Summary To evaluate differences in growth and adaptabil-ity of maritime pine (Pinus pinaster Ait.), we studied growth, polycyclism, needle tissue carbon isotope composition (δ13C) as an estimate of water-use efficiency (WUE) and survival of seven populations at 10 years of age growing in a performance trial at a provenance test site in Escaroupim, Portugal. Six pop-ulations were from relatively high rainfall sites in Portugal and southwestern France (Atlantic group), and one population was from a more arid Mediterranean site in Spain. There were sig-nificant differences between some populations in total height, diameter at breast height, δ13C of bulk needle tissue, poly-cyclism and survival. A population from central Portugal (Leiria, on the Atlantic coast) was the tallest and had the lowest δ13C. Overall, the variation inδ13C was better explained by the mean minimum temperatures of the coldest month than by an-nual precipitation at the place of origin. Analyses of the rela-tionships betweenδ13C and growth or survival revealed a dis-tinct pattern for the Mediterranean population, with lowδ13C (and WUE) associated with the lowest growth potential and re-duced survival. There were significant negative correlations betweenδ13C and height or survival in the Atlantic group. Vari-ation in polycyclism was correlated with annual precipitVari-ation at the place of origin. Some Atlantic populations maintained a high growth potential while experiencing moderate water stress. A detailed knowledge of the relationships between growth, survival andδ13C in contrasting environments will en-hance our ability to select populations for forestry or conserva-tion.
Keywords: climate change, drought tolerance, maritime pine, polycyclism, provenance trial, tree breeding.
Introduction
Pinus pinaster Ait. (maritime pine), a Mediterranean forest tree species that covers large areas and occurs in a wide variety of habitats, has great economic and ecological importance. The climates prevailing in the areas occupied by P. pinaster are characterized by summer droughts. Because water stress is
one of the main factors limiting growth of this species and be-cause predicted climatic scenarios point to an increasing se-verity of summer droughts in areas with a Mediterranean cli-mate, including the Atlantic coast of the Iberian Peninsula (Miranda et al. 2006), large areas of P. pinaster’s current range may become more xeric. Therefore, detecting differences in drought tolerance and growth among populations may allow selection of populations adapted to drought-prone environ-ments.
Genetic differences in growth and survival of P. pinaster populations in provenance tests have been evaluated in several studies (Guyon and Kremer 1982, Kremer and Roussel 1986, Danjon 1994, Alía et al. 1995). These studies show wide dif-ferentiation in growth and survival traits across the natural range of the species, and smaller differences at a regional scale (Harfouche et al. 1995). Differential adaptation to regional and local patterns of climate and soil type plays an important role in determining the quantitative trait structure of the spe-cies (González-Martínez et al. 2002). Overall, drought-adapted provenances display lower vigor than mesic Atlantic prove-nances under favorable conditions, but are better able to sur-vive severe summer droughts (Alía et al. 1995, 1997).
Polycyclism, the production of multiple shoot flushes in a single season, is a highly heritable characteristic, expressing adaptation to intermittent water availability and showing con-siderable variation among provenances (Kremer and Roussel 1986, Danjon 1994) or families (Kremer 1981). Annual shoot development in maritime pine depends only on the expansion of buds containing the primordia of the shoots of the following year. Mono or polycyclic bud formation is highly sensitive to temperature and summer water deficits, and variation in the number of stem units predetermined in the bud explains most of the variation in annual height increment in maritime pine (Kremer and Roussel 1986). The association of polycyclism with growth or precipitation at the place of origin has been in-vestigated in several studies. Although no clear relationship between polycyclism and precipitation at the site of origin (Alía et al. 1997) or with annual height growth (Danjon 1994) was detected in maritime pine at the provenance level, a posi-© 2008 Heron Publishing—Victoria, Canada
Variations in growth, survival and carbon isotope composition (
13
C)
among Pinus pinaster populations of different geographic origins
ISABEL CORREIA,
1,2MARIA HELENA ALMEIDA,
1ALEXANDRE AGUIAR,
3RICARDO
ALÍA,
4TERESA SOARES DAVID
3and JOÃO SANTOS PEREIRA
11
Instituto Superior de Agronomia (Dep. Eng. Florestal), Tapada da Ajuda, 1349-017 Lisboa, Portugal
2
Corresponding author ([email protected])
3
Instituto Nacional dos Recursos Biológicos, I.P., Quinta do Marquês, 2780-159 Oeiras, Portugal
4
Instituto Nacional de Investigación y Tecnologia Agraria y Alimentaria, Carretera de la Coruña Km 7.5, 28040 Madrid, Spain
tive association with early height growth at the individual level was reported by Kremer (1981), suggesting that further inves-tigation of polycyclism is needed.
Water-use efficiency (WUE; leaf photosynthetic rate/tran-spiration rate) and drought tolerance of plants may be assessed by carbon isotope discrimination during photosynthesis. Fo-liar discrimination for13C in C3plants is negatively related to carbon isotope composition (δ13C) and WUE representing an integrated measure of this parameter during carbon fixation (Farquhar et al. 1989, Brugnoli and Farquahr 2000, Máguas and Griffiths 2003). Genetic variations in carbon isotope dis-crimination have been found in many species, including pine populations (Cregg and Zhang 2001, Guy and Holowachuk 2001), families (Prasolova et al. 2000, 2001, Xu et al. 2003) and clones (Prasolova et al. 2003, 2005). A large diversity in carbon isotope discrimination was also found in P. pinaster at the provenance (Guehl et al. 1995, Nguyen-Queyrens et al. 1998) and family levels (Brendel et al. 2002). These early studies demonstrated clear adaptation to conditions at the place of origin, with ecotypes from arid environments display-ing a lower WUE than ecotypes from mesic environments.
