UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO
FELIWAY ® SPRAY ASSESSMENT TO HANDLE STRESS IN
CATS IN VETERINARY CONSULTATION
Masters Thesis in Veterinary Medicine
JOANA SOARES PEREIRA
Supervisor: Artur Severo Proença Varejão, DMV, Ph.D. Co-Supervisor: Gonçalo da Graça Pereira, DMV, Ph.D
Universidade de Trás-os-Montes e Alto Douro
FELIWAY ® SPRAY ASSESSMENT TO HANDLE STRESS IN CATS IN VETERINARY CONSULTATION
Masters Thesis in Veterinary Medicine
JOANA SOARES PEREIRA
Supervisor: Artur Severo Proença Varejão, DMV, Ph.D
Co-Supervisor: Gonçalo da Graça Pereira, DMV, Ph.D
Jurors: Carlos Alberto Antunes Viegas, DMV, Ph.D Adelina Gama Quaresma, DVM, Ph.D Artur Severo Proença Varejão, DMV, Ph.D
Acknowledgements
É graças aos meus pais que tenho hoje a oportunidade de apresentar esta dissertação de mestrado e como não podia deixar de ser, o primeiro agradecimento é para eles. Agradeço-vos a amizade, o apoio incondicional, a educação e todos os valores transmitidos.
É também graças à paciência, dedicação e apoio incondicional e à motivação para continuar do meu namorado Gonçalo que esta tese tem hoje forma. “Obrigada” é pouco.
Ao meu Orientador, Professor Doutor Artur Varejão, muito obrigada por ter aceite o convite de orientar o meu trabalho, por estar sempre disponível e pronto a esclarecer as minhas dúvidas.
Ao Professor Doutor Gonçalo Da Graça Pereira ao qual tenho uma lista de coisas a agradecer: por ter aceite ser meu co-orientador, pela oportunidade que me deu em desenvolver com ele este estudo, por toda a disponibilidade, pelo tempo dispensado, pelas ideias e conhecimento transmitido, pelas portas que abriu por mim, por tudo o que fez por este estudo, pela boa disposição e positivismo constante e ainda por aquilo que faz pelos animais, o meu sincero obrigada.
Agradeço muito à Dra. Sara Fragoso, pelo enorme contributo que deu à estruturação do estudo, pelo conhecimento que me transmitiu e pelas ideias que enriqueceram muito o meu trabalho.
To Dr. Alexandra Beck I am deeply grateful for all the work she had during the production process of this study, for her constant availability and willingness to answer my questions, for all her support, for the opportunity she gave me to do this study and for her super kindness.
To Stephane Lavigne, who contributed a lot to this study with his excellent statistical, thank you very much.
Ao Dr. Luís Almeida por me permitir fazer o estudo no seu Hospital Veterinário de Braga, pelo apoio que deu a e ensinamentos que me transmitiu. Agradeço igualmente a toda a equipa do HVB pela maneira carinhosa como me receberam, pela ajuda imediata sempre que precisei, por todo o conhecimento que me transmitiram, pelos bons momentos passados e por se terem tornado bons amigos.
Devo ainda agradecer a toda a equipa de Funcionários, Médicos Veterinários e Professores da UP-VET, do HVUTAD e da Clínica Veterinária do Carandá por tudo aquilo que me ensinaram e por me fazerem querer aprender cada vez mais sobre Medicina Veterinária.
Quero também agradecer à Dra. Luísa Guardão pela energia positiva, por me ensinar a aplicar os meus conhecimentos, pelos conhecimentos que me transmitiu, por me mostrar que a bondade é superior ao restante e pela dedicação que tem aos animais.
Às minhas amigas Martina, Bião, Amy, Judjitxi, Margas e Fifas agradeço as gargalhadas, o companheirismo, o apoio, os momentos de estudo, os almoços, os jantares, os intervalos, os passeios e as noites de copos que, com elas, se tornaram sempre em bons momentos.
A todos os meus amigos e amigas de Braga agradeço por serem precisamente isso, meus amigos. Mafalda, obrigada por me aturares há 20 anos.
Aos meus primos Paulinho, Nuninho e Tia Gabe por me terem dado a oportunidade de viver com eles durante o estágio no Porto e ao meu irmão por me ter dado música (ou comentadores de futebol aos berros) enquanto escrevia “a minha tese”. Tenho a melhor família do mundo.
Por último mas não menos importante, agradeço à Fofa, à Bianca, ao Hoffman, ao Quim, à Quicas e a todos os animais que passaram pela minha vida por me darem, à sua maneira, a inspiração e motivação para querer saber cada vez mais sobre como os poder ajudar.
Abstract
Going to the veterinary clinic is a stressful experience for most cats. This study aimed to investigate how Feliway® Spray might help reduce cats’ stress and ease their handling, when used in consultation.
A randomized, double-blind, placebo-controlled clinical trial was made, using a sample of 87 cats of both sexes, aged between 7 months and 15 years and whose health on consultation showed no signs of disease. A Feliway® spray and a placebo solution spray were tested in two consultation rooms. During the first phase, Feliway® spray was applied in the examination table of one room and the placebo spray in the other. After a washout period of 15 days the sprays allocation was switched. During the first 15 minutes of general examination by the veterinarian, the stress and behavior of the cats were assessed by the observer based on a 7-level Cat-Stress-Score and a 5-point Scale of Handling. Cats exposed to Feliway® were compared to placebo cats, whatever the room.
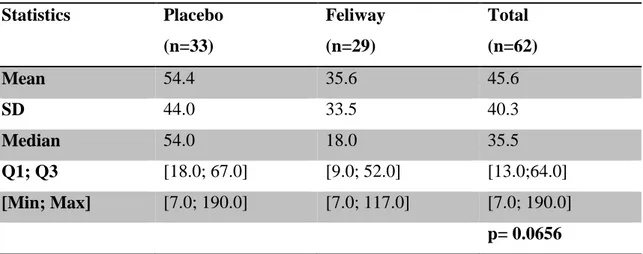
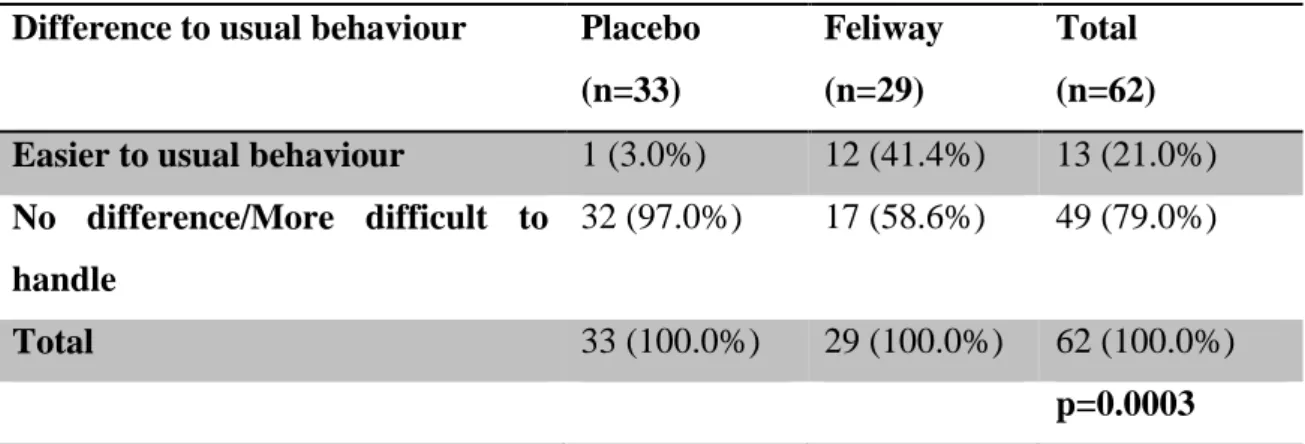
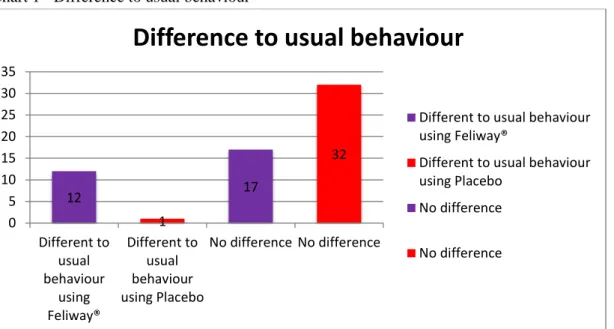
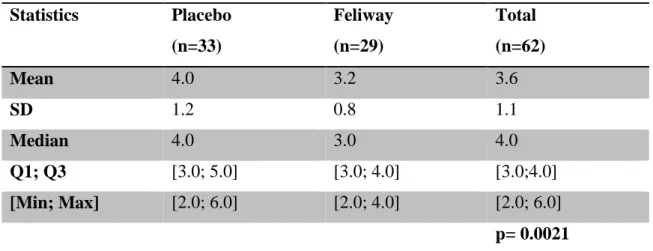
Results show the use of Feliway® leads to statistically significant (p=0.003; Fisher’s Exact test) differences in the behaviour of cats as assessed by their owners, being easier to handle and/or more relaxed compared to previous consultations. Stress-wise, animals exposed to Feliway® demonstrated significantly (p=0.021; Student’s t-test) lower Cat-Stress-Score levels than those treated with placebo. Regarding the Scale of Handling the scoring did not differ in a statistically significant (p=0.1145; Student’s t- test) way between both groups.
