Neuza dos Prazeres Lim a Teix eira
Neuza dos Prazeres Lim a Teix eira
Dissertation presented to obtain the Ph.D degree in Biology
Inst it ut o de Tecnologia Quím ica e Biológica | Universidade Nova de Lisboa
Oeiras,
March, 2014
Neuza dos Prazeres Lim a Teix eira
Dissert at ion present ed t o obt ain t he Ph.D degree in Biology
Inst it ut o de Tecnologia Quím ica e Biológica | Universidade Nova de Lisboa
Oeiras, March, 2014
Role in
Enterococcus faecalis
Neuza Teixeira, Constança Pomba, António Jacinto, Miguel Prudêncio
and Francisco Dionísio.
1 4
ndMarch 2 0 1 4
Second Edition, March 2 0 1 4
Stress by Antibiotics and Virulence of Enterococci Laboratory
Instituto de Tecnologia Química e Biológica (ITQB)
Universidade Nova de Lisboa
Financial Support from
Fundação para a Ciência e Tecnologia
(
FCT
)
iii
PhD Maria de Fátima Gonçalves Ribeiro dos Santos Silva Lopes
Auxiliary investigator at Instituto de Tecnologia Química e Biológica, Oeiras.
Co-supervisor:
PhD Michael S. Gilmore
Sir William Osler Professor at Harvard Medical School, USA
Examining Committee
PhD Miguel Prudêncio (Principal Examiner)
Investigator/ Group Leader at Instituto de Medicina Molecular (IMM), Lisboa.
PhD Constança Pomba (Principal Examiner)
Associate Professor at Faculdade de Medicina Veterinária da Universidade Técnica de Lisboa.
PhD António Jacinto
Principal investigator at Chronic Diseases FCM Nova (CEDOC), Lisboa.
PhD Francisco Díonisio
v
To my supervisor
vii I would like to express my sincere gratitude to everyone who directly or indirectly helped me through the development of this thesis. I would also like to acknowledge the institutes where I worked: ITQB, Shepens Research Eye Institute of Harvard Medical School and CEDOC.
Special thanks to my great supervisor, Maria deFátima Silva Lopes. This thesis is the result of great team work, without you I would never have developed this thesis, that´s why I dedicate this thesis to you, Fátima. I admire you professionally and personally, I learned a lot with you. During the 8 years I worked with you, you never let me down, you were always a great supervisor. Even when you were crossing difficult times you have never put me aside. You are a special person, a special friend, thanks for all our discussions about science, live conversations and all the laughter we had. For me, this PhD is not the end of our team work, it´s the beginning!
Thanks to my co-supervisor, Michael S. Gilmore for having received me in his laboratory and for having accepted to be my co-supervisor. It was a pleasure working in your lab and with your team. Thanks for believing in my abilities/skills and in my work and for helping me during the last four years. Thanks for the advice and wise words that you have always directed me.
Thanks to my collaborators, Kelli Palmer, Lynn Hancock, Jiro Nakayama and
Anna Zaidman-Remy who accepted to work with me and Fátima. We had
interesting discussions about my PhD work.
Thanks to all past colleagues of SAVE laboratory, especially to Sofia Santos,
Paulo Marujo and Teresa Braga for the discussions about science and for being my friends during that time. From the lab next door to: Beatriz, Marta and Rusa for the incredible moments and good laughter we had. Special thanks to my friends Paula Alves and Filipa Silva. Thanks for the nice lunches, dinners and happy moments! I believe our friendship will persist!
A special thanks to all present and past colleges from António Jacinto lab´s. All of you contributed for the success of my PhD. I spent great times with you and I learned a lot about Drosophila and Zebrafish. Thanks to Ana Roberto and Fernanda for always being ready to help me! Thanks to Lara for the great moments while sharing the desk, I really spend wonderful moments with you! Thanks for the help that you gave me in understanding the “Drosophila world”! Thanks to Ana Sofia for helping me with Drosophila protocols and for always being available to help. To Mariana, Maria, Angela, Carolina and Telmo for the great time we spent together in fly room and CEDOC´s sofas! Thanks to Marta Carapuço and Virginia for being great friends and giving me very wise advices, Thanks!!!
Thanks to all my PhD collegues for the nice moments during the PhD program classes, our dinners and parties. We spent great times together!! I made very good friends! Thanks to Margarida Saramago for being my friend and partner in congresses, we spent a great time together in Greece, I´ll never forget! Thanks to
ix moments of my PhD and of my personal life, you are a really good friend!
Thanks to all past and present InteraQB colleagues, we spent great times organizing the parties and SunSetSessions! A special thanks to João Damas, Barbara, Lia,
Fábio, Rui, Joana and both Filipa, through InteraQB we built an incredible team and we built a solid project that is a success in ITQB!
Um grande Obrigado às minhas amigas Ana Margarida Pardelha, Cláudia Xavier e
Rita Fidalgo por terem tido paciência para ouvir todas as minhas preocupações, reclamações e fúrias! Sem o vosso apoio seria difícil ultrapassar as fases menos boas do doutoramento. Sempre serão um grande suporte da minha vida, obrigado!
Um grande obrigado à minha irmã, cunhado e sobrinhos por ajudarem a não me sentir sozinha durante o meu doutoramento e me incluírem sempre nos seus planos!
Um especial obrigado aos meus Pais, por SEMPRE acreditarem em mim e SEMPRE me apoiarem, mesmo não percebendo bem o que eu fazia no laboratório. São os meus heróis, admiro-vos muito por serem tão especiais. Obrigado!
Um obrigado doce ao Ricardo Cesário por ter sido SEMPRE o meu suporte, o meu porto seguro! Pelo amor e carinho incondicional que sempre me deu e por SEMPRE acreditar em mim. Mesmo nos momentos em que estivemos longe, foste sempre a pessoa em que me deu a palavra e a força que mais precisava. És a pessoa que mais me entende, Amo-te!
xi The present thesis dissertation is the result of more than four years of research at the Stress by Antibiotics and Virulence of Enterococci (SAVE) laboratory from Instituto de Tecnologia Química e Biologica (Oeiras, Portugal); Tissue Morphogenesis and Repair laboratory from Chronic Diseases FCM Nova (Oeiras, Portugal) and Departments of Ophthalmology and Microbiology and Immunobiology from Harvard Medical School (Boston, USA), under supervision of Maria de Fátima Silva Lopes and Michael S. Gilmore.
The thesis is divided in six chapters. In Chapter I some general concepts of quorum sensing and Enterococcus genus are introduced. Particular attention is given to
Enterococcus faecalis pathogenesis correlated with quorum sensing Fsr system and
virulence factors it regulates. Additionally, the advantages of using Drosophila
melanogaster as a model organism to study host-pathogen interaction are described.
The Chapters II and III focus on an interesting phenomenon, which is the shutting off of the QS under certain circumstances. Chapter II focus on a particular diary strain,
E. faecalis LN68, previously reported to show incongruence between gelatinase
genotype and phenotype. We report all experiments performed to explain the reason of this incongruence. From this work a manuscript was published, which the author of this dissertation played a major contribution and is the first author. In Chapter III we describe work produced in order to understand the antagonistic effect of vancomycin, a cell-wall active antibiotic, on expression of fsr, gelE and sprE genes. This work resulted in a publication in which the author of this thesis is a co-author.
In Chapter IV we report experiments made to identify all genes regulated, directly and indirectly, by the Fsr system. To complete the study we also established
Drosophila as a good model to study Fsr virulence. This approach allowed the
xiii - Teixeira N, Santos S., Marujo P, Yokohata R.,Iyer V. , Nakayama J., Hancock LE, Serror P. and Maria de Fátima Silva Lopes (2012); The incongruent gelatinase
genotype and phenotype in Enterococcus faecalis are due to shutting off the
ability to respond to the gelatinase biosynthesis-activating pheromone (GBAP)
quorum-sensing signal; Microbiology,158, 519–528 (Doi: 10.1099/mic.0.055574-0).