Carbon isotope composition may vary either directly or in-versely with plant growth depending on the environmental conditions under which comparisons are made. Such variation must be better understood before the carbon isotope technique can be applied in selection programs (Condon et al. 2004). Re-lationships betweenδ13C and growth have been investigated in many forest tree species. Although Pinus ponderosa Dougl. ex Laws. populations differed in growth but not inδ13C, showing no relationship between drought tolerance and WUE (Zhang et al. 1997), there was a strong correlation between height growth andδ13C in Picea mariana (Mill.) B.S.P. genotypes (Johnsen et al. 1999) and weaker phenotypic correlations were found betweenδ13C and growth in Pinus pinaster (Nguyen-Queyrens et al. 1998). Quantitative trait loci were detected for growth (Plomion et al. 1996) and for δ13C (Brendel et al. 2002) in maritime pine, indicating a strong genetic control over these traits that may assist in selecting genetic material with improved growth for afforestation in drought-prone envi-ronments.
We hypothesized that WUE varies among populations from different origins and is related to survival, growth or climatic conditions of a population’s origin. To determine if WUE can be used as a criterion for selection of drought-tolerant genetic material, our specific objectives were: (1) to characterize pop-ulation diversity for growth, survival and polycyclism; (2) to characterize population diversity for WUE, usingδ13C of nee-dle tissue as a proxy; (3) to evaluate relationships between WUE and growth, survival and polycyclism; and (4) to assess relationships between the analyzed traits and climatic condi-tions of each population’s origin. To our knowledge, this is the first study to compare mostly Atlantic populations in the field in a subhumid environment. The seven populations studied ranged in origin from humid to semiarid climates. Six of the populations (from Portugal and Landes, France) belong to the Atlantic group, according to the three P. pinaster geographic races (Atlantic, Mediterranean and Mahgrebian) defined by
Baradat and Marpeau-Bézard (1988). The Cuenca population belongs to the Mediterranean group, its origin being in eastern Spain, which has a more arid climate than the Atlantic habitat. Measurements were made under field conditions in 10-year-old trees growing in a provenance trial at a subhumid site in Escaroupim, Portugal.
Materials and methods
Plant material and experimental design
Our study was conducted at one of five test sites of a maritime pine (Pinus pinaster) provenance trial established in Portugal in 1993. The site selected for our study was Mata Nacional do Escaroupim, in central Portugal (39°04′ N, 8°44′ W). The trial comprises a randomized complete block design with 2–8 blocks in each test site, each of 30 populations being repre-sented in each randomized block by 30 unrelated trees, spaced 2 × 3 m apart in 30-tree square plots. For our study, seven pop-ulations of the 10-year-old trees growing in two blocks in Escaroupim were selected. Six populations were from the At-lantic coast and one from the Mediterranean (Figure 1). Four Atlantic populations originating in Portugal were from stands selected for seed production along a climate gradient (Table 1). The remaining Portuguese population was from open-polli-nated crosses of phenotypically selected trees (plus trees) of the Leiria population. Locations, altitudes and climate data for the source locations of the Pinus pinaster trial populations are provided in Table 1. Escaroupim, the trial site, has a subhumid climate according to the Emberger rain-temperature coeffi-cient (Q3), with a dry season from May to September. How-ever, the proximity of a River Tagus tributary creates a high phreatic surface, representing a source of water that may atten-uate water deficits during the dry season. Escaroupim is lo-cated at 10 m a.s.l., and soil comprises Dystric Regosols (RG dy) deriving from quartz sand deposits from fluvial ori-gin.
Figure 1. Origins of Pinus pinaster populations analyzed for carbon isotope composition in the trial at Escaroupim.
Three climatic variables were used to test correlations be-tween carbon isotope composition and growth, polycyclism, survival and climatic conditions of each population’s origin: annual cumulative precipitation (P; mm); mean minimal tem-perature for the coldest month (m; °C); and Q3 estimated as: Q3 = 3.43 P/(M – m), where M is the mean maximal tempera-ture for the hottest month (°C) (Stewart 1968).
Sampling procedures and field measurements
Eight to 10 trees from each of the seven populations were ran-domly chosen in each of two contrasting randomized blocks. Healthy needles of the selected trees were collected in the spring of 2003 from the west-facing middle part of the crown. All needles were equally sun exposed and growing under simi-lar irradiances. From each tree, six pairs of needles that emerged the previous spring were dried for 5 days at 65 °C and milled to a fine powder. Total height (± 0.10 m) of the trees sampled for13C analysis was determined with a hypsometer (Vertex) and the mean of three measurements is presented. Di-ameter at breast height (DBH; ± 0.5 cm) was determined with a calliper, and polycyclism was inferred from the number of whorls. Survival in the study plots was evaluated as the per-centage of surviving trees of the 30 initially planted in each plot.