Concluding, results suggest Feliway® is an efficient tool to help decrease stress in cats during consultation.
iv Resumo
A ida ao veterinário é uma experiência stressante para a maioria dos gatos. O seguinte estudo teve como objectivo investigar de que modo o Feliway® pode ajudar a reduzir a ansiedade dos gatos e facilitar o seu maneio, quando usado durante a consulta veterinária.
Foi realizado um estudo duplamente cego e controlado por placebo, sobre uma população de 87 gatos de ambos os sexos, com idades compreendidas entre os 7 meses e os 15 anos e cujo estado de saúde na consulta não demonstrasse sinais de doença. Foram testados dois sprays em dois consultórios diferentes, um contendo Feliway® e outro contendo uma solução placebo. Após um período com a duração 15 dias e que serviu para eliminar os vestígios das feromonas anteriormente usadas, a ordem dos sprays foi alterada relativamente aos consultórios. Durante os 15 minutos iniciais da consulta executada pelo Médico Veterinário, o observador avaliou o stresse e o comportamento do gato, tendo como base na “Cat-Stress-Score” composta por 7 níveis e a escala “Escala de Maneio” de 5 níveis. Os gatos expostos a Feliway® foram comparados aos gatos expostos à solução contendo placebo, em ambos os consultórios. Os resultados demonstram que, segundo a opinião dos proprietários, a utilização de Feliway® leva a diferenças estatisticamente significativas no comportamento dos gatos (p=0.003; Teste Exacto de Fisher), sendo que os gatos sob a influência da feromona tiveram mais fácil maneio e/ou estavam mais relaxados comparando com consultas anteriores. Em relação ao stresse, os animais expostos ao Feliway® evidenciaram níveis de Cat-Stress-Score stresse significativamente mais baixos (p=0.021; Teste t de Student) que os gatos sob a influência de placebo. Relativamente à Scale of Handling, as diferenças de pontuação obtidas por ambos os grupos não se mostraram estatisticamente significativas (p=0.1145; Teste t de Student).
Em suma, os resultados sugerem que o spray Feliway®, é uma ferramenta eficaz na redução do stresse nos gatos durante a consulta veterinária.
v Table of contents Acknowledgements ... i Abstract ... iii Resumo ... iv Table of contents ... v
Index of tables ... vii
Index of charts ... viii
Index of images ... ix
Index of abbreviations and symbols ... x
Introduction ... xi
Chapter I – Literature review... 1
1 – The Feline patient ... 1
1.1 - Origins and Domestication ... 1
1.2 - Social Organization ... 2
1.3– Inter-cat and cat-human relationship ... 4
1.3.1- Relationship with conspecifics ... 4
1.3.2 – Relationship with humans ... 5
1.4 - Factors influencing cat’s socialisation towards humans ... 8
1.5 - Communication ... 10 1.5.1 - Visual Communication ... 11 1.5.2 - Vocal Communication ... 15 1.5.3– Olfactory Communication ... 17 1.5.3.1– Pheromones ... 18 1.5.3.2 – Urine marking ... 22 1.5.3.3 - Rubbing ... 24 1.5.3.4 – Scratching ... 24 1.5.3.5 – Faecal marking ... 25 2– Stress ... 26
2.1– Definition and physiology ... 26
2.2– Stressors ... 29
2.3– Behavioural measures of Stress ... 31
3– Decreasing cats’ stress at the veterinary practice ... 34
4– Pheromonatherapy in cats ... 38
vi
4.2– Applications of F4 ... 42
Chapter II – Feliway® spray assessment to handle stress in cats in veterinary consultation ... 45
1 - Objectives ... 45
2 - Materials and Methods ... 45
3 - Results ... 55
3.1- Analysis of the demographic data ... 55
3.1.1 - Age and waiting time ... 56
3.1.2 – Gender ... 57
3.1.3 - Spaying/Neutering condition ... 57
3.1.4 - Indoor and/or Outdoor lifestyle ... 58
3.1.5 - Breeds ... 59
3.1.6 - Reason for consultation ... 59
3.2 - Efficacy of Feliway® spray versus placebo spray ... 60
3.3 – Influence of room A and room B... 65
3.4 - Efficacy of placebo versus “no spray” ... 70
4 - Discussion ... 73
5 - Conclusion ... 77
Index of tables
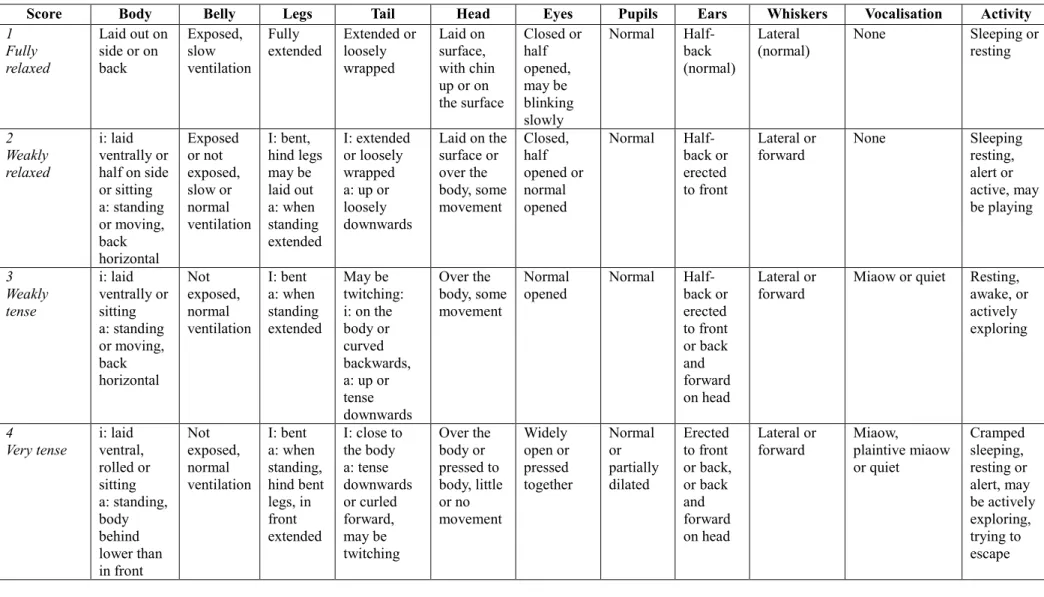
Table 1 - Definitions of seven-level Cat-Stress-score (from Kessler & Turner 1997), i: (or
unspecified) = cat is inactive; a: = cat is active ... 48 Table 2 - Scale for qualifying the handling of cats during consultation ... 50 Table 3 - Phases of the trial ... 53 Table 4 - Comparison of the age in months between the populations tested with "Placebo" and "Feliway" ... 56 Table 5 - Comparison of the waiting time in minutes between the populations tested with
"Placebo" and "Feliway" ... 57 Table 6 - Population distribution by sex between “Placebo” and “Feliway” ... 57 Table 7 - Population distribution by spaying condition between "Placebo" and "Feliway" ... 58 Table 8 - Population distribution by indoor or/and outdoor lifestyle between "Placebo" and
"Feliway" ... 58 Table 9 - Population distribution by breed between "Placebo" and "Feliway" ... 59 Table 10 – Difference to usual behaviour of the cats under the influence of “Placebo” and
“Feliway” ... 60 Table 11 - Sweat from the cat’s paws under the influence of “Placebo” and “Feliway” ... 61 Table 12 – Comparison of the stress scores of the two groups of cats tested with “Placebo” and
“Feliway” ... 62 Table 13 - Stress scores “under or equal to level 3” and “scores above level 3” in the CSS scale,
of cats tested with "Placebo" and "Feliway" ... 63 Table 14 - Handling scores “under or equal to level 3” and “above level 3” in the Scale of
Handling, of cats tested with "Placebo" and "Feliway" ... 64 Table 15 – Descriptive analysis of the Efficacy data by Room for the global population tested
with placebo ... 66 Table 16 - Descriptive analysis of the Efficacy data by Room for the global population tested
with Feliway® ... 68 Table 17 – Odds Ratio Estimates for the influence of the room on the behaviour of the cats .. 69 Table 18 – Odds Ratio Estimates for the influence of the room on the CSS ... 70 Table 19 - Descriptive analysis of the efficacy data for the global population of cats tested with "Placebo" and with no spray “N/A” ... 71