- Teixeira N., Varahan S., Gorman Matthew J., Palmer L. K., Zaidman-Remy A., Yokohata R., Nakayama J., Hancock E. L., Jacinto A., Gilmore M. S., Maria de Fátima Silva Lopes (2013); Drosophila host model reveals new Enterococcus faecalis quorum-sensing associated virulence factors; PLoS One 8(5): e64740 (Doi: 10.1371/journal.pone.0064740).
- Ribeiro T., Teixeira N., Yokohata R., Nakayama J., Gilmore M.S. and Maria de Fátima Silva Lopes (2013); Transcriptomic study Reveals new pathways and
genes involved in Enterococcus faecalis V583 response to a therapeutic dose
of vancomycin, Archives of Microbiology, 4(5:3).(Doi: 10.3823/274).
- Teixeira N., Jacinto A. and Maria de Fátima Silva Lopes; Contribution of
melanization to Drosophila survival changes with E. faecalis V583 genomic
xv When Quorum Sensing (QS) was discovered it was realized that bacteria have a kind of “social life” and they cooperate and coordinate their activities on the bodies/environments they infect/live. Many bacteria only become dangerous to us when they sense that their numbers are high enough to overwhelm human defences. Only then they release their toxins and cause illness and death. Since this important discovery, many bacteria (both Gram negative and Gram positive) were identified as QS participants. In Gram positive bacteria, the Fsr system (Enterococcus faecalis
system regulator) is one example of QS that controls the expression of two virulence factors, gelatinase and serine protease, important for the prevalence and survival of
E. faecalis during infection. Enterococcus is a peculiar and controversial genus of
Gram-positive lactic acid bacteria. It includes commensal species inhabiting the gastro-intestinal tracts of humans and animals. However, they are also capable of causing opportunistic infections including bacteraemia, endocarditis, meningitis, wound, urinary tract and nosocomial bloodstream infections. E. faecalis is the predominant species in human/animal associated environments, and therefore the most studied species of this genus. Recent data indicates that E. faecalis is the third most commonly isolated nosocomial pathogen (12% of all hospital infections). Over representation of E. faecalis among clinical isolates may be related to its natural abundance, to the presence of virulence factors and/or the ability to acquire easily antibiotic resistances. The two most studied virulence factors are gelE (encoding gelatinase, GelE) and sprE (encoding a serine protease, SprE). These genes are present in clinical and diary enterorococal strains but are not always phenotypically positive. Fsr system and GeE-SprE have been proven important for E. faecalis
sprE genes but a negative gelatinase phenotype (Chapter II). The fsr and gelE-sprE
operons were sequenced, and the negative gelatinase phenotype was attributed to a nonsense mutation (a premature STOP codon). This mutation in the fsrC gene is translated into a deficient ATPase sensor domain, responsible for sensing and transducing the signal from the quorum-sensing molecule. This mutation was found in other enterococcal strains revealing that this is a natural way to shutdown the QS-associated production of GelE and SprE and suggesting that some benefits may come from silencing the QS.
In a previous microarray study from our lab in strain E. faecalis V583, fsr-gelE-sprE
genes were found to be down-regulated by vancomycin, a cell-wall active antibiotic,
(Chapter III). In order to check the hypothesis of QS shutdown by vancomycin, we used E. faecalis V583ΔfsrB, which is unable to produce GBAP but is able to sense it. Cells from this mutant were collected after incubation (0 min, 10 and 20 min) with GBAP or with GBAP and vancomycin .Expression levels of gelE, sprE and vanB
genes were evaluated by semi-quantitative RT-PCR. When GBAP was added after vancomycin none of the Fsr regulated genes, gelE and sprE, was induced, suggesting that this antibiotic turns FsrC sensor blind to the QS molecule. It was the first time Fsr system activity was associated with an antibiotic. These two previous studies indicate that Fsr associated QS is not essential for growth as it is repressed in some environments.
xvii resulted in up-regulation of two genes important in cell autolysis – lrgAB – that were confirmed to be regulated by LytRS.
Drosophila melanogaster has proven useful as a tool to study host-pathogen
interactions, both for bacterial pathogens and human commensals. We therefore chose this model organism to gather clues on the role of Fsr, and of the genes it regulates, on host disease and death inflicted by E. faecalis infections. To study the mechanism of pathogenesis associated with Fsr, proteases and new genes,
Drosophila was first established as an infection model. We then infected the fruit fly with V583 mutants on the newly found genes. Two new Fsr - associated virulence factors were found, namely, lrgAB operon and the bacteriocin coding gene ef1097. We also found that inactivation of Fsr and the two proteases increased fly´s tolerance to E. faecalis, whereas flies showed similar resistance towards all studied E. faecalis
strains. These results suggest that future approaches to combat the E. faecalis
infection can be through improving host tolerance, providing an alternative, or a complement, to the use of antibiotics.
These new findings lead us to further investigate the role of Fsr in the cross-talk with
the Drosophila immune system (Chapter V). We measured the expression of
drosomycin; analyzed the phagocytosis and the melanization during Drosophila
infection (control W1118, W1118HmlΔ>GFP/UAS-Bax and W1118 PPO1Δ, PPO2Δ) with
V583 and V583ΔfsrBΔgelEΔsprE strains. We found that Fsr interfered with the melanization process. Moreover, Drosophila was only able to survive in the absence of both Fsr-GelE-SprE factors and melanization. Melanization is used by Drosophila
to combat the pathogens but when exacerbated is also able to cause host injury. We believe that fly death, caused by E. faecalis carrying Frs-GelE-SprE, is due to exacerbated host injury by host´s own immune response.
xix Desde a descoberta do Quorum Sensing (QS), percebeu-se que as bactérias têm uma espécie de “vida social” e que cooperam e coordenam as suas actividades no hospedeiro/ambiente que infectam/habitam. Muitas bactérias só se tornam perigosas quando sentem que estão em número suficiente para sobrecarregar a defesa humana. Em seguida, libertam toxinas e causam doença e, eventualmente, morte. A partir do momento em que foi feita esta importante descoberta, muitas bactérias (Gram negativas e Gram positivas) foram identificadas como participantes no QS. Em bactérias Gram positivas, o sistema Fsr (Enterococcus faecalis system regulator) é um exemplo de QS que tem como molécula sinal a GBAP (Gelatinase biosynthesis
activating pheromone).O Fsr controla a expressão de dois factores de virulência,
gelatinase e proteinase sérica, importantes para a persistência e sobrevivência de E.
faecalis durante a infecção. Enterococcus é um género peculiar e controverso que
pertence ao grupo de bactérias lácticas Gram-positivas. Este género inclui espécies comensais que habitam no trato gastrointestinal de humanos e animais. Contudo, são capazes de causar infecções oportunistas como bacteremias, endocardites, meningites, feridas, infecções urinárias e infecções nosocomiais da corrente sanguínea. E. faecalis é a espécie predominante em humanos/animais, e também a mais estudada. Estudos recentes indicam que E. faecalis é o terceiro patogénico nosocomial mais comum (12% das infecções hospitalares). A abundante presença
de E. faecalis em isolados clínicos poderá estar relacionada com a sua natureza de
produzirem factores de virulência e/ou com a sua facilidade de adquirirem resistência a antibióticos. Os factores de virulência mais estudados são gelE
hospedeiro e o patogénico. O trabalho desta tese foi desenhado para preencher estas lacunas do conhecimento.
Em certas condições, E. faecalis desliga o QS Fsr. É o caso da estirpe E. faecalis
LN68 que tem todos os genes fsr e gelE-sprE mas apresenta um fenótipo gelatinase negativo (Capítulo II). Os operões fsr e gelE-sprE foram sequenciados e o fenótipo gelatinase negativo foi atribuído à presença de uma mutação sem sentido (um codão STOP prematuro). Esta mutação no gene fsrC afecta o domínio sensor ATPase, responsável por sentir e traduzir a GBAP. Esta mutação foi também identificada noutras estirpes de Enterococcus revelando ser uma forma natural de desligar a produção de GelE e SprE associada ao QS, e sugerindo que esta bactéria poderá ter benefícios energéticos com o silenciamento do QS.