Carbon isotope composition
Subsamples of about 1 mg of bulk needle tissue were assayed for δ13C with an isotope ratio mass spectrometer (Mylne-field Research Services, Scotland). Carbon isotope composi-tion (‰) in the needle samples was calculated according to Farquhar et al. (1989): δ13 13 12 Sample 13 12 VPDB C C C C C = ⎛ ⎝ ⎜ ⎞ ⎠ ⎟ ⎛ ⎝ ⎜ ⎞ ⎠ ⎟ − ⎛ ⎝ ⎜ ⎜ ⎜ ⎜⎜ ⎞ ⎠ 1 ⎟ ⎟ ⎟ ⎟⎟ 103
where VPDB refers to the standard Vienna Pee Dee Belem-nite. Theδ13C values in C3plants are directly proportional to WUE during carbon fixation (Jones 1992).
Data analysis
Covariance parameters were estimated by a linear mixed model based on restricted maximum likelihood (Reml) meth-odology. The selected model considered population and block as random effect factors and estimated three variance compo-nents—one for populations, one for blocks and one for resid-ual errors. Alternative models considering a variance for each population, or a tree nested within population effect, estimated null variances and proved to be inadequate in explaining the variation observed.
Best linear unbiased prediction (Blup) values were esti-mated for each population by univariate analyses, and differ-ences between all possible population pairs were evaluated by contrasts. The selected model can be expressed as:
y = Xβ + Zγ + ε
where y = vector of observations; X = design matrix of the in-terception (column of ‘1’);β = unknown vector of the fixed ef-fect parameters (grand mean) with known matrix X; Z = de-sign matrix of the parameters associated to populations and blocks;γ = unknown vector of the random effect parameters
GROWTH, SURVIVAL ANDδ C IN MARITIME PINE 1547
Table 1. Locations, altitudes and climate data for the source locations of the Pinus pinaster trial populations and the Escaroupim trial site. The spe-cific place of seed collection and the seed collection code, if any, are given in parenthesis after each population name. Climatic data represent 30-year means. Abbreviations: P = annual cumulative precipitation; M = mean maximal temperature for the hottest month; and m = mean minimal temperature for the coldest month. Emberger rain-temperature coefficient: Q3 = 3.43 P/(M – m) (Stewart 1968).
Population Altitude (m) Latitude (N) Longitude (W) P (mm) M (°C) m (°C) Q3
Landes 40 – – 984.2 26.6 2.8 141.8 Humid
(Lavercantière)
Bragança 800 41°52′ 6°32′ 742.7 28.3 0.5 91.6 Subhumid
(Albagueiras, Pb42)
Alcácer do Sal 20 37°52′ 8°30′ 572.8 30.8 4.9 75.9 Subhumid
(M. de Moita, Pas54) Escaroupim 10 39°04′ 8°44′ 577.6 28.4 4.5 82.9 Subhumid (M.N. Escaroupim, Pcse0102) Cuenca 800 39°58′ 1°38′ 507 30.7 –0.7 55.4 Semiarid (Boniches) Monção 500 42°04′ 8°24′ 1189.7 28.4 4.5 170.7 Hyperhumid (Anta T23, Pm36) Leiria 55 39°46′ 8°58′ 678 21.5 6.5 155.0 Humid (T164, Pmg52)
(populations and blocks) with known matrix Z; andε = un-known vector of residual errors. Values ofε and γ are normally and independently distributed.
Pearson’s correlation coefficients between all data values for all traits and climatic variables were calculated. General patterns and associations between traits and populations were analyzed by principal component analysis based on Blup mean population estimates and represented by Genotype × Trait biplots (Yan 2002).
Results
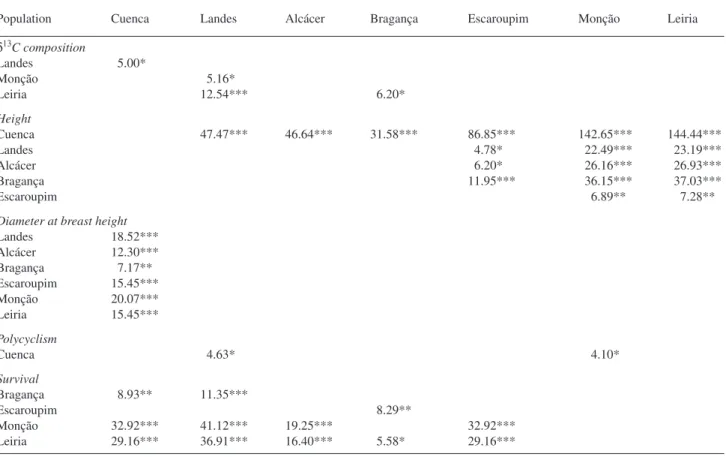
The Landes population had the highestδ13C values and the Leiria population had the lowestδ13C values (Table 2). Among populations, Landes showed the highest polycyclism and the lowest survival rate, and Leiria displayed superior growth and survival. Cuenca, with a relatively lowδ13C value and the low-est growth rate and polycyclism score, differed from the gen-eral population trend of lower δ13C associated with higher growth. Large within-population variability in the measured characteristics was found (coefficients of variation ranging from –3.98 in Leiria to –2.91 in Bragança), as previously re-ported for this species (Guehl et al. 1995, Nguyen-Queyrens et al. 1998, Brendel et al. 2002). Nevertheless, significant population effects (α = 0.05) were detected for δ13C (F = 2.4, Pr= 0.0311), total height (F = 34.32, Pr< 0.0001), DBH (F = 5.3, Pr< 0.0001) and survival (subjected to angular transfor-mation: F = 15.63, Pr< 0.0001) but not for polycyclism (F = 1.39, Pr= 0.2232). The Reml estimates for the variance com-ponents showed that population variance represented 8% of the total variance forδ13C, 7% for polycyclism and between 21 and 63% for the other characteristics (data not shown).