Index of charts
Chart 1 - Difference to usual behaviour ... 61 Chart 2 - Sweat from the cats' paws ... 62 Chart 3 - Comparison of the stress scores of the two groups of cats tested with “Placebo” and
“Feliway" ... 63 Chart 4 - Stress scores “under or equal to level 3” and “scores above level 3” in the CSS scale,
of cats tested with "Placebo" and "Feliway" ... 64 Chart 5 - Handling scores “under or equal to level 3” and “above level 3” in the Scale of
Handling, of cats tested with "Placebo" and "Feliway" ... 65 Chart 6 - Descriptive analysis of the Efficacy data by Room for the global population with
Placebo ... 67 Chart 7 - Descriptive analysis of the efficacy data by room for the global population with
Feliway®... 69 Chart 8 - Descriptive analysis of the efficacy data for the global population of cats tested with
Index of images
.
Image 1 - Typical offensive posture (adapted from Beaver, 2003) ... 12 Image 2 - Tail up position (adapted from Bowen et al., 2005) ... 13 Image 3 - Whole body postures. Aggression increases from left to right and fear from top to
bottom. (Adapted from P, Leyhausen 1979 Cat behaviour. Garland STMP Press, New York. In Bowen et al., 2005) ... 14 Image 4 - Facial postures. Aggression increases from left to right and fearfulness from top to
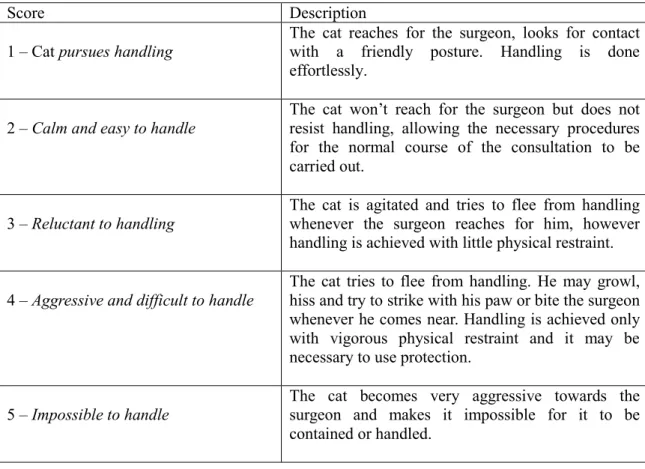
bottom. (Adapted from P Leyhausen 1979 Cat behaviour. Garland STMP Press, New York. In Bowen et al., 2005) ... 15 Image 5 - Typical posture adopted when urine marking (adapted from Beaver, 2003)... 23 Image 6 - Examining a cat while it remains in the bottom (adapted from Rodan, 2011) ... 36 Image 7 - Case Report Form "Feliway assessment to handle stress in cats in veterinary consultation"……….51
Index of abbreviations and symbols
% - Percentage
ºC – Degrees Celsius
ACTH – Adrenocorticotropic Hormone
CSS – Cat-Stress-Score
DSH – Domestic Short Hair
FIC – Feline Idiopathic Cystitis
HPA - Hypothalamic–Pituitary-Adrenal
HVB – Hospital Veterinário de Braga
Introduction
The following Masters Dissertation is composed of two parts: one which comprehends a bibliographic revision on some concepts regarding cat “stress”, “pheromones” and “handling”, and another one through which I present the study carried out on the “Feliway Spray assessment to handle stress in cats in veterinary consultation”.
The domestic cat population has been increasing considerably, in such way the number of cats now exceeds the number of dogs in Europe and in the United States of America (Beaver, 2003; Genaro, 2005). Some reasons pointed out for this fact are the owners’ lifestyle in the Western World, as well as the cat’s ease to adapt its behaviour to its owner’s living, being these criticisable by many behaviour specialists (Beaver, 2003; Heath, 2005). Nevertheless, this growth in the number of cats implies a concomitant increase in the casuistry of this species in veterinary clinics (Genaro, 2005). Still, and although the vast majority of owners consider their cat as a member of their family, the number of cats observed in veterinary clinics is substantially lower than the number of dogs. According to the owners, stress and fear, as well as the difficulty in carrying the cat are the most common reasons for this to happen (Vogt et al., 2010).
A trip to the veterinary clinic is a stressful and fear engaging experience for most animals, and especially for cats, as they feel threatened as they enter a new and confined environment which reduces their normal active behavioural response, due to the contact with unfamiliar animals and people or a previous negative experience. Anxiety can worsen with pain or physical restraint limiting the examination of the animal, thus the need for the use of chemical restraint, which is not risk free and might bring along possible complications (Carlstead et al., 1993a; Kessler et al., 1998; Mills, 2002; Mills et al., 2005). Fear also can change some physiological parameters, making it difficult to truly assess the wellness of the animal (Quimby et al., 2011).
The major role of the veterinarians is to assure the well-being of animals “relieving suffering, whether it is related to physical or emotional pain”, as stated by Beaver and colleagues (2004). By preventing and treating behavioural problems, we have the chance to protect and strengthen the human-pet-veterinary bond (Beaver et al., 2004).
The proper handling of the cat during the consultation is of major importance for reducing anxiety thus increasing the ease of handling making it more pleasant for the animal, owner and veterinary surgeon (Overall et al., 2004; Rodan et al., 2011).
Five functional feline pheromone fractions (F1 – F5) have been isolated from the facial secretions of cats. Two of these – F3 (Feliway®; CEVA Santé Animale) and F4 (Felifriend®; CEVA Santé Animale) were artificially synthesised and commercialized as a possible treatment for stress-related behaviours in cats (Mills, 2005b).
Feliway ® is an emulsion that contains fatty acids (oleic acid, azelaic acid, pimelic acid, palmitic acid) and extract of Valeriana officinalis (which serves to maintain the attraction of cats to this synthetic pheromonal analogue at greater distances) (Pageat, 1998). It has already been studied by several authors and its effectiveness proven on both sexual and reactive urine spraying (Pageat, 1997; White et al., 1997), reducing the signs and the recurrences of feline idiopathic cystitis (Gunn-Moore et al., 2004), preventing behaviour disorders like escape in cats moving from their houses to holiday homes (Pageat et al., 1997), reducing stress during transportation (Gaultier et al., 1998), helping to calm cats and improving interest in food and food intake in unfamiliar surroundings in hospitalised cats (Griffith et al., 2000; Kronen et al., 2006).
