate detoxication and maintaining intracellular homeos‐ tasis. Nrf widely distributed in any of various organs plays a concernful role in protection of the organs, deletion or active disturbance of the factor should lead cells to more sensitivity to oxidative
stress . This study revealed that CP reduced
protein content of Nrf in testis which was in
accordance with Tripathi’s report . Presently,
itisknown that Nrf isphosphorylatedbyPKC to be
Figure3. Contents of heme oxygenase (O‐ and quinone oxidoreductase NQO‐ proteins in testes of mice. Values with different
activated , . Although PKC failed to be phosphorylated in CP‐exposed mice testis, it was activated under treatment together with S)P.
Nrf was regulated by other upstream molecules besides PKC, such as keap and (DAC . Keap is identically considered to be an inhibitor of Nrf . Under normal conditions, Nrf is degraded through ubiquitin‐ proteasome pathway after the protein is bound in the
Neh domain by keap , and is separated from
keap for reasons such as phosphorylation of PKC on
Nrf , , to play biological roles. )n this paper we
found S)P increased content of keap protein, which may be originated from positive feedback regulation by increase of Nrf protein content, which would be subjected to binding and decreasing dissociative Nrf protein in cytoplasm and combined Nrf protein bound
to AREs in nucleus . The results suggested that S)P
not only improved expression of keap gene in testes of normal mice, but also positively affected contents of the protein in testes of chemotherapeutic mice, which was subjected to resisting expression increase of Nrf gene. (DAC is an enzyme that can deacetylate Nrf protein to harvest stabilized protein that can activate the
downstream effective molecules , , such as
antioxidative enzymes and phase Τ enzymes.
According to the data, CP did not change (DAC content in testes, but S)P significantly improved the expression level of (DAC gene in both S)P‐ and co‐ treated mice.
Although our data have proved upstream regulatory factors of Nrf were affected by stimulation of S)P, the regulation was just acted on Nrf protein. This paper discovered that S)P interfered with contents of Nrf protein as determined by Western blotting, which was similar to present reported antioxidants, such as green
tea , astaxanthin and curcumin , , and
the metallic element zinc . The increase in Nrf
protein may be originated from improvement of expression of Nrf gene caused by S)P.
From above mentioned information, activated Nrf protein plays roles through activating downstream effective molecules,which contain many enzymes, such as heme oxygenase (O‐ and quinone oxidoreductase NQO‐ , which are two important enzymes in antioxidation and detoxification. Background researchers reported that Nrf binds to ARE through using bZ)P basic leucine zipper, a domain found in many DNA binding eukaryotic proteins to form dimers with Maf a transcription factor and then activate the downstream genes, (O‐ and NQO‐ ; expression levels of the two genes were positively
correlated with the Nrf gene ‐ . This study
showed that S)P effectively changed effects of CP on expression levels of (O‐ and NQO‐ genes in testes.
Summarily, in this study we firstly found that S)P employed the Nrf /ARE signal pathway to activate downstream target genes, (O‐ and NQO‐ , to exert preventive roles against CP‐induced damage on mice
testis, and that PKC acted as an upstream regulator that participated in the regulative process on Nrf . )n addition, two other upstream molecules, keap and (DAC , were also vital proteins in the process of regulation of S)P on activity of Nrf . Thus, we can conclude that S)P triggers Nrf /ARE signal cascades via regulating PKC, keap and (DAC , whereas we are still ignorant of the accurate regulative mechanisms of S)P on PKC, keap and (DAC , which would be an important content in our future research.
Conclusion
This study showed that S)P reduces CP‐induced testicular damage via Nrf /ARE activation pathway in mice. To our knowledge this study is the first to report that intervention of S)P on CP‐mediated testicular damage is connected with Nrf /ARE signalling pathway, which suggests that Nrf /ARE may be an important signalling pathway that is used by S)P to weaken chemotherapeutic drugs‐induced damage of male reproductive ability. Further investigation is necessary to learn more details about the regulatory mechanisms, the results would be helpful to developing S)P as a potential chemotherapeutic adjuvant drug.
Acknowledgment
The work was supported by the National Natural
Science Foundation of China . The results
reported in this paper were part of a student thesis.
Conflict
of
interests
The authors declare no potential conflict of interests.
References
. Emadi A, Jones RJ, Brodsky RA. Cyclophosphamide and cancer: golden anniversary. Nat Rev Clin Oncol
; : ‐ .
. Pryzant RM, Meistrich ML, Wislon G, Brown B, McLaughlin P. Long‐term reduction in sperm count after chemotherapy with and without radiation therapy for non‐hodgkin’s lymphomas. J Clin Oncol
; : – .
. Elangovan N, Chiou TJ, Tzeng WF, Chu ST. Cyclophosphamide treatment causes impairment of sperm and its fertilizing ability in mice. Toxicology
; : ‐ .
. Takaya Y, Uchisawa (, Narumi F, Matsue (. )llexins A, B, and C from squid ink shoule have a branched
structure. Biochem Biophys Res Commun ;
: ‐ .
. Chen S, Xu J, Xue C, Dong P, Sheng W, Yu G, Chai W. Sequence determination of a non‐sulfated
glycosaminoglycan‐like polysaccharide from
melanin‐free ink of the squid Ommastrephes bartrami by negative‐ion electrospray tandemmass spectrometry and NMR spectroscopy. Glycoconjugate
J ; : ‐ .
cyclophosphamide‐associated testicular damage in
mice. Bratisl Lek Listy ; : ‐ .