Num estudo anterior de transcriptómica em E. faecalis V583, os genes fsr-gelE-sprE
foram identificados como serem negativamente regulados pela vancomicina, antibiótico que inibe a síntese da parede celular, (Capítulo III). Para testar a hipótese de que o QS é desligado pela presença da vancomicina, usou-se a estirpe mutante E. faecalis V583ΔfsrB, que é incapaz de produzir GBAP mas sente o seu sinal e activa a síntese das proteases. Células deste mutante foram recolhidas após incubação (0 minutos, 10 e 20 minutos) com GBAP ou com GBAP e vancomicina. Os níveis de expressão dos genes gelE, sprE e vanB foram quantificados por RT-PCR semi-quantitativo. Quando a GBAP foi adicionada depois da vancomicina nenhum dos genes regulados pelo Fsr, nomeadamente gelE e sprE, foram induzidos sugerindo que este antibiótico torna o sensor FsrC cego à molécula QS. Esta constitui a primeira vez em que a actividade do sistema Fsr foi correlacionada com um antibiótico. Estes dois estudos previamente descritos demonstraram que o QS associado ao Fsr não é essencial ao crescimento da bactéria mas é reprimido em alguns ambientes/condições.
Estudos anteriormente realizados têm sugerido que o Fsr é um regulador global em
xxi transcriptómica usando mutantes isogénicos de V583 no Fsr e nas proteases, para identificar os genes directamente e indirectamente regulados por este sistema de
QS (Capítulo IV). O QS foi artificialmente induzido pela GBAP, adicionada
exogenamente e de uma forma controlada. O sistema Fsr foi identificado como regulador de cinco genes (gelE, sprE, ef1097 e ef1351-52). Doze genes adicionais foram identificados como sendo dependentes da presença das proteases induzidas pelo QS. Adicionalmente, a indução do gelE e sprE resultou na regulação positiva de dois genes importantes para a autólise celular – lrgAB- tendo-se confirmado que são regulados pelo sistema de dois componentes LytRS.
Drosophila melanogaster tem vindo a demostrar ser uma ferramenta útil para o
estudo da interacção do hospedeiro – patogénico, em particular, no estudo de bactérias patogénicas e comensais humanos. Foi por essa razão que se escolheu como modelo para estudar a função do Fsr e dos genes que este regula, no desenvolvimento da doença do hospedeiro e na morte inerente à infecção causada
por E. faecalis. D. melanogaster foi primeiro estabelecida como modelo de infecção
para estudar o mecanismo patogénico associado ao Fsr, proteases e os novos genes. Em seguida, a mosca da fruta foi infectada com mutantes de V583 nos genes novos anteriormente identificados. Dois novos factores de virulência associados ao Fsr foram assim identificados, nomeadamente lrgAB e a bacteriocina ef1097. Também foi observado que a inactivação do Fsr e das duas proteases aumenta a tolerância da mosca relativamente a E. faecalis, sido demonstrada resistência similar para todas as estirpes E. faecalis testadas. Este resultado sugere que o combate de infecções por E. faecalis poderá passar por beneficiar a tolerância do hospedeiro, proporcionando uma alternativa ou um complemento ao uso de antibióticos.
Estas novas descobertas levaram-nos a investigar a função do Fsr na comunicação com o sistema imunitário da Drosophila (Capítulo V). Durante a infecção de
Drosophila (controlo W1118, W1118HmlΔ>GFP/UAS-Bax and W1118 PPO1Δ, PPO2Δ)
sobreviver quando, em simultâneo, não estão presentes nem o Fsr e as proteases GelE-SprE, nem a melanização. Esta é um mecanismo utilizado pela Drosophila
para combater os patogénicos mas quando exacerbada é capaz de provocar danos no próprio hospedeiro. É possível que a morte da mosca da fruta causada por E.
faecalis com Fsr-GelE-SprE seja devida às lesões decorrentes de uma exacerbada
resposta imune do hospedeiro.
xxiii
Abbreviation Full form
Δ Deletion
ΔCt Cycling threshold Ace Adhesion to collagen
agr Accessory gene regulator AIP Autoinducer peptide
AHL Acyl-homoserine lactone
AIs Autoinducers
AMPs Antimicrobial peptides
A. thaliana Arabidopsis thaliana
AS protein Aggregation substance
Bee Biofilm enhancer
BHI Brain heart infusion
bp Base pairs
°C degree Celsius
C. elegans Caenorhabditis elegans
cDNA Complementary DNA
CFUs Colony forming units
cps cluster Capsular polysaccharides
Cyl Cytolysin
ddl D-alanine-D-alanine ligase D-Lac D-lactate
D-Ser D-serine
DNA Deoxyribonucleic acid
Drosophila Drosophila melanogaster
Ebp Endocarditis and biofilm associated pili
(e)DNA Extracellular DNA
ElrA Surface protein
epa cluster Enterococcal polysaccharide antigen EPS Extracellular polymeric substances
E. gallinarum Enterococcus gallinarum E. faecalis Enterococcus faecalis E. faecium Enterococcus faecium
Esp Surface protein
fsr E. faecalis regulator iRNA RNA interference
GBAP Gelatinase biosynthesis activating pheromone
GelE Gelatinase
G. melonella Galleria melonella
GRAS Generally Recognized As Safe
h hours
HK Histidine Kinase
hld virulence factor δ-lysin
JAK/STAT Janus kinase/signal transducer and activator of transcription
LB Luria Bertani Broth
LN68 Strain isolated from Niza milk
LSE4 Strain isolated from Serra da Estrela milk
MDR Multi-Drug Resistance
mg milligram
min minute
ml millilitre
M. luteus Micrococcus luteus
mRNA messenger RNA
M. sexta Manduca sexta
NaCl Sodium Chloride
NCBI National Center for Biotecnology Information
nl nanoliters
ng nanogram
nM nanoMolar
nt Nucleotides
mM milliMolar
OD Optical density
OG1RF E. faecalis OG1RF
xxv
PRR Pattern recognition receptor
PG Peptidoglycan
(PGRP)-LC Peptidoglycan recognition protein
QA29b Strain isolated from Azeitão cheese
qRT-PCR Quantitative real-time polymerase chain reaction
QS Quorum sensing
S. aureus Staphylococcus aureus
SPHs Serine protease homologues
SprE Serine protease
StrA Sortase
TM Melting temperature
VRE Vancomycin resistant Enterococci
V583 E. faecalis V583 RHK Receptor histidine kinase
RNA Ribonucleic Acid
rDNA Ribossomal RNA
ROS Reactive oxygen species
RR Response Regulator
RT-PCR Reverse Transcriptase-polimerase chain reaction
V. fischeri Vibrio fischeri V. harveyi Vibrio harveyi
wt Wild-type
xxvii
Acknowledgments ... vii
Thesis outline ... xi
Thesis publications ...xiii
Abstract ... xv
Resumo ... xix
Abreviations ... xxiii
Table of contents ... xxvii
INTRODUCTION
1. QUORUM SENSING
A Way to Communicate………..……….5 1.1 Different QS Systems Among Bacteria……….6
1.2 Enterococcus faecalis Fsr Quorum Sensing System………..8
2. ENTEROCOCCUS GENUS
General Characteristics……….………12
2.1 Enterococcus spp. - An Opportunistic Pathogen………...13
3. ENTEROCOCAL VIRULENCE
The Role of Fsr, Gelatinase and Serine Protease………...16 3.1 Animal Models to Study Fsr and Proteases………...20