There were significant differences in height (α = 0.05) be-tween most of the populations, except for Landes versus Alcácer and Bragança, Alcácer versus Bragança, and Monção versus Leiria. Cuenca differed significantly from all other pop-ulations in DBH. Differences in survival were observed for Cuenca and Landes comparisons versus Bragança, Monção and Leiria and between most Portuguese populations. Signifi-cant differences inδ13C, matching differences in total height, were found for some of the comparisons of the Landes popula-tion (versus Cuenca, Monção and Leiria) and for Leiria versus
Bragança (Table 3). Few population differences in poly-cyclism were detected (Cuenca versus Landes and Monção). Results are in agreement with the study by Kremer and Roussel (1986) who found significant differences between provenances, the most polycyclic being Landes and the least polycyclic being a provenance from the center of the Iberian Peninsula, certainly from the Mediterranean group.
Estimates of Pearson’s correlation coefficients between all data values in the Atlantic group (i.e., excluding Cuenca) re-vealed significant negative correlations (α = 0.05) between δ13
C and total height, survival and m (Table 4). Total height was positively correlated with DBH, P, m and Q3. Survival was positively correlated with P and, most closely of all, with Q3. At the population level, no correlation was found between polycyclism and growth (data not shown); however, at the in-dividual level, a positive association between polcyclism and DBH was detected. Polycyclism was also positively correlated with P of the populations’ origin.
Multivariate analysis explained 82.1% of the observed vari-ation (Figure 2) and the biplot representvari-ation revealed a posi-tive association betweenδ13C and polycyclism and between growth and survival (although not significant based on Pearson correlation coefficients). The biplot representation shows that Landes had the highestδ13C and polycyclism values, whereas Leiria had high growth and survival.
Discussion
Growth, polycyclism and survival
Significant differences in survival and total height were de-tected for most pairwise comparisons of the P. pinaster popu-lations growing in Escaroupim. Compared with trees originat-ing from sites with low P and cold winters, trees originatoriginat-ing from sites with higher P and warmer winters displayed signifi-cantly better growth in Escaroupim, reflecting the adaptive po-tential of this species. In contrast, we found only small differ-ences among populations for the other analyzed traits. Ribeiro et al. (2001) reported low differentiation among Portuguese populations and high differentiation within populations based on nuclear and cytoplasmic markers. The higher differentia-tion in survival and total height detected in our study than in
Table 2. Carbon isotope composition (δ13C), total height, diameter at breast height (DBH), polycyclism of the sampled trees and survival rate on a plot basis: best linear unbiased prediction values (± confidence limits) for each population forα = 0.05.
Population δ13C (‰) Total height (m) DBH (cm) Polycyclism Survival rate (no. whorls) Landes –26.72 ± 0.80 6.97 ± 0.62 13.57 ± 1.83 19.59 ± 1.46 0.58 ± 0.09 Bragança –26.98 ± 0.80 6.55 ± 0.62 11.91 ± 1.83 19.18 ± 1.46 0.75 ± 0.09 Alcácer do Sal –27.14 ± 0.80 6.89 ± 0.60 12.68 ± 1.75 18.83 ± 1.42 0.67 ± 0.09 Escaroupim –27.16 ± 0.80 7.70 ± 0.60 13.10 ± 1.75 18.33 ± 1.42 0.61 ± 0.09 Cuenca –27.25 ± 0.80 4.67 ± 0.60 9.18 ± 1.75 18.12 ± 1.42 0.61 ± 0.09 Monção –27.26 ± 0.80 8.56 ± 0.60 13.65 ± 1.75 19.48 ± 1.42 0.87 ± 0.09 Leiria –27.56 ± 0.80 8.58 ± 0.60 13.82 ± 1.75 18.60 ± 1.42 0.86 ± 0.09 Mean –27.15 ± 0.77 7.13 ± 1.30 12.56 ± 1.80 18.88 ± 1.16 0.70 ± 0.13
the study by Ribeiro et al. (2001) may be associated with the different sampling strategies and the poor correlation between neutral molecular markers and quantitative variation (Rolf et al. 2006).
The positive association of polycyclism with DBH, but not with total height, may result from a high incidence of diseased trees in the trial plots, especially from processionary caterpil-lar Thaumetopea pytiocampa Schiff. which causes main stem damage and hence leads to underestimation of total height. In contrast to Kremer and Roussel (1986), who found no connec-tion between polycyclism and the place of origin, our data
sug-gest that trees originating from sites with high annual rainfall tended to be more polycyclic when growing in Escaroupim than trees originating from sites with low rainfall (Table 4). This increase in polycyclism could result from an ability of the trees to exploit a longer growth season favored by higher tem-peratures and abundant water in Escaroupim, where a high wa-ter table contributes to minimize the effects of summer drought. The results also suggest that differences in poly-cyclism between populations could be more closely related to short-term climatic variation or small-scale spatial variation than to regional climate differences among the sites of origin.