Could this pheromone act as a supplement to help in the handling of cats and reducing their fear and anxiety while they are at the vet?
This study focuses in the attempt to handle and decrease the anxiety of cats during their stay in the veterinary clinic which contributes positively to both the course of the consultation and the welfare of the animal.
The aim of this research is to investigate to what extent the use of a Feliway® spray (synthetic copy of the feline facial pheromone) affects and helps to decrease stress as well as to ease their handling during veterinary consultation.
Chapter I – Literature review
1 – The Feline patient
1.1 - Origins and Domestication
Several species of small cats of the genus Felis, including the wildcat Felis silvestris, evolved in Asia three million years ago. Felis silvestris was widely distributed through Europe, Middle East and Asia (Lipinski et al., 2008; Vaughan et al., 2011). Driscoll and colleagues (2007) analysed mitochondrial DNA from 979 cats including wildcats, Sand cats, Chinese desert cats, and domestic cats (feral and purebreds) ailing from three continents and concluded that domestic cats had their origin in five female ancestors of Felis silvestris lybica (wildcats) from the Near Eastern.
It has been a long time since animals have been connected to humans (Turner, 2000a). Although Egyptians were thought to be responsible for domesticating the cat about 4000 years ago, recent archaeological findings, in Cyprus, showing a cat buried alongside a human, aged approximately 9500 years, provide evidence that the human-cat relation has existed for longer than originally thought (Vigne et al., 2004; Bernstein, 2005). Driscoll and colleagues (2007) also found strong evidence that cat domestication occurred in the Near Eastern part of the Fertile Crescent.
It is believed that humans have done little more for the cats taming besides living in harmony, providing food and the opportunity for successful reproduction, bringing to life the recently hypothesis that this species has undergone a process of natural “self-domestication” rather than artificial selection (Beaver, 2003; Driscoll et al., 2009). In Egypt, the cat-human relation was based on a mutual association where the cats were kept to control the rodents that destroyed the crops, stole from granaries and later to fish and hunt (Beaver, 2003).
The domesticated cat slowly spread to the Asian and European continents with the expansion of the sea routes. It is known to have arrived in Britain between 300 and 500 AD and to the North America in the seventeenth century (Beaver, 2003).
Throughout history, cats have been linked to religion: in Egypt, Bastet, the cat goddess represented fertility and health; in the Islamic countries, being Mohammed’s favourite animal, those who harmed or mistreated cats would be punished; in Europe, the cat was believed to have protected the Christ child from the Devil’s mouse and also
was associated with the moon goddess Diana. During the Middle Ages, many cats were exterminated, as well as man who showed compassion for them, due to their association with witchcraft. This situation reversed when a plague of brown rat was brought by the crusaders around 1600 AD and cats were a rat-control method being slowly accepted by people. They were also kept in monasteries to protect manuscript from rodents (Beaver, 2003; Genaro, 2004).
Later in 1800, with Pasteur’s finding about bacteria the consciousness about cleanliness grew and cats earned the status of a clean animal, becoming the favourite animal of merchants (Beaver, 2003).
Nowadays, although the exact population data is inaccurate due to wild populations, the number of cats has been increasing in part because of their adaptability to small houses. It is also achieving, much like other pets, many important roles like research animal for aggression, neurology, ecology and aging studies, as well as companion animal providing companionship, motivation and the sensation of safety (Beaver, 2003; Genaro, 2004; Rodan, 2010).
1.2 - Social Organization
The domestic cat’s ancestor, Felis silvestris lybica, was a solitary and territorial species. Nowadays, the domestic cat is very flexible and may adopt a series of life-styles: a self-sufficient and independent social life, a group living social life in colonies of free-ranging cats not depending on humans or a group living social life in colonies of free-ranging cats depending on humans to get food resources. Lastly, we have domestic cats living only indoors and completely dependent of their owners (Miller, 1996; Turner, 2014).
Food resources affect the free-ranging domestic cats spatial pattern and population density, affecting their social organisation, socialization towards conspecifics and humans, birth rate, mating system, hunting behaviour and even prey populations (Turner, 2014). According to food abundance and distribution, cats either live alone or in colonies. Large groups are formed where food is abundant and small ones where food is scarce. When food is too widely dispersed to support a group, cats tend to live a solitary life (Liberg et al., 1988; Yamane et al., 1996; Natoli et al., 2001; Crowell-Davis et al., 2004; Turner, 2014). During the female’s oestrous season, males expand
significantly their mean home range size which correlate with the number of females they courted, meaning that, for tomcats, the mating resource is more important determining home range size than food resource (Yamane et al., 1994).
Some authors (Natoli et al., 2001; Crowell-Davis et al., 2004; Sousa et al., 2010) agree when saying that, as in many other groups of animals, cats form a nonlinear hierarchy with ties and reversals within their colony. These hierarchies affect access to food and shelter, mating and frequency of affiliative and agonistic behaviours (Natoli et al., 2001; Crowell-Davis et al., 2004; Sousa et al., 2010).
The core of the groups is composed by females and their descendants, forming a matrilineal colony which is based on a social structure built on the co-operative relationships between the females of the group which defend their feeding sites from females of other colonies, care and rear the kittens together (Yamane et al., 1994; van den Bos, 1998; Bradshaw et al., 2012). Males may either wander between several colonies or be more attached to one single colony (Davis et al., 2004; Crowell-Davis, 2005).
Cats within groups form friendly relationships with certain other cats (“preferred associates” - cats that are found close to each other more often than they are to other conspecifics), engaging in a series of affiliative behaviours like touching noses, social grooming (allogrooming), social rubbing (allorubbing), cuddling and sleeping next each other. Other cats are loosely attached to the members of their colony. As a consequence cats societies are complex (Crowell-Davis et al., 2004; Crowell-Davis, 2005; Sousa et al., 2010).
The absence of post-conflict reconciliation mechanisms and a repertoire of behaviours designed to increase distance between individuals, make cats avoid outsiders, through spatial distribution and timing-patterns, and tolerate other cats of the group, limiting their interactions and thus reducing agonistic behaviours (van den Bos, 1998; Rochlitz, 1999; Bowen et al., 2005). Despite this, whenever fewer resources available for each individual, conflict may arise due to competition. Since cats can distinguish members of their group from strangers, serious disruption of the social order in the colony can occur if a new cat joins the group, both in colonies of free-living cats and in the domestic household (Crowell-Davis, 2005) .
Yamane and colleagues (1997) studied a colony of cats and found that the factors affecting the feeding order were age among the males and body weight among the females. They also concluded that adult male and female cats permit kittens to skip
stages in the feeding order. In other study, Yamane and colleagues (1996) found that severe fights occur among males for the mating resource and thus body weight was very important to their success.
1.3– Inter-cat and cat-human relationship
Different to what it was thought many years ago, when cats were considered an asocial carnivore that preferred to be alone and had no need for companionship, it is now known that the cat is a social species that regularly interacts with conspecifics and contact frequently and effectively with humans (Voith et al., 1986; Crowell-Davis, 2005).
The sociability of cats towards people and conspecifics is influenced by many factor like poor early socialization, exposure to other cats and people and genetic influences (McCune et al., 1995; Crowell-Davis et al., 2004).
1.3.1- Relationship with conspecifics
Preferential associations between individuals from the same colony are formed based of their familiarity and proximity during the socialization period as kittens (Bradshaw et al., 1999).