4. DROSOPHILA MELANOGASTER
A model to Study Host-pathogen Interaction………..………23
5. AIMs AND SCOPE OF THESE THESIS………...………...30
6. Bibliography……..…...………..………32
1. Summary………...……….51 2. Introduction………52 3. Material and methods………...55 4. Results and Discussion………59 5. Acknowledgements………..67 6. Bibliography……...………68 7. Supplementary data……….74
FSR AND VANCOMYCIN:
The antagonistic relation
1. Summary………81 2. Introduction………82 3. Material and methods………...84 4. Results and Discussion………...86 5. Bibliography………...90
NEW FINDINGS ON FSR SYSTEM:
New
virulence genes and their impact during Drosophila infection
1.Summary……….99 2. introduction………..100 3. material and methods……….103 4. Results………..110 5. Discussion………118 6. Acknowledgements………124 7.Bibliography……….125 8.Supplementary data………135
CHAPTER III
xxix
The collapse of the immune system
1.Summary………...145 2. Introduction………...…...146 3. Material and methods……….149 4. Results ……….152 5. Discussion………159 6. Acknowledgements………162 7.Bibliography……….………163
GENERAL DISCUSSION
1.FSR QUORUM SENSING SYSTEM
- Different environments lead different ways to persist……….……173
2.FSR REGULON
- New genes and potential virulence factors………...177
3.FUTURE PRESPECTIVES
- Fsr system can be a future target for therapy………..183
4. Bibliography………...………..185
CONTENTS
1. QUORUM SENSING
A Way to Communicate ... 5
1.1 Different QS Systems Among Bacteria ... 6
1.2 Enterococcus faecalis Fsr Quorum Sensing System ... 8
2. ENTEROCOCCUS GENUS
General Characteristics ... 12
2.1 Enterococcus spp. - An Opportunistic Pathogen ... 13
3. ENTEROCOCAL VIRULENCE
The Role of Fsr, Gelatinase and Serine Protease ... 16
3.1 Animal Models to Study Fsr and Proteases ... 20
4. DROSOPHILA MELANOGASTER ... 23
A model to Study Host-pathogen Interaction
5. AIMs AND SCOPE OF THESE THESIS ... 30
5
Chapter I
1.QUORUMSENSING
A Way to Communicate
Many bacteria are known to regulate their cooperative activities and physiological processes trough a mechanism called quorum sensing (QS), in which bacterial cells communicate with each other by releasing, sensing and responding to small diffusible signal molecules. QS as a concept grew out of the work of JW Hastings in the 1960’s (Hastings & Greenberg, 1999). QS describes a process of cell-to-cell communication used by bacteria to understand and adapt in their environment and consequently to apply specific strategies that allow adaptation to environmental stress in space and time (Fuqua et al., 1994; Skandamis & Nychas, 2012).
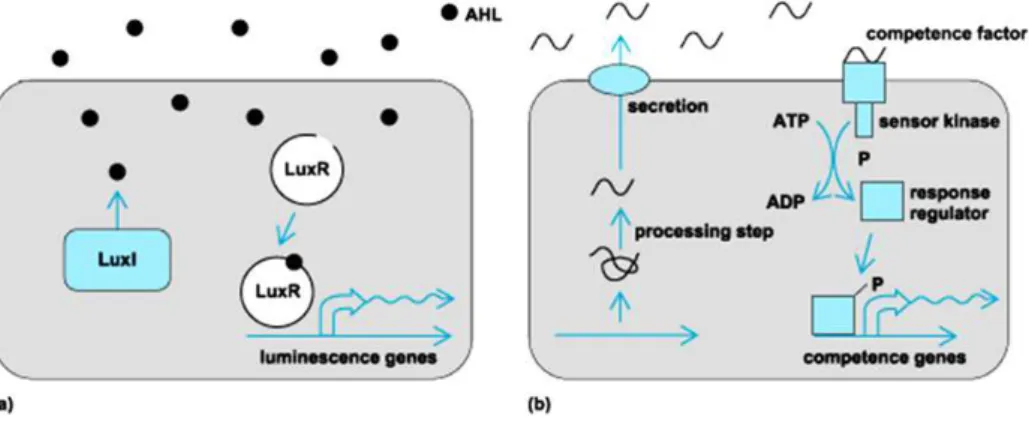
QS cell-to-cell signaling results from production of small, diffusible signal molecules called autoinducers that are secreted at a basal level during bacterial growth by emitter cells. This signal accumulates in the surrounding environment. When the concentration of these signal molecules reaches a threshold level (the quorum level), the signal molecule binds to receptors on or in the bacteria cell and alter gene expression (Figure 1) (Skandamis & Nychas, 2012).
A B
QS control genes that are beneficial when expressed by groups of bacteria acting in synchrony. The list of processes that bacteria coordinate by QS is extensive. These activities are generally unproductive when carried out by small number of cells. Bacteria use quorum sensing communication circuits to regulate a diverse array of physiological activities that include symbiosis, virulence, competence, conjugation, antibiotic production, motility, sporulation, and biofilm formation (Rutherford & Bassler, 2012).
One classical example of QS is the production of bioluminescence by the marine bacteria Vibrio fischeri, a symbiont of Hawaiian bobtail squid Euprymna scolopes
(Nealson & Hastings, 1979; Reading & Sperandio, 2006). V. fischeri lives in squid light organ (nutrient-rich environment) and is only beneficial to the bacteria to synthesize the light producing enzymes (luciferase) when certain number of autoinducers inside the squid light organ are detect. In contrast, autoinducers do not accumulate to any significant level in free ocean and V. fischeri does not make light. On the other hand, light production by the bacteria enables the squid to eliminate its shadow in the ocean and thus the light is used as squid in a strategy to evade predators (Bassler, 1999; Reading & Sperandio, 2006).
1.1 Different QS Systems Among Bacteria
QS systems in bacteria have been generally divided into at least three classes: (1)
LuxI/LuxR-type QS in Gram negative bacteria which use acyl-homoserine lactones (AHL) as signal molecules; (2) oligopeptides-two-component-type QS in Gram positive bacteria, which use small, often post-translationally modified peptides as a signal molecule; and (3) luxS-encoded autoinducer 2 (AI-2) QS in both Gram negative and Gram positive bacteria (Li & Tian, 2012).
7
Chapter I
high concentration, the autoinducer binds to a transcription factor of the LuxR type in the cytoplasm, and this LuxR-AHL complex binds to a specific promoter thereby activating gene expression for group-specific processes, like bioluminescence (lux) (Figure 2)(Bassler & Vogel, 2013; Bassler, 1999).
In Gram positive bacteria there are two types of QS systems. The first type uses autoinducer peptides (AIP), called peptide pheromones. Gram positive bacteria normally produce a signal peptide precursor, which is cleaved and the active AIP is then exported through a peptide-specific ABC transporter into their environment. AIP is recognized by the input domain of a typical sensor element of a histidine kinase (HK) two-component signal transduction system. HK phosphorylates the response regulator (RR) which binds to the promoter of genes of interest (Figure 2).
Figure 2 - Quorum-sensing signalling pathways in Gram negative (A) and Gram positive bacteria
(B). (A) LuxI/LuxR in a Gram-negative bacteria. The autoinducer (black dots) synthesized through pathways involving LuxI, is released, and then reenters into bacteria and binds to receptors (LuxR) that alter cellular response elements. (B)QS in Gram positive bacteria. Amino acids or short peptides (wavy lines) are exported and then bind to cell surface-bound sites that activate phosphorylation cascades, leading to transcriptional changes (Raffa et al., 2005) (http://www.accessscience.com).
sensing system, as well as those involved in the secretion of peptide, are transcriptionally linked, and the synthesis of the peptide signal forms an autoregulatory loop (Figure 2) (Waters & Bassler, 2005).