. Tang Q, Zuo T, Lu S, Wu J, Wang J, Zheng R, Chen S, Xue C. Dietary squid ink polysaccharides ameliorated the intestinal microflora dysfunction in mice
undergoing chemotherapy. Food Funct ; :
‐ .
. Zuo T, Cao L, Xue C, Tang QJ. Dietary squid ink polysaccharide induces goblet cells to protect small intestine from chemotherapy induced injury. Food
Funct ; : ‐ .
. Zuo T, Cao L, Li X, Zhang Q, Xue C, Tang QJ. The Squid )nk Polysaccharides Protect Tight Junctions and Adherens Junctions from Chemotherapeutic )njury in the Small )ntestinal Epithelium of Mice. Nutr
Cancer ; : – .
. Luo P, Liu (Z. Antioxidant ability of squid ink polysaccharides as well as their protective effects on
DNA damage invitro. Afr J Pharm Pharmacol ;
: ‐ .
. Li Y, Paonessa JD, Zhang Y. Mechanism of chemical
activation of Nrf . Plos One ; : e .
. Tripathi DN, Jena GB. Astaxanthin inhibits cytotoxic and genotoxic effects of cyclophosphamide.
Toxicology ; : ‐ .
. Balogun E, (oque M, Gong P, Killeen E, Green CJ,
Foresti R, et al. Curcumin activates the haem
oxygenase‐ gene via regulation of Nrf and the
antioxidant‐responsive element. Biochem J ;
: ‐ .
. Garg R, Gupta S, Maru GB. Dietary curcumin
modulates transcriptional regulators of phaseΣand
phaseΤenzymes in benzo[a]pyrene‐treated mice:
mechanism of its anti‐initiating action. Carcinogenesis
; : ‐ .
. Liu F, Li XL, Lin T, (e DW, Wei G(, Liu J(,etal.
The cyclophosphamide metabolite, acrolein, induces cytoskeletal changes and oxidative stress in Sertoli
cells. Mol Biol Rep ; : ‐ .
. Miao W, (u L, Scrivens PJ, Batist G. Transcriptional regulation of NF‐E p ‐related factor NRF expression by the aryl hydrocarbon receptor‐xenobiotic response element signaling pathway direct cross talk
between phaseΣand phaseΤ drug‐metabolizing
enzymes. J Biol Chem ; : ‐ .
. Stepkowski TM, Kruszewski MK. Molecular cross‐ talk between the Nrf /KEAP signaling pathway,
autophagy, and apoptosis. Free Radic Biol Med ;
: ‐ .
. Tripathi DN, Jena GB. Astaxanthin intervention
ameliorates cyclophosphamide‐induced oxidative
stress, DNA damage and early hepatocarcinogenesis in
rat: role of Nrf , p , p and phase‐Τ enzymes. Mutat
Res ; : ‐ .
. (uang (C, Nguyen T, Pickett CB. Phosphorylation of Nrf at Ser‐ by protein kinase C regulates antioxidant response element‐mediated transcription. J Biol Chem
; : ‐ .
. Niture SK, Jain AK, Jaiswal AK. Antioxidant‐
induced modification of )Nrf cysteine and PKC‐
δ‐mediated phosphorylation of Nrf serine are
both required for stabilization and nuclear translocation of Nrf and increased drug resistance. J
Cell Sci ; : ‐ .
. Keum YS, Yu S, Chang PP, Yuan X, Kim J(, Xu C, etal.
Mechanism of action of sulforaphane: inhibition of p mitogen‐activated protein kinase isoforms contributing to the induction of antioxidant response element– mediated heme oxygenase‐ in human hepatoma hepG
cells. Cancer Res ; : ‐ .
. Velichkova M,(asson T. Keap regulates the
oxidation‐sensitive shuttling of Nrf into and out of the nucleus via a Crm ‐dependent nuclear export
mechanism. Mol Cell Biol ; : ‐ .
. Sun Z, Chin YE, Zhang DD. Acetylation of Nrf by
p /CBP augments promoter‐specific DNA binding
of Nrf during the antioxidant response. Mol Cell Biol
; : ‐ .
. Mercado N, Thimmulappa R, Thomas CM, Fenwick PS, Chana KK, Donnelly LE, Biswal S, )to K, Barnes PJ. Decreased histone deacetylase impairs Nrf activation by oxidative stress. Biochem Biophys Res
Commun ; : ‐ .
. Forney GB, Morre DJ, Morre DM. Oxidative stress reduced by a green tea concentrate and capsicum
combination: synergistic effects. J Diet Suppl ;
: ‐ .
. Li B, Cui W, Tan Y, Luo P, Chen Q, Zhang C, etal.
Zinc is essential for the transcription function of Nrf
in human renal tubule cells in vitro and mouse
kidney invivo under the diabetic condition. J Cell Mol
Med ; : ‐ .
. Kobayashi M, Yamamoto M. Nrf –Keap regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv
Enzyme Regul ; : ‐ .
. Yang J(, Shin BY, (an JY, Kim MG, Wi JE, Kim YW,
etal. )sorhamnetin protects against oxidative stress
by activating Nrf and inducing the expression of its
target genes. Toxicol Appl Pharm ; : ‐ .
. Zhang C, Su ZY, Khor TO, Shu L, Kong AN. Sulforaphane enhances Nrf expression in prostate cancer TRAMP C cells through epigenetic regulation.