The preferred associates exhibit a greater number of affiliative behaviours and remain much more frequently close to each other, approximately less one meter, in different contexts and locations (Bradshaw et al., 1999; Curtis et al., 2003). These affiliative behaviours include: 1. nose touching (a greeting behaviour, observed most commonly between preferred associates); 2. allogrooming (cat licks a conspecific in the head or the neck, often when the cats are resting together)– for reinforcing bonds between related individuals and provides stability within the group; 3. allorubbing (when, for several minutes, two cats rub their head, bodies and tails against each other) – besides the purpose of exchanging scents this behaviour is believed to maintain friendly bonds between two cats of the group; 4. playing – it is thought to be relevant for establishing and maintaining social bonds because it happens throughout the life of the cats (peaking between four weeks to four months) even in poor nutritional
conditions and for social learning; and 5. resting together – cats that rest together in close physical contact even when there is room for them to spread out and even if there is no need for thermoregulation, choosing to rest together because of social bonds (Crowell-Davis et al., 2004; Bowen et al., 2005; Crowell-Davis, 2005).
In a study consisting of 60 households with dyads of neutered indoor cats of both genders and where no food, mating nor shelter are limited resources, Barry and colleagues (1999) found no correlation between gender, household type or between recipients in affiliative and aggressive behaviour.
On the other hand, if food, mating or other resources like elimination sites (litter boxes in a household scenario) are limited, there is a change for competition. The organisation within groups can mediate access to resources and the expression of aggression. When a cat defers or gives away to another is setting forth to a consequence of previous interaction with that conspecific that could be seen as a dominant to him. Submission patterns are replaced by deference patterns. This is highlighted by behaviours such as walking around the other cat, waiting for the other cat to pass before moving into an area, retreating and avoiding eye contact (Bernstein, 1981; Bowen et al., 2005; Crowell-Davis, 2005).
Like many other mammals, cats need to experience with their conspecifics to develop appropriate specie-specific social behaviour. There is no wonder that, when adopt as kittens and kept in a single-cat household they miss important learning in social behaviours and when adult, such cats may exhibit lack of knowledge of how to interact with conspecifics, arising aggression and other behavioural problems when an attempt to introduce another cat in the household is made (Crowell-Davis et al., 2004).
1.3.2 – Relationship with humans
Cats obviously benefit from living with humans. Some of the benefits include reliable food supplies, veterinary care, protection from predators, good environmental conditions, and decreased stress levels (Carlstead et al., 1993a).
Likewise, as reviewed in Friedmann and colleagues (2010), Kruger and colleagues (2010) and Fine and colleagues (2010), there is a number of studies that suggest that humans benefit physiologically - blood pressure control, relaxation, less levels chemicals associated with anxiety, improved survival and longevity after heart
attacks, and psychologically – cats seem to lower negative moods, from pets’ companionship (Turner et al., 2001; Turner et al., 2003).
There is a variation in bonding and interaction styles between people and cats. For some cats, owners are just a food resource. Other cats greet their owners when they come back home and some cats even join their owners for a ride. This variation depends on the strength of the bond between people and their pets (the urge of the cat and owner to be next to each other), the quality of the attachment they have (related to the sense of safety and calmness they provide to each other in case of stressful events), what activities they do together (interactions and behavioural rituals they develop). This set of variables together will form the bio-psychological and behavioural syndrome of relational needs, attitudes, affects and interactions called “relationship” (Bernstein et al., 2014).
A study by Edwards and colleagues (2007), shows that cats spend more time in locomotion and exploring when the owner is present than when are left alone or with a stranger. Since, in cats, locomotion is an indicator of attachment and stillness an indicator of stress, these findings demonstrate that there is an attachment between the cat and their owners (Carlstead et al., 1992; Kessler et al., 1997; Kessler et al., 1998; Edwards et al., 2007) and that cats are less likely to be destructive then dogs when suffering from separation anxiety (Edwards et al., 2007).
Facial marking is considered to be a special means of communication in which cats deposit pheromones that help them differentiate between familiar or unfamiliar objects or humans (Pageat, 1997; Pageat et al., 1997). Edwards and colleagues (2007) found that owners are marked more frequently than strangers which means that cats recognize their owners and are attached to them.
Mertens (1991), observed the interactions between owners and cats in the household environment and found that the interactive behaviours of both partners in a human-cat dyad increased with increasing duration of human presence at household. Gender as well as age also affected the relationship: women-cat dyads had the best relationship and juvenile-cat dyads had the worst one. Although the housing condition affects the durations of the cat’s presence at home it didn’t seem to affect the interactions between cat and human. Mertens (1991) noticed that in general, interactions were few in number and lasted less than one minute. People tended to approach the cat more frequently than cats approached the owner and whenever the cat was the one to approach the human, then the interaction would last longer.
Bradshaw and colleagues (1996), also studied cat behaviour and their interactions with owners at feeding occasions and found that cats have visual, vocal tactile and olfactory signs directed at the owner such as rub head/flank/tail, look, miaow and walk past owner and that these signs are shown in a higher rate when the cat is anticipating food than in other occasions when owner and cat are in close proximity, although some cats will still perform these after that phase.
Wedl and colleagues (2011) study, which highlights that the gender of the cat doesn’t influence the patterns of dyadic behaviour, contradicts Bernstein et al. (2014), in which the authors found evidences of sex-related differences in cats behaviour towards their owners. In the mention study the conclusion shows that, although all cats were neutered, male cats were more expressive, more communicative and interactive with the owner.
Until this moment, we have been analysing the cats’ characteristics that could influence the relation with humans. But is there any evidence that some human characteristics can also influence this relation?
Wedl and colleagues (2011) corroborate Mertens (1991) results, when concluding that: female owners tend to interact more frequently with their cats. Interestingly they found that the personality of the human affects temporal patterning of the behaviour between the human and the cat, meaning that people scoring high on neuroticism had fewer and less complex patterns of interaction with their cat.
Adameli and colleagues (2005), stated that the care provided to the cat, its behaviour as well as the amount of time the owner spends with the pet is mainly, but not exclusively, influenced by the owner’s gender. They also found that the cat’s features do not influence its own behaviour as much as the owner’s features (such as gender) do. According to these authors on the list of the owner’s features that do influence the amount of care give to the cat also are: previous experience with cats, level of education and number of friends as well as emotional bonds to people. In fact, cats are more often correctly fed by individuals who claim to meet a smaller number of friends (less than 5 a month), and to maintain fewer bonds (5 to 10). The fact that these individuals are less prone to receiving advices from people with no experience, such as neighbours and friends, may back this statement. People with a lower education level do not brush theirs pets as owners whose education reached a higher level. In terms of experience with cats, they found that owners that have had cats before are more likely to let their cats express their behavioural needs such as clawing.
Cats whose ownership is shared by a larger number of elements of the family, receive a more appropriate veterinary assistance, as the responsibility for the health and comfort of the cat is shared and embraced by every member of the family. Still on the family’s page, the cat’s behaviour towards its owner is also related to the number of members and children in the family and number of people who look after him, wherein small families with no children make place for more sociable cats, which is probably explained by the facts that these households are able to create deeper and stronger bonds with the cat, are more organized regarding pet management and make more room for attention towards the cat. Also, the absence of children in the household could mean a quieter environment to which cats are very sensitive to. Cats are also very sensitive to the lack of a definite routine (Adamelli et al., 2005).
One of the most common interactions people have with their cats is petting (Bernstein et al., 2014). Owners can identify specific body parts and the areas in the household where their cats prefer to be petted (Bernstein, 2000).Some cats lead their owners to pet them in certain areas where they can get the most satisfying petting and they show clearly positive and negative responses according to the area their being petted (Bernstein, 2000; Soennichsen et al., 2002). In Soennichsen et al. (2002) study, cats evidenced more positive and fewer negative responses to petting in the temporal region and more negative responses to petting in the caudal region, perioral and non-gland areas.
Regarding first encounters, Mertens and collegues (1988) concluded that, regarding the influence of human gender on human-cat interactions, men tend not to vocalise as much as women do on first encounters, and cats are more prone to approach women than man.
1.4 - Factors influencing cat’s socialisation towards humans
Plasticity to learning about environmental stimuli is more pronounced during particular periods of development called “sensitive periods” (Bateson, 1979).