GROWTH, SURVIVAL ANDδ C IN MARITIME PINE 1549
Table 4. Pearson’s correlation coefficients (and significance levels), between all data values in the Atlantic group of populations (Cuenca not in-cluded), for all analyzed traits and some climatic variables at the population origin. Abbreviations:δ13C = carbon isotope composition; DBH =
di-ameter at breast height; P = annual cumulative precipitation; m = mean minimal temperature for the coldest month; and Q3 = Emberger rain-temperature coefficient. Significant correlations are in bold.
δ13
C Height DBH Polycyclism Survival
Height –0.22 (0.019) DBH –0.05 (0.624) 0.70 (< 0.0001) Polycyclism 0.03 (0.729) 0.15 (0.102) 0.36 (< 0.0001) Survival –0.25 (0.006) 0.13 (0.173) –0.11 (0.220) 0.06 (0.536) P 0.07 (0.454) 0.20 (0.034) 0.09 (0.329) 0.23 (0.014) 0.26 (0.005) m –0.26 (0.004) 0.45 (< 0.0001) 0.15 (0.105) –0.14 (0.147) 0.18 (0.055) Q3 –0.105 (0.058) 0.44 (< 0.0001) 0.17 (0.072) 0.13 (0.153) 0.41 (< 0.0001)
Table 3. Summary of F statistics of contrasts of populations pairs for carbon isotope composition (δ13C), total height, diameter at breast height, polycyclism and survival (subjected to angular transformation). Only pairs showing differences with Pr< 0.05 are represented. Significance is
in-dicated by asterisks: *, Pr< 0.05; **, Pr< 0.01; and ***, Pr< 0.001.
Population Cuenca Landes Alcácer Bragança Escaroupim Monção Leiria δ C composition Landes 5.00* Monção 5.16* Leiria 12.54*** 6.20* Height Cuenca 47.47*** 46.64*** 31.58*** 86.85*** 142.65*** 144.44*** Landes 4.78* 22.49*** 23.19*** Alcácer 6.20* 26.16*** 26.93*** Bragança 11.95*** 36.15*** 37.03*** Escaroupim 6.89** 7.28**
Diameter at breast height
Landes 18.52*** Alcácer 12.30*** Bragança 7.17** Escaroupim 15.45*** Monção 20.07*** Leiria 15.45*** Polycyclism Cuenca 4.63* 4.10* Survival Bragança 8.93** 11.35*** Escaroupim 8.29** Monção 32.92*** 41.12*** 19.25*** 32.92*** Leiria 29.16*** 36.91*** 16.40*** 5.58* 29.16***
Carbon isotope composition
The range of variation inδ13C among populations (–27.56 to –26.72‰) was smaller than that found by Guehl et al. (1995) who compared provenances from the three geographic races of P. pinaster: Atlantic, Mediterranean and Mahgrebian. The Landes population, displaying the highestδ13C (and WUE), was clearly distinguishable from Mediterranean Cuenca and also from the Atlantic coastal populations Leiria and Monção, which had the lowestδ13C of the populations studied. Signifi-cant differences inδ13C values between Landes and other pop-ulations may be explained by the adaptive evolution of Landes which originates in a northern environment with less severe summer droughts, cold winters and shorter growing seasons. Landes likely initiated stomatal closure at less severe water deficits than the other populations studied, leading to lower discrimination (highestδ13C).
Differences inδ13C values among the Portuguese popula-tions were small, nevertheless there was a significant differ-ence between Bragança and Leiria. This differdiffer-ence may reflect divergent climatic conditions at the locations of origin: annual rainfall is similar at both origins, but Bragança originates in an environment with lower winter temperatures and greater sea-sonal temperature variation than Leiria, which is adapted to climatic conditions similar to those at Escaroupim. Therefore, Leiria could exploit a longer period of needle formation and stomatal aperture in Escaroupim, leading to lower δ13C, whereas Bragança, adapted to colder winters and hotter sum-mers, could be under a genetic control inducing early stomatal closure. Differences between Bragança and Leiria may also be related to the relatively well preserved identity of the Bragança population because of its geographic isolation, which likely reduced natural or human-induced gene flow from other
popu-lations. The distinct performance of Bragança has been docu-mented by Bahrman et al. (1992) using biochemical markers.
Relationships betweenδ13C, growth and climate
Landes and Bragança, originating from environments with colder winters than in Escaroupim (m = 4.5 °C), displayed higherδ13C and slower growth at this site than the other popu-lations. Leiria and Monção, originating from regions with milder temperatures than in Escaroupim, presented the lowest δ13
C and higher growth than the other populations. However, the few differences inδ13C among populations in this trial were insufficient to explain the observed growth differences. Cuenca followed a distinct pattern, with lowδ13C (and WUE) associated with the lowest growth potential of the populations studied, which may be explained by a different adaptive strat-egy of this population, the only one belonging to the Mediter-ranean group. Cuenca’s MediterMediter-ranean climate (affected by the continental influence of the Iberian landmass) is character-ized by high daily temperature amplitudes throughout the year, especially in the hottest months. Guehl et al. (1995) re-ported that the drought-adapted Moroccan populations have higher carbon isotope discrimination (i.e., lower δ13C and WUE) than the other Mediterranean and Atlantic races. Nguyen-Queyrens et al. (1998) also found lowerδ13C for the drought-tolerant Tamjoute (Mahgrebian) provenances than for mesic Landes provenances (Atlantic). Drought-adapted ecotypes favor carbon assimilation under well-watered condi-tions, leading to higher discrimination against13C, and rely on higher root to shoot biomass ratios (Guyon and Kremer 1982). Drought adaptation in tree populations is not necessarily re-flected in higher WUE, as verified in other drought-tolerant species, and a lower WUE may be related to adaptive mecha-nisms under genetic control (DeLucia and Schlesinger 1991, Read and Farquhar 1991, Hubick and Gibson 1993).
