Tephrosia toxicaria
Pers. reduces temporomandibular joint
inflammatory hypernociception: The involvement of the
HO-1 pathway
D.R. do Val1, M.M. Bezerra2, A.A.R. Silva3, K.M.A. Pereira3, L.C. Rios2, J.C. Lemos2, N.C. Arriaga2,
J.N. Vasconcelos4, N.M.B. Benevides5, V.P.T. Pinto2, G. Cristino-Filho2, G.A.C. Brito6, F.R.L. Silva4,
G.M.P. Santiago4, A.M.C. Arriaga4, H.V. Chaves3
1 Master of Biotechnology Degree Program, Federal University of Ceará, Sobral, Brazil
2 Faculty of Medicine, Master of Biotechnology Degree Program, Federal University of Ceará, Sobral, Brazil 3 Faculty of Dentistry, Master of Healthy Sciences Degree Program, Federal University of Ceará, Sobral, Brazil 4 Department of Chemistry, Federal University of Ceará, Fortaleza, Brazil
5 Department of Biochemistry and Molecular Biology, Federal University of Ceará, Fortaleza, Brazil 6 Department of Morphology, Federal University of Ceará, Fortaleza, Brazil
Correspondence Hellíada V. Chaves
E-mail: [email protected]
Funding sources
This work was supported by Brazilian grants from Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP), Conselho Nacional de Pesquisa (CNPq) and Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflicts of interest None declared.
Accepted for publication 16 February 2014
doi:10.1002/j.1532-2149.2014.488.x
Abstract
Background: We investigated both the efficacy and the sub-chronic tox-icity ofTephrosia toxicariaPers. in the zymosan-induced temporomandibular joint (TMJ) inflammatory hypernociception in rats evaluating the possible role of heme oxygenase-1 (HO-1).
Methods: Rats were pretreated with T. toxicaria (0.2, 2.0 or 20 mg/kg) 60 min before the intra-articular injection of zymosan (2 mg, 40μL) in the left TMJ. In another series of experiments, rats were treated with ZnPP-IX (3 mg/kg), a specific HO-1 inhibitor, before T. toxicaria (20 mg/kg). Von Frey test was used to evaluate inflammatory hypernociception (g) 4 h after zymosan injection. Six hours after zymosan injection, the synovial lavage was collected for total cell count and myeloperoxidase (MPO) activity, and joint tissue for histopathological analysis and immunohistochemistry for HO-1. To evaluate the sub-chronic toxicity, mice received T. toxicaria (20 mg/kg) or saline once a day for 14 days to analyse body mass, organ weight and biochemical parameters.
Results: T. toxicaria partially reversed the zymosan-induced head withdrawal threshold, the number of cells and the MPO activity.T. toxicaria reduced the inflammatory cell influx in the synovial membrane. TMJ immunohistochemical analyses treated withT. toxicariashowed increased HO-1 expression. These effects of T. toxicaria were not observed in the presence of ZnPP-IX. T. toxicaria treatment for 14 days did not show significant signs of toxicity when administrated to mice.
Conclusions: T. toxicariadid not produce any signs of toxicity and effec-tively decreased zymosan-induced TMJ inflammatory hypernociception dependent, at least in part, upon the HO-1 pathway integrity.
1. Introduction
Temporomandibular joint (TMJ) disorders are a group of conditions that result in TMJ pain, which frequently
inflamma-tory and pain conditions are of great clinical relevance. In this regard, we have developed a rat model of TMJ inflammation using intra-articular (i.art.) injections of pro-inflammatory agent, zymosan (Chaves et al., 2011). Zymosan is a polysaccharide from yeast cell walls that produces a severe and erosive synovitis (Gegout et al., 1994; Bezerra et al., 2004) associated with inflammatory pain in animal models of knee arthritis (Bezerra et al., 2007).