In recent years, a second type of QS system has been identified in several groups of Gram-positive streptococci. This system is called ComRS, and uses a small double-tryptophan peptide pheromone (XIP) as a signal molecule. XIP is internalized by an oligopeptide ABC transporter typified by Opp/Ami, and interacts with transcriptional regulator ComR, a proximal regulator of sigX that encodes a master regulator or alternative sigma factor SigX (ComX). Later , the competence genes for genetic transformation are activated (Li & Tian, 2012).
In additional to these QS systems, another QS type called autoinducer 2 (AI-2) has been described in both Gram negative and Gram positive organisms. This type of QS system enables inter-species signaling. AI-2 was first characterized in marine bacterium Vibrio harveyi and contributes to regulation of cell-density-dependent bioluminescence. The synthesis of AI-2 depends on a luxS encoded synthase, which is a metabolic enzyme involved primarily in the conversion of ribosyl-homocysteine into homocysteine and 4,5-dihydroxy-2,3-pentanedione (DPD), the precursor of AI-2. The LuxR protein is a cytoplasmic receptor and transcription activator. Homologues
of luxS have been found in many species of bacteria, suggesting that AI-2 QS is
widely spread among prokaryotes (Li & Tian, 2012).
1.2 Enterococcus faecalis Fsr Quorum Sensing System
One example of QS with a cell density-dependent two-component regulatory system mechanism in Gram positive bacteria is the Fsr system in Enterococcus faecalis (E.
faecalis regulator). This QS system, first described by Qin et al, controls the
9
Chapter I
2001b; Nakayama et al., 2006; Qin et al., 2000; Qin et al., 2001). The fsr locus consists of four genes fsrA, fsrB, fsrD and fsrC, which collectively regulate the expression of gelE and sprE. The fsr locus has significant similarity to the accessory gene regulator (agr) locus of staphylococci that includes within it the virulence factor
δ-lysin (hld) and regulates expression of a number of additional exotoxins (Figure 3). In the Fsr system, a cyclic peptide, gelatinase biosynthesis-activating pheromone (GBAP), acts as the autoinducer.
Figure 3 –Fsr Quorum Sensing.A schematic diagram with proposed mechanism of fsr activation and its effect on gelatinase and serine protease synthesis (www.agr.kyushu-u.ac.jp/lab/microbt/Research/QuorumSensing.html).
transcripts (Figure 3) (Nakayama et al., 2001a; Nakayama et al., 2001b; Nakayama
et al., 2006; Qin et al., 2000; Qin et al., 2001).
Qin et al (2000) reported that the gelatinase phenotype requires the concomitant
presence of the fsr and gelE (Qin et al., 2000). In the years following this finding, several studies reported the loss of the gelatinase phenotype in different
Enterococcus strains. In some cases, this phenomenon was found to be associated
with a deletion of part of Fsr operon, but in other cases incongruence between the genotype and the phenotype was reported. All these reports indicate that the loss of GelE phenotype, in the presence of an apparently complete fsr operon, occurs both in natural and laboratory subcultured E. faecalis isolates (Eaton & Gasson, 2001; Galloway-Pena et al., 2011; Lopes Mde et al., 2006; Nakayama et al., 2002).
Figure 4 – Structure and properties of GBAP (Gelatinase Biosynthesis Activating Pheromone).
GBAP is a cyclic peptide pheromone in E. faecalis with 11 aminoacid residues and a cyclic peptide containing a lactone linkage. The lactone ring is indispensable for the activity (Nakayama et al., 2001b).
In 2006, Bourgogne et al. made a transcriptomic study comparing OG1-RF (a clinical E. faecalis isolate) with its isogenic fsrB deletion mutant and suggested that
11
Chapter I
regulator of biofilm formation in plastic surface. The detailed mechanism of gene regulation and pathogenesis associated with Fsr system and proteases are currently incomplete (Bourgogne et al., 2006).
2.ENTEROCOCCUSGENUS
General Characteristics
The name “entérocoque” was first used in 1899 by Thiercelin to identify a new species of Gram positive coccus found in the human gut (Thiercelin, 1899). Enterococci are lactic acid Gram positive bacteria, with ovoid shape (coccus) that grows in short chains, pairs or as single cells. They are facultative anaerobic bacteria, catalase negative and can grow between 10-45ºC, although their optimal temperature is 35-37ºC. Most enterococcal species are able to grow in the presence of 6, 5% of NaCl, at pH 9, 6 and 40% of bile salts. They are homo-fermentative; and produce lactic acid from glucose (Mundt, 1986).
The identification of species from the genus Enterococcus by physiological tests has always been problematic because of their considerable phenotypic diversity. Furthermore, identification of species by conventional tests often requires long incubation times. Genotypic identification methods using the 16S and 23S rDNA genes are more accurate; although they cannot differentiate between all
Enterococcus species (e.g. Enterococcus gallinarum and Enterococcus casseliflavus
show 99.8% homology in their 16S rDNA). Alternative methods have been successfully applied using amplification of specific genes, for example, the ddl gene with encodes D-alanine-D-alanine ligase (Ogier & Serror, 2008). Nowadays, the genus Enterococcus is composed of forty-five species, with the most common species studied being Enterocccus faecalis and Enterococcus faecium
(http://old.dsmz.de). Historically E. faecalis has been the most intensively studied due to its prominence in the nosocomial setting.
13
Chapter I
(Bourgogne et al., 2008) was reported and in 2010 the genome of 28 other enterococcal strains (including E. faecalis, E. faecium, E. casseliflavus and E. gallinarum species) became available (Palmer et al., 2010).
E. faecalis and E. faecium are natural members of the gastrointestinal microbiota in humans, varying in abundance among individuals along the gastro-intestinal tract. Enterococci are commonly isolated from foods, plants, water and soils, because of their use in fermentations, and also as a result of dissemination from fecal sources combined with their natural tolerance to adverse environmental conditions (Lopes et al., 1999; Ogier & Serror, 2008). Unlike many other lactic acid bacteria, enterococci are not considered as “Generally Recognized As Safe” (GRAS) and their detection in water is regarded as an indicator of fecal contamination (Godfree et al., 1997). Enterococci therefore have an ambiguous status concerning assessment of enterococci food containing safety. On the one hand, enterococci are used in cheese making, in the development of flavors, aroma and contributing to the ripening of cheeses such Cheddar, Feta and Mozarella. On the other hand, their ability to produce biogenic amines in cheese and fermented sausages and their propensity for genetic exchange constitute negative aspects for their utilization as probiotic (Foulquie Moreno et al., 2006; Giraffa, 2003; Ogier & Serror, 2008).
2.1 Enterococcus spp. - An Opportunistic Pathogen
indicate that E. faecalis is the third most commonly isolated nosocomial pathogen (12% of all hospital infections) (Hollenbeck & Rice, 2012).
Currently, in the United States and Europe, infections caused by E. faecium are much more frequently resistant to vancomycin and ampicillin than those caused
by E. faecalis. E. faecium is now almost as common a cause of nosocomial infection
as E. faecalis. This change in species is of paramount clinical importance, as E.
15
Chapter I
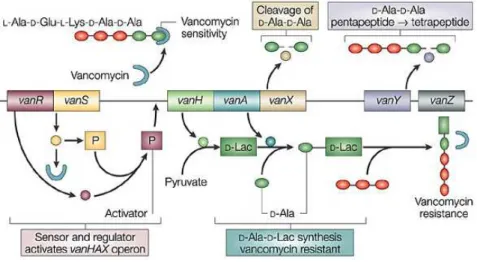
of VRE was reported in the United States (Murray, 1990). The mechanism of vancomycin resistance has been extensively studied. Vancomycin binds with high affinity to the D-Ala-D-Ala C-terminus of peptidoglycan, blocking the addition of late precursors by transglycosylation to nascent peptidoglycan chain and preventing subsequent cross-linking by transpeptidation and stopping the peptidoglycan production (Arthur & Courvalin, 1993; Hughes, 2003). Glycopeptide resistance involves two pathways: replacement of the terminal D-Ala of peptidoglycan precursor with D-lactate (D-lac), which produces high-level resistance, or with D-Ser, which produces low level resistance (Arias & Murray, 2012). In the most common mechanism of vancomycin resistance, the two component system VanR-VanS regulates the expression of the vancomycin resistance operon (Figure 5).