As discussed by Tognetti et al. (2000), mesic ecotypes of P. pinaster rely mainly on stomatal control to reduce transpira-tion during mild water stress. Tognetti et al. (2000) compared four Mediterranean and one Atlantic provenance (Leiria) in four subhumid Mediterranean sites and used carbon isotope discrimination as a physiological index of drought adaptation to rank xeric and mesic populations, with Leiria ranking as the most mesic. A strong negative correlation was found between carbon isotope discrimination and growth at the coldest site but the authors hypothesized that an opposite relationship or no relationship could be expected if the same ecotypes were compared at sites with higher evapotranspiration demand.
Different adaptive mechanisms in different ecotypes grow-ing in distinct conditions may result in apparently contradic-tory performances, indicating that comparison ofδ13C studies must be carefully approached. It has been shown that the growth versus carbon isotope discrimination relationships in several species may change when plants are tested in different environments (Zhang et al. 1993, Flanagan and Johnsen 1995, Condon et al. 2004), demonstrating variability in adaptive mechanisms. In our study, when correlations between all data values were analyzed for the Atlantic group, negative
correla-Figure 2. Genotype × Trait patterns for carbon isotope composition (δ13
C), total height, diameter at breast height (DBH), polycyclism and survival in the Atlantic group of populations: Landes; Bragança (Pb42); Alcácer do Sal (Pas54); Escaroupim (Pcse0102); Monção (Pm36); and Leiria (Pmg52). Abbreviations: PC1 and PC2 = principal components 1 and 2, respectively; and SVP, singular-value partition-ing.
tions were found betweenδ13C and height or survival in Esca-roupim. Differences inδ13C reflect differences in WUE, and these can be predominantly determined by stomatal conduc-tance or by assimilation rate, or both. The negative correlation that we found between needleδ13C and growth suggests that, in Escaroupim,δ13C is more closely controlled by stomatal conductance than by assimilation, as predicted by the Far-quhar model (Brendel et al. 2002).
Annual total precipitation did not explain the population dif-ferences in δ13C: Landes and Leiria, originating from environ-ments with similar rainfall, occupied opposite positions in δ13
C ranking, and populations originating from environments with distinctly different rainfalls (Alcácer and Monção) had similarδ13C. Although no association was found betweenδ13C values and P, a negative correlation betweenδ13C and m was revealed (Table 4). Landes (highestδ13C) and Leiria (lowest δ13
C) have similar annual rainfalls at their sites of origin, but Landes has colder winters. Among the Atlantic group of popu-lations, when water is not limiting, the length of the growing period may depend more on minimum temperatures in the cold season. Populations originating from warmer climates have longer vegetative periods and show more discrimination against13C during the longer period when needles are formed, leading to lower WUE. A preliminary analysis within the Por-tuguese group showed that variation inδ13C was negatively correlated to m and positively correlated to M, but more posi-tively correlated with the difference (M – m), which can be considered an indicator of habitat continentality. Overall, dif-ferences inδ13C among populations could be more closely re-lated to microenvironmental conditions or to short-term clima-tic variations than to regional differences in climate. Large variation was found among individuals within a population, suggesting a high dependence on localized conditions. This finding is consistent with maritime pine’s high phenotypic plasticity for drought tolerance revealed in several studies (Hopkins and Butcher 1993, Zas et al. 2004, Chambel et al. 2007).
In conclusion, significant differences in growth were found among 10-year-old populations from different geographic and climatic origins growing in a subhumid site, with Leiria dis-playing superior growth, but only small differences were found in polycyclism and needleδ13C. The relationship be-tween growth andδ13C differed between the Atlantic group and the Mediterranean population (Cuenca), which had low δ13
C (and WUE) and the lowest growth of the populations studied. This may reflect an adaptation to more xeric environ-ments, where plants are less dependent on stomatal control of water loss and invest more in root growth. Mesic ecotypes mainly rely on stomatal sensitivity to reduce transpiration and display high aboveground growth rates. Within mesic eco-types, populations like Leiria and Monção, which are adapted to mild winters similar to those at Escaroupim, may initiate stomatal closure later than populations from the coldest cli-mates when subjected to moderate water stress. This suggests a genetic control to maintain photosynthetic rate and high growth potential with reduced WUE. Growth was associated with P and m, whereas polycyclism was associated with P and
δ13C variation was negatively associated with m. The differ-ences found between some populations and the negative corre-lations betweenδ13C (and WUE) and height or survival indi-cate the potential for selection for forestry and conservation purposes under conditions of climate change. However, a larger sampling and evaluations at several sites are needed to gain further insight into the mechanism underlying the rela-tionships between δ13C values, growth and environmental variables.
