cry1ac
gene by biolistic, without the use of markers
De Guglielmo-Cróquer, Z.
a, Altosaar, I.
b, Zaidi, M.
band Menéndez-Yuffá, A.
a*
aLaboratorio de Clonación y Genética Vegetal, Instituto de Biología Experimental,
Facultad de Ciencias, Universidad Central de Venezuela – UCV, Calle Suapure, Colinas de Bello Monte, 1041A Caracas, Venezuela
bUniversity of Ottawa, 40 Marie Curie, Ottawa ON KIN6N5, Canada
*e-mail: andrea.menendez@ciens.ucv.ve (With 3 figures)
Received November 3, 2008 – Accepted June 1, 2009 – Distributed May 31, 2010
Abstract
The transformation of coffee plantlets with the cry1ac gene of Bacillusthuringiensis was achieved by biolistic using
either the whole pUBC plasmid or only the ubi-cry1ac-nos genetic cassette. The cry1ac gene was inserted into
cof-fee plants in order to confer resistance to the leaf miner Leucopteracoffeella, an insect responsible for considerable
losses in coffee crops. Bearing in mind that the genetic cassettes used for this study lack reporter genes and/or selec-tion marker genes, the parameters for the transformaselec-tion procedure by biolistic were previously standardised with a plasmid carrying the gus reporter gene. The presence of the cry1ac gene in young plantlet tissues was determined by
PCR, Southern blot and reverse transcription-PCR. Our results show that the obtainment of viable coffee plantlets, transformed by bombardment with the cry1ac gene and without selection markers nor reporter genes, is feasible.
Keywords: coffee, biolistic, genetic cassette, cry1ac gene, transformation without markers.
Transformação do café (
Coffea arabica
L. cv. Catimor)
com o gene
cry1ac
por bombardeamento, sem usar marcadores
Resumo
A transformação das plântulas de café com o gene cry1ac de Bacillusthuringiensis foi realizada por biobalística,
utilizando todo o pUBC plasmídeo ou só o cassete genético UBI-cry1ac-nos. O gene cry1ac foi inserido no cafeeiro
a fim de conferir resistência à folha mineiro Leucopteracoffeella, um inseto responsável por perdas consideráveis
nas culturas de café. Tendo em conta que ao plasmídeo e ao cassete genético utilizados para este estudo faltam genes repórteres e/ou de seleção, os parâmetros para o processo de transformação por biobalística foram previamente pa-dronizados com um plasmídeo transportando o gene repórter gus. A presença do gene cry1ac em tecidos de jovens
plântulas foi determinada por PCR, Southern blot e transcrição reversa-PCR. Nossos resultados mostram que a obten-ção de plântulas de café, transformado por bombardeamento com o gene cry1ac sem genes de seleção genética nem
repórteres é viável.
Palavras-chave: café, biobalística, cassette genético, gene cry1ac, transformação sem marcadores.
1. Introduction
Coffee stands among the first five crop products sold worldwide and the number of people that live from this crop is estimated at 125 million, including some 25 million producers and processors (O´Brien and Kinnaird, 2003; ICO, 2007). In 2006/07 world production reached over 125 million bags, an increase of 14% compared to crop year 2005/06, when output was 110 million bags (ICO, 2007). Insects have been blamed as the most diverse and numerous plagues affecting this crop, and the lepidop-teran Leucoptera coffeella (leaf-miner) and the
coleop-teran Hypothenemus hampei (borer) stand out as
thur-ingiensis, which codifies for a crystalline protein that is specifically toxic to lepidopteran larvae (Guerreiro et al., 1998). Using Agrobacteriumtumefaciens or A. rhizogenes
several genes have been inserted into coffee plants, such as bar (for resistance to the FinaleTM herbicide), gus
(re-porter gene for β-glucuronidase), nptII (resistance to kanamycin), hpt (resistance to hygromycin), csr1
(resist-ance to the herbicide chlorosulphuran), rolB (a gen from A. rhizogenes) and cry1ac (resistance to lepidopterans).