A study by Kuo (1930), in which the author intended to investigate the instinctive behaviour among cats, showed interesting findings: cats raised in isolation or in a killing rat environment killed 54% and 85% rats respectively, while cats raised in the same cage as rats didn’t kill any rat. This shows that cats raised with rats become
“socialised” to them and those experiences during the early ages of cats profoundly affect the future behaviours that they will display.
Socialisation is a term used to describe the early sensitive period for learning known as the “socialisation period”, as well as the process of interaction between animals and humans in adulthood (Casey et al., 2008).
Karsh and colleagues (1988), identified the socialization period of cats beginning at two weeks of age and finishing approximately at seven weeks of age. They also reported that increasing, fifteen to forty minutes per day, the period of time of handling will significantly increase the period of time kittens, at fourteen weeks of age, spend in close contact with people.
The socialisation in early ages of the cat, particularly during the first weeks of age, the amount of handling received, the number of handlers and the style of handling, influence kitten’s willingness to approach humans (in abstracts:Wilson et al., 1965; Collard, 1967; Reisner et al., 1994; and reviwed in McCune, 1995).
In a study with the objective to reduce the stress and aggressiveness associated with fear of handling in laboratory cats, in litters of five different fathers, Reisner and colleagues (1994), found a remarkable influence of the kitten’s father on the socialisation towards humans, the way kittens respond to restraint for examination and venipuncture.
McCune (1995), investigated the influence of genetics and early socialisation in the outcome of a cat’s friendliness and boldness to humans and objects. For that she studied thirty-seven kittens. These kittens were the offspring of two fathers: one father was considered to be friendly towards people and the other father was considered unfriendly. From two to twelve weeks of age these kittens were either handled (socialised) or received no handling (unsocialised). By the time the kittens turned one year oldthey went through three experiments: response to a familiar person, response to a stranger and response to a novel object. McCune (1995) clearly concluded that: 1. early handling produces significant differences in friendliness in cats towards people; and 2. early handled kittens become friendlier adult cats. It was also clear that the descendants of the friendly father were friendlier to people and less anxious when unfamiliar people touched them. Unsocialised and unfriendly-fathered cats were more likely to hiss, adopt a flattened posture and to hide, expressions of aggression (Bowen et al., 2005), when in proximity to a person.
The benefits of increasing socialization, in the early ages of kittens, beyond those provided ordinarily in cat shelters/rescue/re-homing centres, are highlighted in Casey et al. (2008) study as they found that kittens receiving additional socialisation were evidently less fearful than the ones that had been through the normal interaction with caretakers. They were also able to conclude that the bonding between cats that have had been early socialised and their owners was stronger.
The acceptance of conspecifics or individuals of other species is a special learning process and that’s why puppy socialization classes also known as “puppy preschools” and “kitten kindergarten” or “kitten classes” are an important way to socialize puppies and kittens, exposing them to various stimulus in a nonthreatening way and teaching good manners (Seksel et al., 1999; Seksel, 2008). In these classes kittens are allowed to play together and to be exposed to people of different ages and genders. This exposure together with positive reinforcements, helps the kittens better adapt to possible changes that may occur, making the cat prepared to accept without fear new environments and experiences (Rodan et al., 2011).
As reviewed in Bernstein et al. (2014), a variety of factors contribute for cats behavioural pattern towards humans: genetics, early socialisation, housing conditions, owner attachment and other owner-related factors.
1.5 - Communication
Domestication may have favoured individuals that were capable of communicating effectively with their owners. Because cat domestication is relatively recent, their communicative capabilities are likely to have yet reached the peak of its evolution, thus making it possible for both intra and interspecific communication to still be evolving (Brown et al., 2014).
Cats communicate through three main categories of signalling: visual (often subtle but very important for inter and intraspecific communication), vocal (highly conditioned for the domestic cat) and olfactory (with a very important role in the social communication but often unappreciated by humans) (Beaver, 2003; Bowen et al., 2005).
Regarding the domestic cat, communication, mainly vocal communication, must be interpreted cautiously as it might result of a learned process of communicating with
the owner, leading him to perform certain tasks as feeding or opening doors/windows (Bowen et al., 2005).
1.5.1 - Visual Communication
In spite of their limited colour perception and limited ability to focus from near to distant objects than humans do, cats are able to tell differences in shape, texture and size of objects. Furthermore, they seem to be unable to focus objects sitting closer than a 25cm distance (Brown et al., 2014).
Cats also show a more reduced ability to detect objects moving within their visual field than humans, who are able to detect motion on objects moving 10 times slower. On the other hand, regarding rapid movements, to which their vision is highly adapted, these are prone to trigger much more of a response from cats. A fast-moving object (such as a mouse or other prey) is easily followed by rapid movements of the eye called saccades during which the object is continuously tracked while the sensory information is carried and processed by the nervous system (Rodan, 2010; Brown et al., 2014).
The use of visual signals by cats is obviously influenced by their elements of vision, which are more limited in comparison to humans’, in some cases, and more evolved in others. The visual displays based on colour contrast are very ineffective for cats due to their little ability to differentiate colours (Brown et al., 2014).
Body language is undoubtedly an important way of visual communication for cats, designed to avoid conflict instead of inciting it. It is also important for veterinarians and those dealing with feline behaviour problems to understand how cats use their body posture and their facial expressions to communicate (Beaver, 2003; Bowen et al., 2005; Rodan, 2010).
There are some behavioural displays with the ability to create a very well distinguishable image, preventing misunderstandings, which might have evolved from the need to communicate through clear and unambiguous signals when facing encounters with unknown conspecifics (Beaver, 2003; Bowen et al., 2005; Brown et al., 2014).
Facial expressions are clearly the most important way of passing on a clear message, associated with its body posture that gives the first impression to an approaching cat (Bowen et al., 2005).
One of the best known postures is the defensive one where cats raise their hairs along the back, flatten their ears caudally, lean their whiskers against the cheeks, stretch their labial commissures caudally so that the teeth become visible, arch their back and stand sideways on to a potential opponent also known as the inverted U shape. This comes has a way of looking twice its size to the opponent cat. They also engage in a slow moving behaviour, which serves to prevent chasing by the attacker, and in a sideways position, which allows to keep its adversary in view. This behaviour is not always considered by the cat as an appropriate response and so, when they feel threatened, since they prefer to avoid and flight from conflict, cats often shrink to the smallest possible dimensions and try to hide (Beaver, 2003; Bowen et al., 2005). The classic offensive posture is seen as a direct visual contact, with whiskers forward, body angled forwards and the tail might do front-rear movements or body along the ground with anterior tilt, piloerection commencing in the thoracolumbar region that extends caudally, rigid tail with caudal direction with the tip bent down and lateral movements of the head end (Beaver, 2003).
The tail plays an important role in the visual communication: rapid movement of the tail indicates that the cat is agitated, and although it has been associated with anger and possible attack, in fact the wagging of the tail indicates agitation and a state of emotion which might in fact lead to aggression if the source of disturbance does not cease. Both a concave and a lowered tail position can indicate an aggressive state and are often associated with conflict. The “bottlebrush” tail is normally associated with fear and defence. In the opposite state, a cat which approaches another cat or a human with an upright position, also called “tail up”, is friendly greeting, usually followed by a touching noses or rubbing behaviour, or offering an invitation to be stroked and played with. The tail up also serves to test the potential reaction of the conspecific to an attempt of rubbing behaviour, thus avoiding trouble by being rejected (Bowen et al., 2005; Cafazzo et al., 2009).