3.ENTEROCOCALVIRULENCE
The Role of Fsr, Gelatinase and Serine Protease
For enterococci to act as pathogen they must first adhere to host tissues. During infection of sterile tissues, enterococci encounter an environment vastly different than the gut, with high redox potentials, limited nutrients, phagocytic leukocytes, and other host defenses. Enterococci express factors – virulence factors - that permit adherence to host cell and extracellular matrix, facilitate tissue invasion, effect immunomodulation and cause toxin-mediated damage (Gilmore, 2002).
The first examination of enterococal virulence was reported in 1899, the same year this organism was discovered. MacCalum and Hasting described a fatal case of endocarditis caused by an organism that they termed Micrococcus zymogenes. The bacteria expressed cytolitic (or hemolytic) and protease (gelatinase) activities and likely represented E. feacalis (MacCallum & Hastings, 1899). Since then
Enterococcus virulence has been extensively studied. About a dozen putative
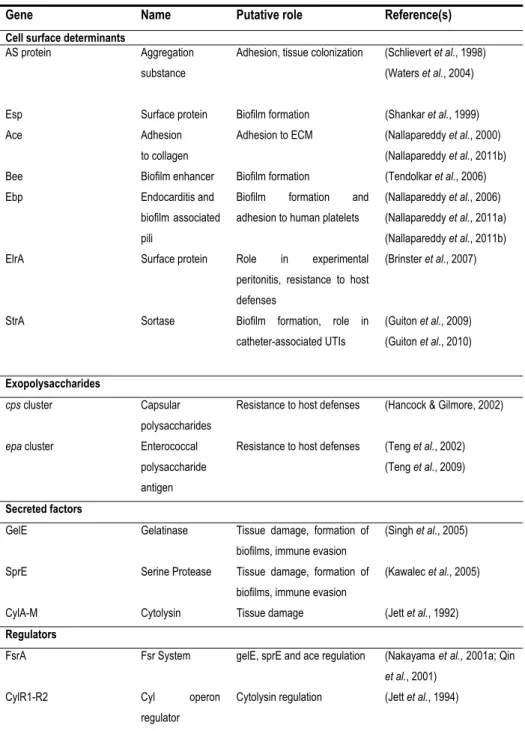
virulence factors have been reported from virulence analysis in various animal models (Table 1). They are involved in attachment both to host cells and to extracellular matrix proteins (AS, Esp, EfaA), in resistance to macrophages (AS, HypR), in cell and tissue damage (Cyl, GelE, SprE) and in immune system evasion (capsular polysaccharides) (Gilmore, 2002; Tendolkar et al., 2003).
17
Chapter I
Table 1 – E. faecalis virulence factors and their putative role (Arias & Murray, 2012; Jett et al., 1994; Ogier & Serror, 2008).
Gene Name Putative role Reference(s)
Cell surface determinants
AS protein Aggregation substance
Adhesion, tissue colonization (Schlievert et al., 1998) (Waters et al., 2004)
Esp Surface protein Biofilm formation (Shankar et al., 1999) Ace Adhesion
to collagen
Adhesion to ECM (Nallapareddy et al., 2000) (Nallapareddy et al., 2011b) Bee Biofilm enhancer Biofilm formation (Tendolkar et al., 2006) Ebp Endocarditis and
biofilm associated pili
Biofilm formation and adhesion to human platelets
(Nallapareddy et al., 2006) (Nallapareddy et al., 2011a) (Nallapareddy et al., 2011b) ElrA Surface protein Role in experimental
peritonitis, resistance to host defenses
(Brinster et al., 2007)
StrA Sortase Biofilm formation, role in catheter-associated UTIs
(Guiton et al., 2009) (Guiton et al., 2010)
Exopolysaccharides
cps cluster Capsular polysaccharides
Resistance to host defenses (Hancock & Gilmore, 2002)
epa cluster Enterococcal polysaccharide antigen
Resistance to host defenses (Teng et al., 2002) (Teng et al., 2009)
Secreted factors
GelE Gelatinase Tissue damage, formation of biofilms, immune evasion
(Singh et al., 2005)
SprE Serine Protease Tissue damage, formation of biofilms, immune evasion
(Kawalec et al., 2005)
CylA-M Cytolysin Tissue damage (Jett et al., 1992)
Regulators
FsrA Fsr System gelE, sprE and ace regulation (Nakayama et al., 2001a; Qin
et al., 2001) CylR1-R2 Cyl operon
regulator
Cytolysin regulation (Jett et al., 1994)
E. faecalis is an example of an opportunistic pathogen that uses QS system to produce virulence factors to succeed during infection. One of the most studied is the
fsr QS (see below) that regulates the virulence factors serine protease and gelatinase. The serine protease has high similarity to the Staphylococcus glutamil-endopeptidases but has not been purified or characterized. Some studies have reported this protease to have some role in biofilm formation, but its exact role is still unknown (Kawalec et al., 2005). The gelatinase has been largely described as an important virulence factor. This protease was first described in 1975 by Gold et al. who identified a protease in E. faecalis OG1-10 responsible for human gelatin oral degradation, suggesting that it was a virulence factor (Gold et al., 1975). This protein is secreted as a zinc metalloprotease (thermolysin-M4 protease) capable to hydrolyze numerous subtracts, including gelatin, collagen, casein, fibrin, hemoglobin and other small bioactive peptides. The protein gelatinase produced by E. faecalis
OG1-10 was isolated by Makinen et al. in 1989 (Makinen et al., 1989). The gene was subsequently identified (gelE) and its sequence determined (Su et al., 1991). Gelatinase activity was detected in enterococal clinical strains (Lopes Mde et al., 2006; Singh et al., 2005).
19
Chapter I
turnover, cell separation, recycling of muropeptides, lysis by cell-wall synthesis inhibitors, competence, sporulation, flagellum formation and pathogenicity (Bayles, 2007; Mohamed & Huang, 2007). Autolysis control may also be involved in tolerance to cell-wall active antibiotics, as previously demonstrated for the homologous Agr system of Staphylococcus aureus (Antignac et al., 2007; Bayles, 2007). Knowing that PG degradation products are also a major elicitor of the host immune response it is obvious to assume/speculate a relation between autolysis and host immune recognition and response. Therefore, autolysis control is crucial for virulence, stress response and host immune system modulation (Antignac et al., 2007; Bayles, 2007; Thomas et al., 2008).
Different studies described that gelatinase has a critical role in biofilm formation (Hancock & Perego, 2004; Kristich et al., 2004; Mohamed & Murray, 2005; Mohamed & Murray, 2006). In 2008, Thomas et al invoked the fratricidal model for E. faecalis
biofilm development. In this model, GelE activated the lysis of a subpopulation of bacteria and thereby catalyzes the release of genomic DNA (e)DNA, as originally proposed for S. pneumoniae autolysis (Gilmore & Haas, 2005). SprE negatively affects autolysis, (e)DNA release and early biofilm maturation by negatively regulating GelE activity, and thereby acts as an immunity protein against lysis. GelE and SprE execute their characteristic effects following downstream interactions with the primary autolysin, AtlA, by modifying the cell-wall affinity of proteolytically processed AtlA. The interplay of the two secreted and co-regulated proteases seems to be tightly regulated. A minority subpopulation of quorum non-responders (GelE– SprE–) act as prey cells, for targeted fratricide mediated by the quorum-responsive predator cells that form the majority in the biofilm. In response to the GBAP peptide, predator cells secrete GelE and SprE proteases. Prey cells are susceptible cells that have not (yet) responded to GBAP. If GelE reaches the cells before SprE, this results in lysis via release of AtlA from their surface, and this in turn can also lyse neighboring cells (Thomas et al., 2008; Thomas et al., 2009).
ability of cells to adhere to collagen. The bacterial surface protein has an important role in E. faecalis virulence by mediating adherence and colonization to host tissue which is an early step toward clinical infection (Pinkston et al., 2011). Another study indicated that, the absence of gelatinase (in E. faecalis OG1RF) leads to high levels of secreted antigenic SalB, in the exoproteome. The relation between GelE and SalB it still not clear but it is known that the absence of SalB increases autolysis and cell morphological changes (Shankar et al., 2012). In addition to these studies, GelE and SprE have also been reported to have an important role in translocation across intestinal T84 cells and in degradation of antimicrobial peptides (AMPs) from immune system of Galleria mellonella (Cytrynska et al., 2007).