Hatanaka et al. (1999), Leroy et al. (2000), Perthuis et al. (2005), Ribas et al. (2006), Alpizar et al. (2006) and Kumar et al. (2006) have all reported regeneration of such transformed tissues. The genes gus, bar and nptII
have also been incorporated by electroporation; notable are the studies of van Boxtel et al. (1995) who described the transitory expression of the gus gene, Barton et al.
(1991) who transformed coffee-plant embryos with the
nptII gene, and Fernández-Da Silva and Menéndez-Yuffá
(2003) who reported the regeneration of transformed tissues with positive results for the GUS and PCR reac-tions for gus and bar genes, while da Cunha et al. (2004)
achieved the transformation of C. arabica using biolistics
with the nptII and gus genes. The insertion of the cry1ac
gene into coffee plants has been accomplished by trans-formation with A. tumefaciens (Leroy et al. 2000);
how-ever, the biolistic procedure has the advantage of simple construction vectors besides easier transformation proto-cols as complex plant-bacteria interactions are eliminated (Gray and Finer, 1993, cited by Ribas, et al. 2006).
Transformations by microparticle bombardment are usually carried out using whole plasmids, which carry se-quences that may negatively affect the transgen stability and its expression. Plant transformations have been de-scribed in recent years using genetic blocks assembled by the gene of interest flanked by the promotor and ending sequences, which in turn eliminate, or at least minimise, those negative effects; also this procedure reduces the number of foreign sequences introduced into the plant ge-nome (such as reporter genes, selection markers and genes for resistance to bacterial antibiotics), all of which could represent an advantage regarding biosafety (Fu et al., 2000; Breitler et al., 2002; Vianna et al., 2004). In view of this, the present study was aimed at the transformation of arabica coffee by means of biolistic using a whole plasmid or a genetic block with the cry1ac gene, in order to
intro-duce resistance to the leaf-miner. It is noteworthy that the conditions of the procedure as followed (including kind of explant, helium pressure, distance between the micropar-ticle carrier and explant, shot time and post-bombardment recovery medium) were determined in a previous work (De Guglielmo et al., 2007, in preparation), which in turn facilitated the transformation with the cry1ac gene, as the
genetic construction used for the present study lacked both reporter genes and selection markers.
2. Materials and Methods
2.1. Plant material
Torpedo-shape somatic embryos of C. arabica cv. Catimor were used. This material was obtained by the in-duction methods for somatic embryogenesis of coffee as
described by Menéndez-Yuffá et al. (1994). Conditions in the growth room were: illumination 12 µmol.m–2.s at a
mean temperature of 24 °C.
2.2. Genetic constructions
We used the pUBC plasmid (approximately 7,000 bp), kindly donated by Dr. I. Altosaar (University of Ottawa, Canada), which is based on the pGEM 4Z vector (Promega) containing a synthetic version of the cry1ac gene under control of the ubiquitine pro-moter of corn and the nopaline synthetase terminator (Sardana et al., 1996). This genetic block (approximately 4,200 bp), was introduced in the HindIII site of the base
vector which also contains the resistance gene to ampi-cillin, an antibiotic used as a selection agent for trans-formed bacteria. Plasmid DNA was isolated using the Wizard® Plus Midipreps DNA Purification System Kit
(Promega), and following its instructions, from E. coli
bacterial strain DH5α transformed with the pUBC plas-mid as described by Chung et al. (1989). This DNA was digested with the HindIII enzyme (Invitrogen),
follow-ing the manufacturer`s recommendations, and subjected to electrophoresis in 0.8% agarose gel (Promega), con-taining TAE 1X buffer (4.84 g Tris base, 1.14 mL glacial acetic acid and 2 mL EDTA 0.5 M at pH 8, final volume 1 L) at 80 V. The gel was then stained with ethidium bro-mide and the resulting bands were analysed using the Gel Doc 2000 Gel Documentation System (BIORAD). The ubi-cry1ac-nos genetic block was also used, lib-erated from the whole plasmid by digestion with the
HindIII enzyme (Invitrogen). The band concerned
(ap-proximately 4,200 bp) was separated from the agarose gel and the DNA from this block was purified by cen-trifuging on glass wool, followed by precipitation with absolute ethanol.