Image 3 - Whole body postures. Aggression increases from left to right and fear from top to bottom. (Adapted from P, Leyhausen 1979 Cat behaviour. Garland STMP Press, New York. In Bowen et al., 2005)
Although the body postures adopted give important information about the overall state of the cat, it’s the face that gives the most detailed information and clues of what to expect from it. The shape of the ears, the eyes and the position of the whiskers are effective and rapid vehicles for sending a message but these should always be interpreted with information based on the whole body language and the environment (e.g. low light makes the pupil dilate). Dilated pupils are commonly associated with fear or high arousal (unrelated with fear) and narrow ones are a sign of contentment. Blinking is believed to be connected with the seeking of reassurance when the cat is in a tense environment. On other hand, staring is believed to be a sign of an assertive individual and prolonged eye contact is used to intimidate an opponent. The ears are yet another indicator of the feline emotion and intention. Whenever they are folded sideways and downwards the cat is trying to avoid confrontation and preparing to defend itself. When the ears are rotated backwards and flattened against the head the cat is ready to attack. Cats also use their ears to test reactions of the opponent, meaning they alter their ear position several times through quick small movements (Bowen et al., 2005; Rodan, 2010).
Image 4 - Facial postures. Aggression increases from left to right and fearfulness from top to bottom. (Adapted from P Leyhausen 1979 Cat behaviour. Garland STMP Press, New York. In Bowen et al., 2005)
The visual signals can also remain for a long time even if the cat is no longer present, which happens with the scratch marks. These can be performed on both horizontal and vertical objects with the forelimbs extended. This marking behaviour also serves to help the maintenance of the claws, stretch and exercise muscles and tendons from the forelimbs and to deposit secretions from the glands of the foot pads (Beaver, 2003; Bowen et al., 2005; Crowell-Davis, 2005).
1.5.2 - Vocal Communication
This type of communications is used to transmit general messages (Beaver, 2003). The variety of vocalizations of cats comes from a voiced column of air during inhalation or exhalation, by means of tensional changes in the throat and mouth, speed and energy of the column of air, extent, time and rate of the opening of the mouth, meaning that vocalization is due to the constitution of the physiology of the cat (Moelk, 1944).
Association with man affects the strength and temper of the vocalizing effort but not directly through a choice among vocal patterns (Moelk, 1944).
Cats normally tend to vocalize while they are kittens (Bowen et al., 2005).
There are at least sixteen different distinct vocal signals known in the cat but their exact meaning is still to unveil (Bowen et al., 2005). The vocalizations of adult cats can be divided in three major classes (Moelk, 1944; Bowen et al., 2005; Rodan, 2010):
1. Sounds made with the mouth closed: murmurs including the purr and the trill. The purr, produced by a continuous vibration during both inhalation and exhalation, and the trill are friendly greetings. The purr is also a care-soliciting call that typically occurs during amicable social interactions or on the contrary when ill or injured.
2. Sounds made while the mouth is kept opened and then gradually closed. These include a vast range of meows that are made in diverse situations - demands, begging demands, bewilderment, complaint, mild forms of the mating cry and the anger wail, to interact with other cats, dogs and humans.
3. Sounds produced while the mouth is tensely opened in one position: including growl, yowl, snarl, hiss, spit and shriek which are usually related to aggression.
McComb and colleagues (2009) found that when humans were played purrs recorded while cats were asking their owner for food at equal amplitude to purrs recorded in non-solicitation contexts, even individuals with no experience of owning cats judged the ‘solicitation’ purrs to be more urgent and less pleasant.
A study by Schötz (2014) where 36 human listeners classified 28 cat vocalisations into seven emotion categories (“Joy”, “SorrowFear”, “Anger, “Desire”, “Neutral”, “Other” and “Don’t know”) demonstrates that human listeners are not very accurate judging the emotional state of cat vocalizations. Their results also agree with McComb and colleagues (2009) study in the proposition that cats are able to adapt their vocalizations to human listeners, varying their intonation to signal different emotions, and humans perceive them based in their own cues to signal emotions in human speech.
1.5.3– Olfactory Communication
The communication within cats is achieved by a whole variety of olfactory signals and every single one of these signals demands sniffing by the communication recipient (Brown et al., 2014).
It seems to be a widely spread truth that smell consists of an important part of the cat’s sensory input, based on the size of their olfactory bulbs, olfactory epithelium, the complexity of their olfactory receptors and the glands present throughout their body, which show their high adaptation to send and receive scent signals (Bowen et al., 2005; Brown et al., 2014).
Such a complex olfactory apparatus seems to obviously support the ability to smell and track a prey (Brown et al., 2014). It also serves as a means of self-communication, as a scent can be deposited in the environment to signal familiarity and security for itself. Other use of the scent is the avoidance conflicts with unknown conspecifics while hunting, a process cats prefer to do alone, and to protect their hunting area. Cats also use scent to mark other kinds of territory, such as sleeping places and play areas. By using a communication mode that signals from a distance its occupancy of territory, cats prevent direct physical interaction with strangers. Also, the scent that the individual produces is believed to be unique, thereby being used to identify it to other cats of the colony and to unknown individuals. (Pageat, 1998; Bowen et al., 2005).
The substances used in olfactory communication are produced by a wide set of sebaceous glands located all along the cat’s body, especially their face, perianal area and in between the digits (Crowell-Davis et al., 2004; Bowen et al., 2005). These secretions include, among the various odour molecules, molecules that transmit highly specific information between conspecifics, called Pheromones. This term was developed by Karlson, Luscher and Butenand, in 1959, which combined the Greek verbs pherein (to carry) and horman (to stimulate) (Pageat et al., 2003).
Olfactory communication makes use of urine, faeces and scratching and rubbing behaviours which also materialize in visual stimulus (Pageat, 1998; Crowell-Davis et al., 2004).
1.5.3.1– Pheromones
Although they might have olfactory characteristics and there are structures present in olfactory mucosa, which serve for pheromone detection, pheromones are not simple odours (Pageat, 1997; Pageat et al., 2003). Their perception is also different from the perception of the rest of the odours, which is a spontaneous phenomenon (Pageat et al., 2003).
In invertebrates these secretions are monomolecular or composed by a small number of molecules. In mammals these secretions are much more complex. The identified components are organic compounds of moderate molecular weight comprehending the following chemical classes: carboxylic acids, alcohols ketones, aldehydes, amines, sterols, terpenes and alkanes. Some amine secretions, such as amine anal secretions (putrescin, cadaverine and trimethylamine associated with indol), can be very unpleasant but, in general, these secretions are too discreet for the human olfactory system (Pageat, 1997).
Depending on the molecular weight of the components, the volatility of the molecules can vary and so can the range in which they can still act in the individuals. Therefore two types of pheromones can be found (Pageat, 1997):
Near-acting pheromones: only detected in a small distance and might be associated with emission or deposition modes which highlight their presence (vocalizations, movements like leaps and to-and-fro or urine traces);
Remote-acting pheromones: pheromones that can act at great distance and can be associated with visible signs like scratching.
The transport of pheromones by other molecules is a process still to confirm in carnivores. The carrier molecules are complex protein structures which form weak electrical bonds thus enabling reversible attachment of different organic molecules (especially carboxylic acids). These protein are very volatile, have very marked adhesive properties and have a number of structural analogies with the Fel d 1 protein of the cat, an well-known allergen which can firmly adhere to any support (Pageat, 1997; Pageat et al., 2010). The Fel d 1 protein has a high immunological polymorphism among the cat population but its exact biological function and role on communication is still unknown (Pageat et al., 2010). It has been proved that this protein is produced in
the sebaceous cells and squamous epithelial cells of the skin spreading to the surface of epidermis and fur, anal sacs, and to a lesser extent in the salivary glands (Charpin et al., 1991; Mata et al., 1993; De Andrade et al., 1996).
Cats are abundantly provided with pheromone-producing structures which include certain glands and mucosa (Pageat, 1997; Pageat et al., 2003):
Anal sacs or anal glands: a set of functionally and chemically secretions produced in this area are known but the tissues that produce them are not located precisely. These glands are well-known by veterinarians because inflammations can occur in particularly in tomcats. Although they are related to the identification of the male cat by the female in estrus, their exact function is not totally known. The perianal hepatoid glands, the sebaceous glands of the skin portion of the anus and probably the rectal mucosa must also be mentioned;
Subcaudal glands: secreting cells diffused in the ventral surface of the tail’s root of cats;
Supracaudal glands/Hepatoid: sebaceous glands of elliptical shape located on the dorsal surface of the tail’s root. Its activity seems to depend on secretion of sexual steroids. When cats are castrated, reductions in volume of its secretions are clearly observed;
Jugal and perioral glands: these comprise a set of secreting structures spread through the chin, lips, skin region of the snout’s vibrissae and cheeks. Cats deposit the secretions of these glands in objects, conspecifics or humans by rubbing the lateral portion of the face, starting from the corner of the lips to the front region of the ear.