Acknowledgments
We are grateful for the dedicated assistance provided by Eng. João Pessoa during needle collection and for the availability of Laboratório de Ecofisiologia Florestal in Departamento de Engenharia Florestal, Instituto Superior de Agronomia (Universidade Técnica de Lisboa), where the preparation of needle samples for carbon isotope analysis took place. We also thank Dr. Weikai Yan from the Eastern Cereal and Oilseed Research Centre (Canada) for providing GGE Biplot soft-ware for Genotype × Trait analysis. This study was funded by INIAP/ FCT project PARLE D, LT 4 - Contribuição para o estudo da varia-bilidade genética do pinhal (2001–2004).
References
Alía, R., L. Gil and J.A. Pardos. 1995. Performance of 43 Pinus
pinaster Ait. provenances in five locations in central Spain. Silvae
Genet. 44:75–81.
Alía, R., J. Moro and J.B. Denis. 1997. Performance of Pinus pinaster provenances in Spain: interpretation of the genotype by environ-ment interaction. Can. J. For. Res. 27:1548–1559.
Bahrman, N., P. Baradat and R. Petit. 1992. Structuration de la variabilité génétique du pin maritime dans l’ensemble de l'aire naturelle. Hypothèses explicatrices. In Complexes d'Espèces, Flux de Gènes et Ressources Génétiques des Plantes. Bureau des Ressources Génétiques Eds., Lasoisier, Cachan, pp 155–163. Baradat, P. and A. Marpeau-Bézard. 1988. Le pin maritime Pinus
pinaster Ait.: biologie et génétique des terpènes pour la
connaissance et l’amélioration de l’éspèce. Ph.D. Thesis, Université Bordeaux I, France, 510 p.
Brendel, O., D. Pot, C. Plomion, P. Rozenberg and J.M. Guehl. 2002. Genetic parameters and QTL analysis ofδ13C and ring width in maritime pine. Plant Cell Environ. 25:945–953.
Brugnoli, E. and G.D. Farquhar. 2000. Photosynthetic fractionation of carbon isotopes. Photosynthesis: physiology and metabolism. Adv. Photosynth. 9:399–434.
Chambel, M.R., J. Climent and R. Alía. 2007. Divergence among spe-cies and populations of Mediterranean pines in biomass allocation of seedlings grown under two watering regimes. Ann. For. Sci. 64:87–97.
Condon, A.G., R.A. Richards, G.J. Rebetzke and G.D. Farquhar. 2004. Breeding for high water-use efficiency. J. Exp. Bot. 55: 2447–2460.
Cregg, B.M. and J.W. Zhang. 2001. Physiology and morphology of Pinus sylvestris seedlings from diverse sources under cyclic drought stress. For. Ecol. Manage. 154:131–139.
Danjon, F. 1994. Stand features and height growth in a 36-year-old maritime pine (Pinus pinaster Ait.) provenance test. Silvae Genet. 43:52–62.
DeLucia, E.H. and W.H. Schlesinger. 1991. Resource-use efficiency and drought tolerance in adjacent Great Basin and Sierran plants. Ecology 72:51–58.
Farquhar, G.D., J.R. Ehleringer and K.T. Hubick. 1989. Carbon iso-tope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:503–537.
Flanagan, L.B. and K.H. Johnsen. 1995. Genetic variation in carbon isotope discrimination and its relationship to growth under field conditions in full-sib families of Picea mariana. Can. J. For. Res. 25:39–47.
González-Martínez, S.C., R. Alía and L. Gil. 2002. Population ge-netic structure in a Mediterranean pine (Pinus pinaster Ait.): a comparison of allozyme markers and quantitative traits. Heredity 89:199–206.
Guehl, J.M., A. Nguyen-Queyrens, D. Loustau and A. Ferhi. 1995. Genetic and environmental determinants of water-use efficiency and carbon isotope discrimination in forest trees. In EUROSILVA —contribution to forest tree physiology. Eds. M. Bonnet-Masim-bert and H. Sandermanns. Editions Colloquies de l’INRA, Paris, pp 297–321.
Guy, R.D. and D.L. Holowachuk. 2001. Population differences in sta-ble carbon isotope ratio of Pinus contorta Dougl. ex Loud.: rela-tionship to environment, climate of origin, and growth potential. Can. J. Bot. 79:274–283.
Guyon, J.P. and A. Kremer. 1982. Phenotypic stability of height growth, daily changes in sap pressure and transpiration in maritime pine (Pinus pinaster Ait.). Can. J. For. Res. 12:936–946. Harfouche, A., P. Baradat and C.E. Durel. 1995. Variabilité
intra-spécifique chez le pin maritime (Pinus pinaster Ait.) dans le sud-est de la France. I. Variabilité des populations autochtones et des populations de l’ensemble de l’aire de l’éspèce. Ann. Sci. For. 52:307–328.
Hopkins, E.R. and T.B. Butcher. 1993. Provenance comparisons of
Pinus pinaster Ait. in Western Australia. CalmScience 1:55–105.
Hubick, K.T. and A. Gibson. 1993. Diversity in the relationship be-tween carbon isotope discrimination and transpiration efficiency when water is limited. In Stable Isotopes and Plant Carbon Water Relations. Eds. J.R. Ehleringer, A.E. Hall and G.D. Farquhar. Aca-demic Press, New York, pp 311–324.
Johnsen, K.H., L.B. Flanagan, D.A. Huber and J.E. Major. 1999. Ge-netic variation in growth, carbon isotope discrimination, and foliar N concentration in Picea mariana: analyses from a half-diallel mating design using field-grown trees. Can. J. For. Res. 29: 1727–1735.