2.3. Biolistic-mediated transformation
Bombardments of somatic torpedo embryos of
C. arabica cv. Catimor with the whole pUBC plasmid
or with the ubi-cry1ac-nos genetic block were carried
out using a low-pressure helium pistol manufactured by the Instituto Venezolano de Investigaciones Científicas (IVIC), based on Vain et al. (1963). Relevant procedures were as in Klein et al. (1987, 1988) with the following standard conditions: helium pressure 70 psi, shot time 0.1 seconds, distance between the microparticle carrier and explants 14 cm, vacuum pressure 21 mm.Hg–1;
re-covery medium was the solid medium for germination without hormones described by Hermoso-Gallardo and Menéndez-Yuffá (2000).
2.4. Evaluation of transformation
2.4.1. PCR analysis
Plant DNA was extracted according to Doyle and Doyle (1990) with some modifications to reduce the amount of foliar tissues used, as well as DNA extrac-tion time and length of PCR tests (Fernández-Da Silva and Menéndez-Yuffá, 2003). This material was used for PCR tests with specific primers for the cry1ac gene,
un-der conditions as described by Sardana et al. (1996). The primer pair Ac-f (ATGGACAACAACCCAAACATC) and Ac-r (TCATGTCGTTGAATTGAATAC) were used to amplify a sequence of 416 bp. The fragments were visualised by electhrophoresis on 2% agarose gels, TAE 1X buffer and ethidium bromide staining (Chandler and Vaucheret, 2001; Jana et al., 2004).
2.4.2. Southern blot hybridization
This procedure was done with the DIG DNA Labelling and Detection Kit (Roche), according to its indications to attain a non-radioactive label of the probe. Roughly 30 µg of plant DNA from each sample were treated with HindIII enzyme. The ubi-cry1ac-nos genet-ic block was used as probe; it was obtained by enzymatgenet-ic digestion of pUBC plasmid DNA with HindIII, followed
by labelling with digoxigenin.
2.4.3. Reverse transcription-PCR (RT-PCR)
This was carried out using the Super Script II re-verse transcriptase (Invitrogen). Total RNA of the bom-barded plantlets was used, extracted with the Plant RNA Purification Reagent (Invitrogren), and the synthesis of cDNA was performed with random primers (Invitrogen). PCR was then carried out with specific primers for the
cry1ac gene, as detailed above for PCR analysis.
2.5. Statistical analyses
Non-parametric Kruskal-Wallis tests were applied to compare results of bombardments with the whole plasmid or with the genetic block containing the cry1ac
gene, regarding numbers of bombarded embryos, surviv-ing embryos, regenerated plantlets, analysed plantlets
and transgene detection by means of the various molecu-lar techniques employed (see above).
3. Results
3.1. Bombardments with the pUBC plasmid or the genetic block
Twelve torpedo embryos were used for each bom-bardment, corresponding to the area covered by the shots under our experimental conditions (Table 1). The embryos usually showed a dark brown colouration af-ter bombardment, probable due to mechanical damages. Out of the 12 embryos bombarded in each assay, four or five (roughly 42%) died within two weeks of the treat-ment and showed necrosis, further darkening and tissue softening. Two months after the bombardments some of the surviving embryos showed the emergence of second-ary somatic embryos, which died off shortly afterwards. At two and a half months the development of cotyledons was noted in an average of six surviving embryos from each of the replicates. One cotyledon was removed from each for PCR analysis with the specific primers for the
cry1ac gene.