Podal glands: diffuse glands present in the foot pads and in the interdigital skin.
The perception of pheromones is not yet completely understood. Despite this, the method comprising the Jacobson’s organ or the vomeronasal organ (VNO) and the Flehmen behaviour are the best known cases (Pageat, 1997; Pageat et al., 2003).
The vomeronasal or Jacobson’s organ seems to have specialised only for the purpose of detecting and processing social scents, in the cat. The paired vomeronasal organs are situated under the anterior end of the nasal cavity and are linked to both nasal and oral passages through the nasopalatine canal (Liman, 1996; Pageat et al., 2003;
Brown et al., 2014). The lumen of the VNO is covered with two kinds of ephitelium: the medial one that includes the nervous cells whose axons merge to create the vomeronasal nerve, and the lateral one, which consists of a respiratory mucous membranes (Pageat et al., 2003).
The Flehman behaviour exists in mammals such as felines, canines, equines, bovines, ovines and porcines. Although its role has been associated to detection of sexual pheromones, this behaviour has been observed in castrated individuals and occurring in situations not related to sexual activity. When a cat faces a new scent which draws its attention and will to inspect, it is prone to initially sniff it. This behaviour may be followed by the Flehman behaviour, which can be seen as a raising of the upper lip with mouth slightly opened. Cats also move their tongue to complete the behaviour. The lip rising muscles are pulled upwards and on both sides, on the cartilaginous valve which partially covers the incisive meatus so this opens its lumen and a small reduction on the blood pressure in the lumen of the Jacobson’s organ occurs. This causes a collapse of the erectile vascular body of the vomeronasal organ which unblocks access to the olfactory part of the canal and leads to the suction of dissolved substances into the mucus. These substances contact with olfactory cells from the mucosa of the Jacobson’s organ and the stimulus initiates pulses that go through the vomeronasal nerve into the accessory olfactory bulbus and from there, fibbers go through the olfactory tract to the corticomedial structure of the amygdaloid body. This differs from the other olfactory neurons that emit extensions towards the olfactory tubercles- hypothalamic region, but not the amygdaloid body – limbic system (Pageat, 1997; Pageat et al., 2003).
There is still doubts about the possibility that some pheromones can be detected by gustatory cells as secretions such as the sexual ones are also explored through this route (Pageat, 1997). Actually, when it comes to sexual pheromones, cats normally make direct nasooral contact with the stimulus source before performing the Flehmen behaviour, indicating that the Flehman also serves to transport fluid-borne signals from the oral cavity to the VNO (Hart et al., 1987).
Work from Hart and Leedy (1987) showed that the Flehmen in cats in the context of sexual encounters between the two genders is highly sexually dimorphic as it is only performed by males. However, when the males were not present, females performed the Flehmen if the stimulus was presented to them as urine marking (even though less than males in the absence of females) and when the stimulus was applied by the authors on the nose of the cats, the dimorphism disappeared. These results suggest that there is a
sexual predisposition of males and females to explore and contact with the stimulus material, rather than a difference in the probability in responding to the presence of urine stimuli. Hart and Leedy (1987) also treated spayed females with testosterone and verified that this increased their performance of genital inspection and display of Flehmen thus proving that in addition to being stimulus dependent, this behaviour is also under hormonal control.
It is clear that the Flehmen behaviour is mediated by a different neuronal system than the one which governs male sexual behaviour as bilateral lesions performed in the medial preoptic-anterior hypothalamic (MP-AH) made in prepubertal male cats, impaired their adult copulatory behaviour but spared their Flehman behaviour (Leedy et al., 1986).
The fact that the fibbers carry the impulse from the accessory bulbs to the amygdaloid body of the limbic system suggests two major actions of the pheromones: intervene on hormonal secretions via the hypothalamus (sexual steroids are the most notable) and induce emotional modifications causing variations of reactional status, this last causing orientation of behaviour such as avoiding reaction, running away, inclination to inhibition or submission or even responding aggressively (Pageat, 1997).
The analysis sebaceous secretion isolated from the cheeks of cats has detected 40 different chemical components but only 13 of these are common to all cats. Between these common elements, five associations corresponding to “different messages” were named from F1 to F5 (Pageat, 1997; Pageat, 1998; Pageat et al., 2003).
The F2 secretion has been associated depositions by facial marking during sexual excitation and to activation of urine marking (Pageat, 1997).
The F3 is deposited on familiar objects, having an emotional stabilization and special orientation effect thus lowering the probability of emergence responses due to fright or fear, promotes feeding and exploratory behaviours and inhibits urine marking (Pageat, 1997; Gaultier et al., 1998; Pageat, 1998; Frank et al., 1999; Griffith et al., 2000; Pageat et al., 2003).
The F4 seems to be the chemical support of allomarking, being deposited on cats within the same colony and also on human beings or other animals with whom the cat has a social relationship inhibiting aggressive behaviour (Pageat, 1997; Pageat et al., 1997; Pageat et al., 1997; Pageat et al., 2003).
These pheromones seem to play an important role on geographic orientation, being mostly deposited on objects which consist of boundaries between a passage and a territorial zone (Pageat, 1998; Pageat et al., 2003). The role of these pheromones in managing space is completed by those of the podal glands deposited by scratching and by the ones which have an alarm function which are conveyed by the secretions from anal sacs and from podal glands (Pageat, 1997).
For cats and dogs, pheromone investigations have been focused in the ones comprising the sexual and the territorial behaviour. For cats there has been special interest in urine marking (Pageat, 1997).
1.5.3.2 – Urine marking
This kind of marking is a very well-known behaviour in the cat being considered as a problem for most owners (Pageat et al., 2003). Urine marking, also called spraying, has a typical behavioural posture performed by the cat, where it is standing on its four limbs, with the tail up quivering, slightly arches its back and projects a small volume of urine on a vertical object. Mydriatic pupils and a vacant expression on the face of the cat while performing the marking can be seen. Despite this most classical sequence, some cats urine mark in a squatting position. Females and males can urine mark but males tend to perform this behaviour more frequently. Castration in prepuberal stages reduces the probability of triggering urine marking (Dehasse, 1997; Pageat, 1997; Pageat et al., 2003; Bowen et al., 2005).
Image 5 - Typical posture adopted when urine marking (adapted from Beaver, 2003)
There is a large number of contexts within which urine marking occurs, and it sure has an even larger variety of functions such as communicating specific identifying information, e.g. ‘intact female/male ready for mating’, communicating location information and elaborating a time-share system of the territory thus avoiding contact with unfamiliar individuals, e.g. ‘I entered this field X hours ago’, communicating emotional information, e.g. ‘I am here and I am highly aroused (Crowell-Davis et al., 2004; Bowen et al., 2005). Within the domestic environment, urine marks can be self-directed signalling a territory as its own (a reassuring message) or even marking something unpleasant and signalling a particular location as a potential danger (Bowen et al., 2005). Some cats use spraying for seek attention from their owners or redirect activity when other forms of interaction are frustrated (Bowen et al., 2005; Rodan, 2010). Urine spraying can also occur in situations of extreme excitement like during the hunting (as some cats may spray on rabbit burrows) or when a stranger invades its privacy area (Dehasse, 1997). Both sexual and reactional marking are the most common kinds of urine marking. Sexual marking, often comes with vocalization, is strongly related to sexual arousal on male cats and as to the female, the marking is related to the beginning of estrus. Reactional urine marking is prone to occur due to changes in the cat’s environment: new furniture, new house or new occupant (Pageat et al., 2003).