Over the past few years, numerous studies have led to the appreciation of heme oxygenase 1 (HO-1). To date, three isoforms of HO (HO-1, HO-2 and HO-3) have been identified. HO-1 is inducible (Bednarz et al., 2007; Bezerra et al., 2008), whereas HO-2 and HO-3 are constitutively expressed (Bezerra et al., 2008). HO-1 is induced in a variety of cells including endothelial cells, monocytes/macrophages, neutro-phils and fibroblasts by heme, endotoxins, cytokines, nitric oxide (NO) and other mediators produced during inflammatory responses (Botros et al., 2005; Bressan and Tonussi, 2008; Choi et al., 2008), and its induction would provide a negative feedback for cell activation and the production of inflammatory media-tors, beyond its role in the antioxidant defence system, which could modulate, at least in part, the inflamma-tory pain process (Alcaraz et al., 2003; Grangeiro et al., 2011; Vanderlei et al., 2011).
In order to develop potential tools for new therapies to ameliorate inflammatory pain, there has been a continuous increase in the use of natural products in the past decades, which has encouraged scientific studies to search for new substances with therapeutic action and to confirm the efficacy of medicines derived from plants.
Tephrosia (Fabaceae) is a large perennial genus, which is distributed in the tropical and subtropical
regions of the world (Tarus et al., 2002).Tephrosia toxi-cariaPers., also namedT. sinapou(Buc’hoz) A. Chev., is a shrub that has been used in Amazonian countries traditional medicine to alleviate genital inflammation and pain due to gonorrhoea (Moretti and Grenand, 1982). Our group has reported that this species are a source of flavonoids, isoflavonoids and rotenoids (Vasconcelos et al., 2009) that possess various biologi-cal effects such as antioxidative and larvicidal (Vasconcelos et al., 2012), besides cancer chemopre-ventive (Udeani et al., 1997; Lee et al., 2005) and anti-inflammatory (Martinez et al., 2012).
Thus, the present study was aimed at investigating the unexplored antinociceptive and anti-inflammatory efficacy ofT. toxicariain the model of zymosan-induced TMJ inflammatory hypernociception in the rat. Addi-tionally, considering our previous findings showing that the inhibition of HO-1 pathway is associated with worsening of inflammatory response (Grangeiro et al., 2011), we also investigated whetherT. toxicariaefficacy depends upon the integrity of the HO-1 pathway. Further, a systemic evaluation of the sub-chronic tox-icity ofT. toxicariawas also performed.
2. Methods
2.1 Plant materials
T. toxicariaPers. was collected in Guaraciaba do Norte, Ceará State, Brazil. A voucher specimen (# 32.139) was deposited at the Prisco Bezerra Herbarium, Federal University of Ceará, Brazil.
2.2 Extraction
The air-dried and powdered roots (710.0 g) of T. toxicaria were extracted with ethanol at room temperature to afford a crude extract (78.9 g) in 11.1% yields on fresh weight basis.
2.3 Animals
Male Wistar rats (160–220 g) (n=60) and Swiss mice (20– 25 g) (n=20) were housed in standard plastic cages with food and water availablead libitum. They were maintained in a temperature-controlled room (23±2 °C) with a 12/12-h light–dark cycle. All experiments were designed to minimize animal suffering and to use the minimum number of animals required to achieve a valid statistical evaluation. This study was conducted in accordance with the Institutional Animal Care and with the approval of the Ethics Committee of the Federal University of Ceara, Fortaleza, Brazil (CEPA No. 57/10).
What’s already known about this topic? • Temporomandibular joint (TMJ) disorders are
associated with high levels of inflammatory pain-related disability.Tephrosia toxicariais a shrub that has been used in Amazonian countries tradi-tional medicine to alleviate inflammation and pain.
What does this study add?
2.4 Induction of TMJ inflammatory hypernociception
Rats were anaesthetized with tribromoethanol (1 mL/100 g, i.p.) and received an i.art. injection of 2 mg zymosan (40μL total volume) dissolved in sterile saline into the left TMJ using a 30-gauge needle and a 0.5-mL syringe. Sham animals received only saline i.art. Before zymosan or saline injec-tions, the TMJ skin region was carefully shaved, the postero-inferior border of the zygomatic arch was palpated, and the needle was inserted inferior to this point and advanced in a medial and anterior direction until the needle made contact with the condyle. This contact was verified by moving the mandible, and the puncture of the needle into the joint space was confirmed by the loss of resistance. Gentle aspiration ruled out intravascular placement, after which the specified volume of zymosan or saline was injected.