Jones, H.G. 1992. Plants and microclimate: a quantitative approach to environmental plant physiology. 2nd Edn. Cambridge University Press, New York, 452 p.
Kremer, A. 1981. Déterminisme génétique de la croissance en hauteur du pin maritime (Pinus pinaster Ait.). 1. Role du polycyclisme. Ann. Sci. For. 38:199–222.
Kremer, A. and G. Roussel. 1986. Décomposition de la croissance en hauteur du pin maritime. Variabilité géographique des compo-santes morphogénétiques et phénologiques. Ann. Sci. For. 43: 15–34.
Máguas, C. and H. Griffiths. 2003. Applications of stable isotopes in plant ecology. In Progress in Botany, Vol. 64. Eds. K. Esser, U. Lüttge, W. Beyschlag and F. Hellwig. Springer-Verlag, Heidel-berg, Germany, pp 472–505.
Miranda, P.M.A., M.A. Valente, A.R. Tomé, R. Trigo, M.F.E.S. Coe-lho, A. Aguiar and E.B. Azevedo. 2006. O clima de Portugal nos séculos XX e XXI. In Alterações Climáticas em Portugal. Cenários, Impactes e Medidas de Adaptação. Eds. F.D. Santos and P. Miranda. Gradiva, Lisboa, pp 45–113.
Nguyen-Queyrens, A., D. Loustau, A. Ferhi and J.M. Guehl. 1998. Within-ringδ13C spatial variability and interannual variations in wood cellulose of two contrasting provenances of Pinus pinaster. Can. J. For. Res. 28:766–773.
Plomion, C., C.-E. Durel and D.M. O’Malley. 1996. Genetic dissec-tion of height in maritime pine seedlings raised under accelerated growth conditions. Theor. Appl. Genet. 93:849–858.
Prasolova, N.V., Z.H. Xu, G.D. Farquhar, P.G. Saffigna and M.J. Diet-ers. 2000. Variation in branchletδ13C in relation to branchlet
nitro-gen concentration and growth in 8-year-old hoop pine families (Araucaria cunninghamii) in subtropical Australia. Tree Physiol. 20:1049–1055.
Prasolova, N.V., Z. Xu, G.D. Farquhar, P.G. Saffigna and M.J. Diet-ers. 2001. Canopy carbon and oxygen isotope composition of 9-year-old hoop pine families in relation to seedling carbon isotope composition, growth, field growth performance, and canopy nitro-gen concentration. Can. J. For. Res. 31:673–681.
Prasolova, N.V., P.G. Saffigna, Z.H. Xu, M.J. Dieters, S. Walker, K. Lundkvist and G.D. Farquhar. 2003. Genetic variation in foliar carbon isotope composition in relation to tree growth and foliar ni-trogen concentration in clones of the F1 hybrid between slash pine and Caribbean pine. For. Ecol. Manage. 172:145–160.
Prasolova, N.V., Z.H. Xu and K. Lundkvist. 2005. Genetic variation in foliar nutrient concentration in relation to foliar carbon isotope composition and tree growth with clones of the F1 hybrid between slash pine and Caribbean pine. For. Ecol. Manage. 210:173–191. Read, J. and G.D. Farquhar. 1991. Comparative studies in
Noto-phagus (Fagaceae). I. Leaf carbon isotope discrimination. Funct.
Ecol. 5:684–695.
Ribeiro, M.M., C. Plomion, R. Petit, G.G. Vendramin and A.E. Szmidt. 2001. Variation of chloroplast simple-sequence re-peats in Portuguese maritime pine (Pinus pinaster Ait.). Theor. Appl. Genet. 102:97–103.
Rolf, H., K. Urs and G. Felix. 2006. Adaptive vs. neutral genetic di-versity: implications for landscape genetics. Landscape Ecol. 21: 797–807.
Stewart, P. 1968. Quotient pluviothermique et dégradation bio-sphérique. Bull. Soc. Hist. Nat. Afr. Nord. 59:23–36.
Tognetti, R., M. Michelozzi, M. Lauteri, E. Brugnoli and R. Giannini. 2000. Geographic variation in growth, carbon isotope discrimina-tion, and monoterpene composition in Pinus pinaster Ait. prove-nances. Can. J. For. Res. 30:1682–1690.
Xu, Z., N. Prasolova, T. Leaman, K. Lundkvist and C. Beadle. 2003. Genetic variation in branchlet carbon and nitrogen isotope compo-sition and nutrient concentration of 11-year-old hoop pine families in relation to tree growth in subtropical Australia. For. Ecol. Man-age. 186:359–371.
Yan, W. 2002. Singular-value partitioning in biplot analysis of multi-environment trial data. Agron. J. 94:990–996.
Zas, R., E. Merlo and J. Fernández-López. 2004. Genotype x environ-ment interaction in maritime pine families in Galicia, northwest Spain. Silvae Genet. 53:175–182.
Zhang, J., J.D. Marshall and B.C. Jaquish. 1993. Genetic differentia-tion in carbon isotope discriminadifferentia-tion and gas exchange in
Pseudo-tsuga menziesii. Oecologia 93:80–87.
Zhang, J.W., Z. Feng, B.M. Cregg and C.M. Schumann. 1997. Carbon isotopic composition, gas exchange, and growth of three popula-tions of ponderosa pine differing in drought tolerance. Tree Physiol. 17:461–466.