3.2. PCR
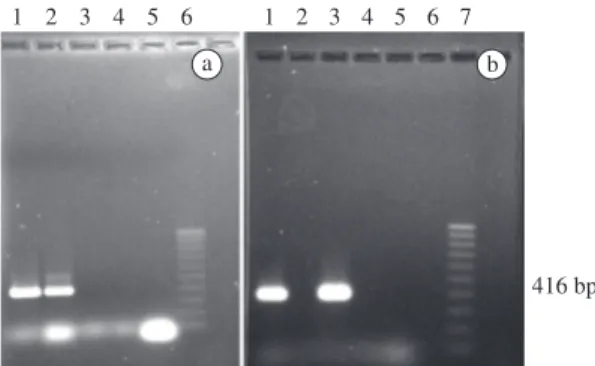
The primers used revealed an amplification product of 416 bp. For the transformation with the pUBC plas-mid 51 plantlets were analysed, of which 7 (14%) were positive for the PCR test (Figure 1). Of these seven, one died shortly afterwards. Of the embryos bombarded with the cry1ac-sequence carrier block, all the regenerated
plantlets (19/19) were examined by PCR, but only one (5%) gave positive results.
3.3. Southern blot hybridisation
The probe used for this assay was prepared with the genetic block containing the cry1ac sequence. Positive control was represented by DNA from the pUBC plas-mid and negative control by DNA from untransformed coffee plants. Controls and PCR-positive plantlets DNA were digested with the HindIII enzyme, which frees the
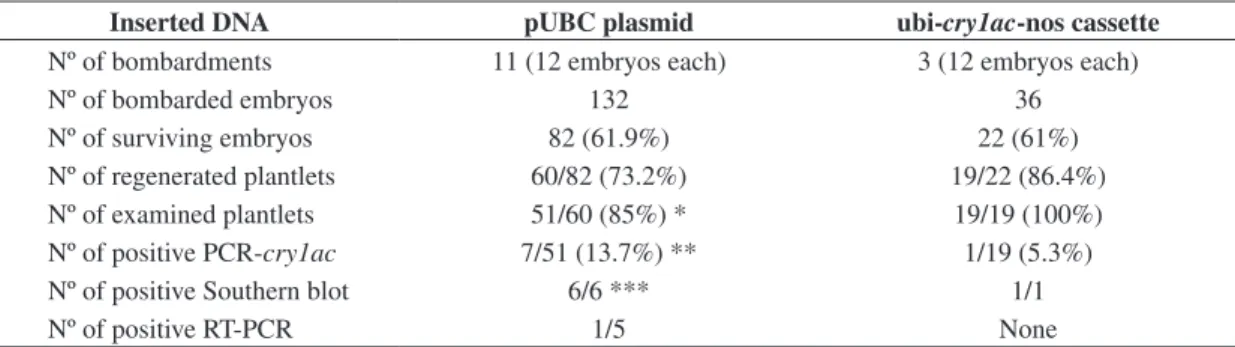
Table 1. Results of the bombardments of torpedo somatic embryos of coffee with the plasmid or the genetic cassette.
Inserted DNA pUBC plasmid ubi-cry1ac-nos cassette
Nº of bombardments 11 (12 embryos each) 3 (12 embryos each)
Nº of bombarded embryos 132 36
Nº of surviving embryos 82 (61.9%) 22 (61%)
Nº of regenerated plantlets 60/82 (73.2%) 19/22 (86.4%) Nº of examined plantlets 51/60 (85%) * 19/19 (100%) Nº of positive PCR-cry1ac 7/51 (13.7%) ** 1/19 (5.3%)
Nº of positive Southern blot 6/6 *** 1/1
Nº of positive RT-PCR 1/5 None
cry1ac fragment together with the promoter and the ter-minator. Results are shown in Figure 2. The positive con-trol showed a strong band whereas no hybridisation took place between the probe and the negative control, nor with the plantlets which were negative for PCR tests with
cry1ac primers (results not shown). The seven surviving
plantlets positive for the later test, of which six belonged to the whole-plasmid bombardment and one to the sin-gle genetic cassette, hybridised with the probe. Of these plantlets (including the latter) three revealed a single band presenting a similar migration observed in the posi-tive control, corresponding to the cry1ac gene fragment
of 4,200 bp (Figure 2a). One of them, resulting from the whole-plasmid assay, showed a mixed hybridisation pat-tern expressed as one band similar to the positive control (4,200 bp) and another heavier (7,500 bp approximately) but less intense one (Figure 2b, lane 2). In the three re-maining samples from the whole-plasmid bombardments a complex pattern of bands was noticed, represented by
two heavier bands (7,500 and 10,000 bp approximate-ly) than that of the positive control (Figure 2b, lane 4) (Spiral, 1993).