All these studies indicate that Fsr-GelE-SprE has an important role in E. faecalis
virulence and place these traits as promising targets to combat the E. faecalis
infection. Nakayama et al., discovered two secondary metabolites, siamycin and ambuic acid, which act as QS inhibitors. Siamycin inhibits GBAP-induced phosphorylation of receptor histidine kinase FsrC and ambuic acid inhibits the proteolytic processing of FsrD, the propeptide of GBAP. However, none of these compounds influence E. faecalis growth (Nakayama et al., 2007; Nakayama et al., 2009).
3.1 Animal Models to Study Fsr and Proteases
21
Chapter I
Table 2 – Host models used to study virulence associated to the Fsr and/or gelatinase.
Animal model used E. faecalis mutants used for
the study
Enterococcus strains References
Rabbit endophthalmitis OG1RFΔfsrB2 E. faecalis OG1RF (Mylonakis et al., 2002)
Rat endocarditis
OG1RFΔfsrB1
OG1RFΔfsrB2
OG1RFΔgelE1
E. faecalis OG1RF (Singh et al., 2005)
Galleria mellonella
QA29bΔfsrB2
QA29bΔgelE2
LSE4aΔfsrB2
LSE4aΔgelE2
E. faecalis OG1RF
E. faecalis QA29B
E. faecalis LSE4a
E. faecalis LN68
E. faecium QSE32
E. durans QN1
E. faecalis ATCC 51299
(Gaspar et al., 2009; Park et al., 2007)
Arabidopsis thaliana
OG1RFΔfsrA, OG1RFΔfsrB1,
OG1RFΔfsrC1,
OG1RFΔgelE1,
OG1RFΔsprE1
E. faecalis FA-2-2,
E. faecalis V583,
E. faecalis OG1RF
(Jha et al., 2005)
Caenorhabditis elegans OG1RFΔfsrB1
OG1RFΔgelE1
E. faecalis OG1RF (Sifri et al., 2002)
Zebrafish OG1RFΔfsrB1
OG1RFΔgelE1
OG1RFΔsprE1
OG1RFΔgelEΔsprE1
E. faecalis OG1RF (Prajsnar et al., 2013)
Different outcomes have been observed in different assays when fsrABC or gelE-sprE mutants were compared with the parental strain. In rat endocarditis a greater decrease in endocarditis severity was observed when the proteases were deleted comparing with deletion of fsrB (Singh et al., 2005). In other studies the observation was the opposite - in rabbit endophtalmitis (Engelbert et al., 2004; Mylonakis et al., 2002), murine and C. elegans infection (Garsin et al., 2001; Sifri et al., 2002), G. mellonella infection (Gaspar et al., 2009) - a greater attenuation was observed when
23
Chapter I
4.DROSOPHILAMELANOGASTER
A model to Study Host-pathogen Interaction
The use of invertebrate animal models has provided tremendous insights into pathogen-host interaction of many human pathogens, and has revealed that many aspects of these interactions in higher host organisms are conserved in invertebrates. The fruit fly Drosophila melanogaster (Drosophila) is one of the most used for studying host-pathogen interaction of bacteria, fungal and viral pathogens. The life cycle of Drosophila has different stages: the embryo, 1st instar larva, 2nd instar larva, 3rd instar larva, prepupa, pupa and adult (Igboin et al., 2012) (Figure 6).
Figure 6 – The life cycle of Drosophila. The life cycle is divided in six stages: embryo, 1st instar larva, 2nd
instar larva, 3rd instar larva, prepupa, pupa and adult.
Drosophila is a model organism with many advantages: small size, short generation time (depending on the ambient temperature, from being an egg to become an adult it takes 7 days), a fully sequenced genome and pre-existing libraries of genetic mutants. Studies often use a clear endpoint (death), and this model host can be used in large quantities to facilitate statistical analysis.Numerous studies have revealed significant parallels between the Drosophila immune response and mammalian innate response. The absence of an adaptive immune response permits the study of interactions between pathogens and the host innate immune response in isolation.
Drosophila loss-of-function immune response mutants have been used to examine
the roles of the genes in the response to infection with various pathogens. Transgenic Drosophila has been used to monitor the activation of immune response pathways upon infection and to examine the effects of transgenically expressed pathogen proteins on the host.Drosophila rely solely on an innate immune system to combat infecting microbes and, like mammals, they detect the presence of invading microbes using pattern recognition receptors, which recognize conserved microbial motifs and activate a response that is specific for the type of invading microbes (Igboin et al., 2012) (Figure 7).
Figure 7 – Scheme of humoral immune system
25
Chapter I
The innate immune system consists of both cellular and humoral components. The cellular response involves specialized hemocytes (blood cells), which engage in phagocytosis and encapsulation of foreign microbes. The body cavity of Drosophila
is filled with circulating hemolymph that contains free-floating hemocytes (Figure 7 and 8). Drosophila larvae contain several thousand hemocytes, which can be divided into the following three cell types on the basis of their structural and functional features: plasmocytes, crystal cells and lamellocytes. Plasmocytes represent 90%-95% of all mature larval hemocytes and function in phagocytic removal of dead cells and microbial pathogens. Lamellocytes are relatively large, flat, and adherent cells that primarily function in encapsulation and neutralization of objects too large to be phagocytized. Crystal cells constitute 5% of the larval hemocytes and are nonphagocytic cells involved in the melanization process (Figure 8).
Figure 8 –Scheme of an overview of Drosophila host defense – cellular and humoral responses.
These overview demonstrated that the two response types are connected (Lemaitre & Hoffmann, 2007)
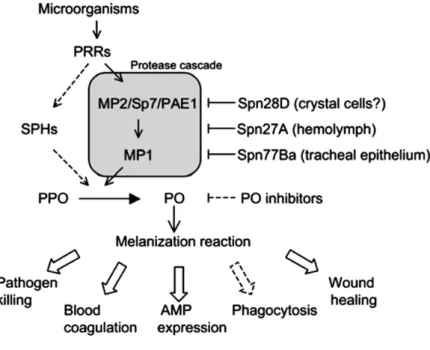
sequestration of microorganisms and production of toxic intermediates that kill the pathogens. Melanization requires the activation of proPhenoloxidase, an enzyme that catalyzes the oxidation of phenols to quinones, which polymerize melanin (Figure 8 and 9) (Lemaitre & Hoffmann, 2007; Meister & Lagueux, 2003). During this process reactive oxygen species are produced, which can harm the host in addition to harming the pathogen (Chambers et al., 2012).
Figure 9 - Model for melanization activation upon microbial Infection. Upon the recognition of a microorganism, a pattern recognition receptor (PRR) is presumed to trigger a protease cascade involving the proteases MP1 and MP2/Sp7/PAE1, which culminates in the cleavage of prophenol oxidase (PPO) to phenol oxidase (PO). The serpin Spn28D controls the release and availability of PO by inhibiting its activation, possibly in crystal cells. Spn27A acts in the hemolymph to inhibit the MP1/MP2 cascade and prevent excessive melanization. Spn77Ba regulates melanization in the tracheal epithelium by inhibiting the same protease cascade. Other studies revealed the involvement of serine protease homologues (SPHs) in activating PO. PO inhibitors limit melanization by directly inhibiting the enzymatic activity of PO. Melanization reaction it also involved in others immune responses such as blood coagulation, AMP expression, wound healing and phagocytosis (Tang, 2009).