As shown by our group previously (Chaves et al., 2011) during the time course of zymosan TMJ inflammatory hypernociception development, it is maximal at 4 h of arthritis whereas cell influx peaks at 6 h. Based upon these results, we used these time points to assess both nociceptive (head withdrawal threshold) and inflammatory parameters [total cell counting and myeloperoxidase (MPO) assay].
2.5 Evaluation of inflammatory hypernociception
Inflammatory hypernociception in the TMJ was evaluated by measuring the threshold of force intensity that needed to be applied to the TMJ region until the occurrence of a reflex response of the animal (e.g., head withdrawal). The mea-surements were performed by an examiner unaware of the treatments and used a digital device (Insight, Ribeirão Preto, SP, Brazil) that consisted of a rigid filament linked with an electronic device that measured the response threshold in grams (g) when the filament was applied to the surface of the tested region (Denadai-Souza et al., 2009). The facial areas to be tested around the TMJ were carefully shaved, and the animals were placed into individual plastic cages 45 min before the beginning of the tests. The animals were submit-ted to a conditioning session of head withdrawal threshold measurements in the testing room for 4 consecutive days under controlled temperatures (23±2 °C) and low illumina-tion. On the fifth day, the basal force threshold value was recorded (in triplicate) before the i.art. injections of either zymosan or vehicle and after 4 h.
2.6 Synovial lavage collection and cell counting
Six hours (6 h) after zymosan injections, the rats were sac-rificed under anaesthesia and exsanguinated. The superficial tissues were dissected, and the TMJ cavity was washed to collect the synovial lavage by a pumping and aspiration technique using 0.05 mL of ethylenediaminetetraacetic acid
(EDTA) in neutral phosphate buffered saline (PBS). This procedure was repeated twice. The total number of white cells in the synovial lavage was counted using a Neubauer chamber.
2.7 MPO activity analysis
MPO is an enzyme found primarily in the azurophilic gran-ules of the neutrophils and has been used extensively as a biochemical marker of granulocyte infiltration into various tissues. The MPO activity assay has been described previ-ously by Bradley et al. (1982). In our study, the MPO assay was conducted on the collected synovial lavage 6 h after zymosan injection. Briefly, the synovial lavage was centri-fuged at 4500 rpm for 12 min at 4 °C. MPO activity was assayed by measuring the change in absorbance at 450 nm usingo-dianisidine dihydrochloride and 1% hydrogen per-oxide. The results are reported as the MPO units/joint fluid. A unit of MPO activity was defined as the conversion of 1μmol of hydrogen peroxide to water in 1 min at 22 °C.
2.8 Histopathological analysis
After sacrifice at 6 h after zymosan-induced TMJ inflamma-tory hypernociception, the TMJ was excised. The specimens were fixed in 10% neutral buffered formalin for 24 h, demineralized in 10% EDTA, embedded in paraffin and sec-tioned along the long axis of the TMJ. Sections of 5μm, which included the condyle, articular cartilage, articular disc, synovial membrane, periarticular tissue and the skeletal muscle periarticular tissue, were evaluated under light microscopy.
For the specimens processed for routine haematoxylin-eosin (H&E) staining, histological analysis considered a 0–4 score grade based upon the cell influx in the synovial mem-brane (SM).
2.9 Immunohistochemistry for HO-1
Immunohistochemistry for HO-1 was performed using the streptavidin-biotin (labelled streptavidin biotin – LSAB) method in formalin-fixed, paraffin-embedded tissue sections (5μm thick), mounted on glass slides pre-viously prepared with an organosilane-based adhesive (3-aminopropyltriethoxysilane). Briefly, it consisted of the following steps: the sections went through two baths in xylol, each lasting 10 min. After this, they were immersed in three passages of absolute alcohol, then washed in running water, and immediately after this, a passage in distilled water. Antigen recovery was performed with citrate at pH 6.0 for 30 min at 99 °C. After returning to ambient temperature, the sections were immersed in a 3% hydrogen peroxide blocking solution for 10 min. After returning to ambient temperature, the sections were incubated overnight (4 °C) with a primary rabbit anti-HO-1 antibody (ab13243, ABCAM®, England,
The samples were incubated with the secondary antibody LSAB Kit for 10 min at ambient temperature. Next, incuba-tion was performed in a chromogen soluincuba-tion prepared with 3,3′diaminobenzidine for 10 min in a dark chamber. After-wards, the specimens were washed in running water and then in distilled water. Counter-staining was performed with haematoxylin, and afterwards the specimens were dehy-drated in alcohol and diaphanized in xylol. Lastly, they were mounted on glass slides, which were examined under optical microscope. The negative control sections were performed, excluding the application of the primary antibody. The parameter of positivity for the immunohistochemical marking of the antigen in all the specimens included in the sample consisted of the cells that exhibited brown staining in their cytoplasm, irrespective of the intensity of the immunomarking.