3.4. Reverse transcription-PCR
This test was performed on six surviving plantlets positive by PCR and by Southern blot, as of the remain-ing seven, one showed transgenic rearrangements by hybridisation when tested by Southern blot but had to be discarded due to contamination. Positive control was the pUBC plasmid DNA while negative controls corre-sponded to non-transformed coffee cDNA and the reac-tion mixture. As shown in Figure 3 (lane 5) only a single sample, from the bombardments with the whole plasmid and which revealed a single band of the expected size by Southern blot, has the expected band of 416 bp cor-responding to the cry1ac gene as revealed by PCR. This
band also shows in the positive control but it is absent from the negative controls.
3.5. Statistical analyses
Based on the non-parametric test used, no significant differences (p > 0.05) were obtained between the results of survivorship, regeneration and PCR detection in the bombardments with the whole plasmid or with the ge-netic cassette.
Figure 1. PCR results with specific primers for the cry1ac gene.1a and 1b: positive controls (pUBC DNA); the ex-pected amplification product of 416 bp is evident. 2 to 4a and b: DNA from the bombarded tissues (lanes 2a and 3b show the positive samples). 5a and b: negative control (DNA from non-transformed coffee). 6a and 7b: Molecular weight markers (100 bp, Invitrogen).
a b
Figure 2. Southern blot hybridisation with the probe based on the ubi-cry1ac-nos block, of DNA from coffee plantlets positive to PCR with cry1ac primers. 3a and 5b: positive controls (pUBC DNA). 4a and 1b: negative controls (DNA from non-transformed coffee). 1a, 2b and 4b: DNA from 3/7 coffee plantlets positive to PCR. 2a and 3b: DNA from plantlets which gave negative results for cry1ac primers PCR. All DNA were digested with HindIII enzyme.
a b
Discussion
The present work achieved the transformation of
C. arabica plants with the cry1ac gene to confer
resist-ance to the leaf-miner, by means of biolistic bombard-ments using whole plasmids or genetic blocks. Table 1 summarises the results of such bombardments. The use of simple genetic blocks or cassettes built with promoter-sequence of interest-terminator has not been previously reported in the literature concerning transformation of coffee plants. This technique for genetic modifications has been used by Fu et al. (2000) and Breitler et al. (2002) to transform rice by means of biolistic, without a significant loss of transformation efficiency nor af-fecting the responses of bombarded explants regarding survival or regeneration, in comparison with the use of whole plasmid DNA. Those authors also pointed out that the rice plants transformed by genetic cassettes exhibited simple integration patterns and low frequencies of trans-gene rearrangements (less than those observed with the use of whole plasmid). This favours the stable expression of the transgene and improves biosafety as less foreign rearranged sequences become inserted in the bombarded tissues. This also reduces or eliminates the possible risk of horizontal or vertical transfer of genes for the resist-ance to antibiotics or herbicides, used as selection mark-er genes in the transformation procedure.
Recently, Vianna et al. (2004) transformed bean plants by biolistic using a genetic cassette containing the bar gene, and their findings agree with those of Fu et al. (2000) and Breitler et al. (2002). The use of the whole plasmid could cause plasmid-plasmid recombina-tion events brought about by sequences rich in A-T (i.e. highly recombinogenic) on the backbone of the transfor-mation vectors, which in turn might be involved in plas-mid multimerization events (Fu et al., 2000). Our own findings did not reveal any statistically significant dif-ferences between responses of tissues bombarded about survivorship, regeneration and detection by PCR, either with the whole plasmid or the genetic block, in agree-ment with other results published elsewhere. Anyhow, the advantage of using limited genetic blocks appears to support transgenic stability expression and inheritance over a long range.