27
Chapter I
transcription) pathways (Figure 8 and 10). The humoral response involves the production of antimicrobial peptides (AMPs) through Toll or Imd pathway.
Figure 10 – Drosophila humoral Immune system model – Toll and Imd pathways. These models
Produced AMPs can be classified in seven groups, with differential effectiveness, against fungi (Drosomycin and Metchnikowin), Gram positive bacteria (Defensin) and Gram negative bacteria (Diptericin, Drosocin, Attacin and Cecropin) (Lemaitre & Hoffmann, 2007)The Imd signaling pathway is homologous to the mammalian tumor necrosis factor receptor 1 signaling pathway, and only differs at the level of detection/activation. This pathway regulates the production of AMPs by fat body cells and gut epithelial cells, respectively, in response to primarily Gram negative bacterial infections. The Imd pathway, activated by its receptor peptidoglycan recognition protein (PGRP)-LC and the Imd protein, acts through TAK1, signaling the IkB kinase complex to activate the NF-kB transcription factor Rel, which responds to infection by Gram-negative bacteria (Figure 8 and 10) (Davis & Engstrom, 2012; Kounatidis & Ligoxygakis, 2012; Lemaitre & Hoffmann, 2007).
The Toll signaling pathway is homologous to the mammalian Toll/IL-1 receptor signaling pathway, although unlike the mammalian receptors Drosophila Toll does not directly recognize bacterial components. Like Imd, this pathway regulates systemic AMP expression in fat body, primarily in response to fungal and Gram-positive bacterial infection. The Toll pathway relies on cleavage of the extracellular ligand, Spätzle, followed by signaling through the Toll receptor and its intracellular adaptor protein complex, which contains MyD88, Tube and the Pelle kinase. The Toll pathway is activated in response to Gram positive bacteria, fungi and yeast (Figure 10) (Davis & Engstrom, 2012; Kounatidis & Ligoxygakis, 2012; Lemaitre & Hoffmann, 2007).
The Drosophila JAK-STAT signaling pathway comprises the same components as
the mammalian pathway, although they differ in the number of JAKs (one in
Drosophila and four in mammalians) and STATs (one in Drosophila and seven in
29
Chapter I
maintain epithelial cell homeostasis by regulating stem cell proliferation (Igboin et al., 2012; Lemaitre & Hoffmann, 2007).
5.AIMS ANDSCOPEOFTHESETHESIS
The discovery of communication between bacteria via quorum-sensing, which orchestrate important temporal events during the infection process, provides a novel opportunity to combat bacterial infection. E. faecalis infections constitute a serious problem in the hospital environment and its ability to acquire/transfer antibiotic resistance and virulence factors is limiting the use of antibiotics. New therapeutics are urgently needed, and these may be based on a better understanding of the factors that are important for E. faecalis infection.
Since the Fsr system was described, numerous studies have shown its importance in the infections process. These studies have demonstrated that fsr is not only involved during the E. faecalis infection, but also contributes to enterococcal survival in different environments. However, the exact mechanism by which Fsr contributes to virulence during infection is not entirely clear. Moreover, many strains have been reported to carry the entire fsr and gelE-sprE operons, despite their inability to phenotypically show gelatinase activity isolates (Eaton & Gasson, 2001; Galloway-Pena et al., 2011; Lopes Mde et al., 2006; Nakayama et al., 2002).
This PhD thesis aims at clarifying the contribution Fsr regulon to E. faecalis biology and to host injury using different approaches. Three main questions were addressed in four chapters of this thesis:
- Under which conditions the Fsr system is shut down? (Chapters II and III)
- Which other genes, if any, do Fsr and/or GelE-SprE regulate? (Chapter IV)
- How does Fsr contribute to E. faecalis infection in Drosophila? (Chapters IV and V)
Chapters II and III describe two different conditions in which fsr is shutdown. Chapter II focuses on a particular strain, E. faecalis LN68 (already described by (Lopes Mde
31
Chapter I
phenotype. In a previous transcriptomic study made in our laboratory, the VRE strain V583 was subjected to a therapeutic vancomycin dose. fsr-gelE-sprE genes were found to be affected by that cell wall active antibiotic. In Chapter III we looked for the reason for vancomycin interference with Fsr activity.
In Chapter IV we looked for genes directly and indirectly regulated by Fsr and attributed a role in E. faecalis virulence. To study these genes, we first established
Drosophila as a model to study Fsr virulence. In Chapter V we investigated the
6.BLIBLYOGRAPHY
Antignac, A., Sieradzki, K. & Tomasz, A. (2007). Perturbation of cell wall synthesis suppresses autolysis in Staphylococcus aureus: evidence for coregulation of cell wall synthetic and hydrolytic enzymes. J Bacteriol189, 7573-7580.
Arias, C. A. & Murray, B. E. (2012). The rise of the Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol10, 266-278.
Arthur, M. & Courvalin, P. (1993). Genetics and mechanisms of glycopeptide resistance in enterococci. Antimicrob Agents Chemother37, 1563-1571.
Bassler, B. & Vogel, J. (2013). Bacterial regulatory mechanisms: the gene and beyond. Curr Opin Microbiol16, 109-111.
Bassler, B. L. (1999). How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr Opin Microbiol2, 582-587.
Bayles, K. W. (2007). The biological role of death and lysis in biofilm development.
Nat Rev Microbiol5, 721-726.
Bourgogne, A., Hilsenbeck, S. G., Dunny, G. M. & Murray, B. E. (2006).
Comparison of OG1RF and an isogenic fsrB deletion mutant by transcriptional analysis: the Fsr system of Enterococcus faecalis is more than the activator of gelatinase and serine protease. J Bacteriol188, 2875-2884.
Bourgogne, A., Garsin, D. A., Qin, X. & other authors (2008). Large scale
33
Chapter I
Braga, T. M., Marujo, P. E., Pomba, C. & Lopes, M. F. (2011). Involvement, and dissemination, of the enterococcal small multidrug resistance transporter QacZ in resistance to quaternary ammonium compounds. J Antimicrob Chemother66, 283-286.
Brinster, S., Posteraro, B., Bierne, H., Alberti, A., Makhzami, S., Sanguinetti, M.
& Serror, P. (2007). Enterococcal leucine-rich repeat-containing protein involved in virulence and host inflammatory response. Infect Immun75, 4463-4471.
Chambers, M. C., Lightfield, K. L. & Schneider, D. S. (2012). How the fly balances its ability to combat different pathogens. PLoS Pathog8, e1002970.
Chow, J. W., Thal, L. A., Perri, M. B., Vazquez, J. A., Donabedian, S. M., Clewell,
D. B. & Zervos, M. J. (1993). Plasmid-associated hemolysin and aggregation substance production contribute to virulence in experimental enterococcal endocarditis. Antimicrob Agents Chemother37, 2474-2477.
Cox, C. R. & Gilmore, M. S. (2007). Native microbial colonization of Drosophila
melanogaster and its use as a model of Enterococcus faecalis pathogenesis. Infect
Immun75, 1565-1576.
Cytrynska, M., Mak, P., Zdybicka-Barabas, A., Suder, P. & Jakubowicz, T.
(2007). Purification and characterization of eight peptides from Galleria mellonella
immune hemolymph. Peptides28, 533-546.
Davis, M. M. & Engstrom, Y. (2012). Immune response in the barrier epithelia: lessons from the fruit fly Drosophila melanogaster. J Innate Immun4, 273-283.
de Fatima Silva Lopes, M., Ribeiro, T., Abrantes, M., Figueiredo Marques, J. J.,