Our hybridisation results tested by Southern blot evidenced that seven plantlets of coffee from the bom-barded and regenerated embryos selected by PCR incor-porated the cry1ac gene. The test also showed that of
seven plantlets, three had a single band agreeing with the genetic block, indicating that no rearrangements had oc-curred. Additionally, four plantlets showed extra bands or did not have the one corresponding to the block, as was shown in Figure 2 (lines 2b and 4b); one possible cause could be the transgene rearrangements.
The six surviving plantlets selected by PCR and which were also positive by Southern blot hybridisation, were examined by RT-PCR. As was shown in Figure 3, only a single sample had the expected 416 bp band on the
PCR test with the cry1ac primers. This suggests that five of the six plantlets positive for PCR and Southern blot hybridisation (including the one resulting from the bom-bardment limited to the genetic cassette) do not express the gene cry1ac. Only the plantlet positive for RT-PCR
would possibly show any resistance to the leaf-miner by its expression of the cry1ac protein. Anyhow, we do not disregard further assessments of the plantlets which gave negative results for RT-PCR tests (these negative results could be associated with molecular technique or DNA detection system sensibility). Their tissues ought to be propagated, while examining their proteins and perform-ing bioassays to expose plantlets to the insect larvae.
Several hypothesis could explain the lack of expres-sion at mRNA level; one of them could be the sensibil-ity of the test, and as reported by Matzke and Matzke (1995), Hiei et al. (1994) and Kumpatla et al. (1997), this can be due to genetic rearrangements, gene silencing by insertion in transcriptionally inactive or highly repetitive regions, or by methylation.
The transformation obtained in this study was per-formed without the use of markers of selection or report genes. This is a very important feature for biosafety in a plant with a product intended for human consumption. However, the marker genes are important to establish the procedure of transformation, for this reason, the pa-rameters for transformation by bombardment were pre-viously established using the plasmid pCAMBIA3201 (De Guglielmo et al, in preparation) which contains the reporter gene gus. The best conditions were selected
ac-cording to the survivorship and the expression of gus. It is remarkable, that even though gus is considered
tran-sient, the expression of this gene was observed seventeen months after bombardment of torpedo shape embryos. Bombardments with pCAMBIA3201 were made in par-allel with those of the gene cry1ac, they were done as a control for the procedure, and to have a faster and easier procedure to follow the procedure of transformation, this control showed gus expression in all the experiments of
bombardments (results not presented).
According to the results, it is advisable to carry out new experiments in the future, to evaluate pre and post bombardment treatments, and also conditions for the re-generation of transformed products. It would also be in-teresting to improve the efficiency of secondary somatic embryogenesis in the bombarded material, to propagate the transformed material and also to diminish the risk of chimerism.
gene which showed expression at the mRNA level, with the further advantage that they contained neither reporter genes nor selection markers that could be expressed in the plants, a rather important aspect in view of the recent tendencies concerning biosafety in plant transformation research.
Acknowledgements — Thanks to FONACIT and to CDCH for financial support of this research project. We also thank Dr. Eva de García, Head of the Lab. of Biotecnología Vegetal, IBE-UCV; and Dr. Maira Oropeza for useful tips on the Southern blot methodology.
References
ALPIZAR, E., DECHAMP, E., ESPEOUT, S., ROYER, M., LECOULS, A., NICOLE, M., BERTRAND, B., LASHERMES, P. and ETIENNE, H., 2006. Efficient production of Agrobacterium rhizogenes-transformed roots and composite plants for studying gene expression in coffee roots. Plant Cell Reports, vol. 25, no. 9, p. 959-967.
BARTON, CR., ADAMS, TL. and ZAROWITZ, M., 1991. Stable transformation of foreign DNA into Coffea arabica plants. In Proceedings of the 14 International Conference on Coffee Science. San Francisco: ASIC. p. 460-464.
BREITLER, J., LABEYRIE, A., MEYNARD, D., LEGAVE, T. and GUIDERDONI, E., 2002. Efficient microprojectil bombardment-mediated transformation of rice using gene cassettes. Theoretical and Applied Genetics, vol. 104, no. 4, p. 709-719.
CHANDLER, V. and VAUCHERET, H., 2001. Gene activation and gene silencing. Plant Physiologyl., vol. 125, no. 1, p. 145-148.
CHENG, X., SARDANA, R., KAPLAN, H. and ALTOSAAR, I., 1998. Agrobacterium transformed rice plants expressing synthetic cry1A b and c genes are highly toxic to striped stem borer and yellow stem borer. Proceedings of the National Academy of Sciences, vol. 95, p. 2767-2772.
CHUNG, C., NIEMELA, S. and MILLER, R., 1989. One-step preparation of competent E. coli: transformation and storage of bacterial cells in the same solution. Proceedings of the National Academy of Sciences, vol. 86, no. 7, p. 2172-2175.
CUNHA, W., MACHADO, F., VIANNA, G., TEIXEIRA, J. and BARROS, E., 2004. Obtenção de plantas de Coffea arábica geneticamente modificadas por bombardeamento de calos embriogenicos. Boletim de Pesquisa e Desenvolvimento, vol. 73, p. 1-15.
DOYLE, JJ. and DOYLE, JL., 1990. Isolation of plant DNA from fresh tissue. Focus, vol. 12, no. 1, p. 13-15.
FU, X., DUC, L., FONTANA, S., BONG, B., TINJUANGJUN, P., SUDHAKAR, D., TWYMAN, R., CHRISTOU, P. and KHOLI, A., 2000. Linear transgene construct lacking vector backbone sequences generate low copy number transgenic plants with simple integration patterns. Transgenic Research, vol. 9, no. 1, p. 11-19.
GUERREIRO, O., DENOLF, P., PEFEROEN, M., DECAZY, B., ESKES, A. and FRUTOS, R., 1998. Susceptibility of the Coffee Leaf Miner (Perileucoptera spp) to B. thuringiensis -endotoxins: a model for transgenic perennial crops resistant
to endocarpic insects. Current Microbiology, vol. 36, no. 3, p. 175-179.
HATANAKA, T., CHOI, YE., KUSANO, T. and SANO, H., 1999. Transgenic plants of coffee Coffea canephora from embryogenic callus via Agrobacterium tumefaciens- mediated transformation. Plant Cell Reports, vol. 19, no. 2, p. 106-110.
HERMOSO-GALLARDO, L. and MENÉNDEZ-YUFFÁ, A., 2000. Multiplicación masiva el café (Coffea arabica L. cv. Catimor) mediante cultivo de suspensiones celulares embriogénicas. Acta Científica Venezolana, vol. 51, no. 2, p. 90-95.
HIEI, Y., OHTA, S., KOMARI, T. and KUMASHIRO, T., 1994. Efficient transformation of rice (Oriza sativa) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. The Plant Journal, vol. 6, no. 2, p. 271-282.
International Coffee Organization - ICO, 2007. Annual review 2006/2007. Available from: <http://dev.ico.org/documents/ review8e.pdf>. Access in: 22/03/2008.
JANA, S., CHAKRABORTY, CH. and NANDI, S., 2004. Mechanisms and roles of the RNA-based gene silencing. Electronic Journal of Biotechnology, vol. 7, no. 3, p. 321-332.
KLEIN, T., WOLF, E., WU, R. and SANFORD, J., 1987. High-velocity microprojectiles for delivering nucleic acids into living cells. Nature, vol. 327, p. 70-73.
KLEIN, T., GRADZIEL, T., FROMM, M. and SANFORD, J., 1988. Factors influencing gene delivery into Zea mays cells by high-velocity microprojectiles. Bio/Technology, vol. 6, p. 559-564.
KOBAYASHI, N., HORIKOSHI, T., KATSUYAMA, H., HANDA, T. and TAKAYANAGI, K., 1998. A simple and efficient DNA extraction method for plants, especially woody plants. Plant Tissue Culture and Biotechnology, vol. 4, no. 2, p. 76-80.
KRANZ, J., SCHMUTTERER, H. and KOCH, W., 1978. Diseases pests and weeds in tropical crops. Chichester: John Wiley & Sons. 670 p.
KUMAR, V., SATYANARAYANA, K., SARALA, S., INDU, E., GIRIDHAR, P., CHANDRASHEKAR, A. and RAVISHANKAR, G., 2006. Stable transformation and direct regeneration in Coffea canephora P ex. Fr. by Agrobacterium rhizogenes mediated transformation without hairy root phenotype. Plant Cell Reports, vol. 25, no. 3, p. 214-222.
KUMPATLA, S., TENG, W., BUCHHOLZ, W. and HALL, T., 1997. Epigenetic transcriptional silencing and 5-Azacytidine-mediated reactivation of a complex transgene in rice. Plant Physiology, vol. 115, no. 2, p. 361-373.
LEROY, T., HENRY, A., ROYER, M., ALTOSAAR, I., FRUTOS, R. and PHILLIPE, R., 2000. Genetically modified coffee plants expressing the B. thuringiensis cry1Ac gene for resistance to leaf miner. Plant Cell Reports, vol. 19, no. 4, p. 382-389.
MATZKE, M. and MATZKE, A., 1995. How and why do plants inactivate homologous (Trans)genes? Plant Physiology, vol. 107, no. 3, p. 679-685.
MCCABE, D. and CHRISTOU, P., 1993. Direct DNA transfer using electric discharge particle acceleration. Plant Cell Tissue Organ Culture, vol. 33, no. 3, p. 227-236.
MONDRAGÓN, M., SILVA, R. and RODRÍGUEZ, G., 2004. Estrategias para el manejo integrado del minador de la hoja y la broca del fruto del cafeto. Venezuela: FONAIAP. Available from: <http://www.ceniap.gov.ve/publica/divulga/fd60/broca. html>. Access in: 30/06/2005.
O’BRIEN, TG. and KINNAIRD, MF., 2003. Caffeine and conservation. Science, vol. 300, no. 5619, p. 587.
PERTHUIS, B., PRADON, J., MONTAGNON, C., DUFOUR, M. and LEROY, T., 2005. Stable resistance against the leaf miner Leucoptera coffeella expressed by genetically transformed Coffea canephora in a pluriannual field experiment in French Guiana. Euphytica, vol. 144, no. 3, p. 321-329.
RIBAS, A., PEREIRA, L. and VIEIRA, L., 2006.Genetic transformation of coffee. Brazilian Journal of Plant Physiology, vol. 18, no. 1, p. 83-94.
SÁGI, L., PANIS, B., SCHOOFS, H., SWENNEN, R. and CAMMUE, B., 1995. Genetic transformation of banana and plantain via particle bombardment. Nature Biotechnology, vol. 13, no. 5, p. 481-485.
SARDANA, R., DUKIANDJIEV, S., CHENG, X., COWAN, K. and ALTOSAAR, I., 1996. Construction and rapid testing of synthetic and modified toxin gene sequences cry1A b and c by expression in maize endosperm culture. Plant Cell Reports, vol. 15, no. 9, p. 677-681.
SILVA, RF. and MENÉNDEZ-YUFFÁ, A., 2003. Transient gene expression in secondary somatic embryos from coffee tissues electroporated with the genes gus and bar. Electronic Journal of Biotechnology, vol. 6, no. 1, p. 29-38.
SPIRAL, J., THIERREY, C., PAILLARD, M. and PETIARD, V., 1993. Obtention de plantules de Coffea canephora Pierre (robusta) transformées par Agrobacterium rhizogenes. Comptes Rendus de l’Académie des Sciences, vol. 316, p. 1-6. (serie III)
VAIN, P., KEEN, N., MURILLO, J., NEMES, C. and FINER, J., 1963. Development of the particle inflow gun. Plant Cell, Tissue and Organ Culture, vol. 33, no. 3, p. 237-246
VAN BOXTEL, J., BERTHOULY, M., CARASCO, C. and ESKES, A., 1995. Transient expresión of β-Glucuronidase following biolistic delivery of foreign DNA into coffee tissues. Plant Cell Reports, vol. 14, no. 12, p. 748-752.