2.10 Treatment
T. toxicaria(0.2, 2 or 20 mg/kg) was diluted in 10μL of Tween 80 and the final volume of solution was adjusted using 0.9% sterile saline. T. toxicaria (0.2, 2 or 20 mg/kg; 1 mL/200 g) was injected (per os) 1 h prior to zymosan i.art. Zymosan group consisted of rats that received (per os) T. toxicaria vehicle (0.9% sterile saline; 1 mL/200 g) 1 h prior to zymosan i.art. To validate the data, a positive control group of rats was pretreated (s.c.) with indomethacin (5 mg/kg) 1 h before zymosan injection. Sham group received (per osand i.art.) 0.9% sterile saline.
To analyse the possible effect of HO-1 pathway on antinociceptive and anti-inflammatory efficacy ofT. toxicaria in the model of zymosan-induced TMJ inflammatory hyper-nociception in the rat, another series of experiments was performed. Animals were pretreated (s.c.) with ZnPP IX (3 mg/kg), a specific HO-1 inhibitor, alone or followed by an injection (per os) ofT. toxicaria(20 mg/kg) 30 min later. After 1 h, zymosan (2 mg) was injected (i.art.), and on the fourth hour of inflammatory hypernociception in the TMJ was evaluated as described above.
2.11 Sub-chronic toxicity ofT. toxicaria
Body mass, organ weight changes and blood levels of the biochemical parameters alanine amino transferase (ALT) and aspartate amino transferase (AST) were evaluated after once-daily treatment of 20 mice withT. toxicaria(20 mg/kg) or 0.9% sterile saline for 14 days. Mice were weighed daily, and after 14 days of treatment, samples of peripheral blood were collected for biochemical analysis (determined by enzy-matic and colorimetric tests – LABTEST). After sacrificing the animal, the liver, kidney and heart were removed and weighed, and possible ulcerative lesions or haemorrhaging were quantified and macroscopically measured.
2.12 Statistical analysis
The data are presented as means±SD or medians, where appropriate. Differences between means were compared
using a one-way analysis of variance, followed by the Stu-dent’st-test or the Bonferroni test. The Kruskal–Wallis test, followed by Dunn’s test was used to compare medians. A probability value ofp<0.05 indicated significant differences.
3. Results
3.1 Efficacy ofT. toxicariaon the zymosan-induced TMJ
inflammatory hypernociception
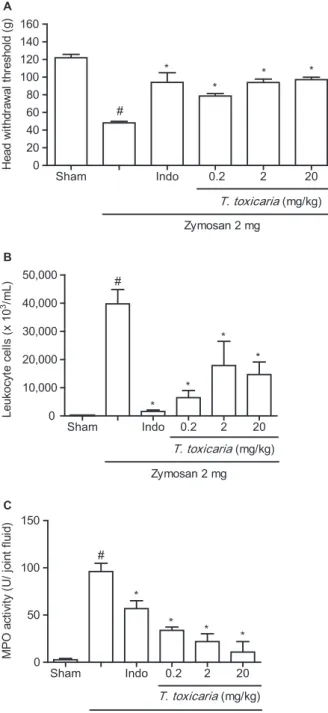
The i.art. 2 mg injection of zymosan resulted in inflammatory hypernociception as measured by a clear decrease in the mechanical threshold for head withdrawal at 4 h (Fig. 1A). Further, a 2 mg injection of zymosan resulted in a significant increase in the number of polymorphonuclear cells (Fig. 1B). This increase in neutrophils was certified by the increase of MPO activity in the TMJ synovial lavage following zymosan injection (Fig. 1C). In the intra-articular saline-injected animals (sham), no significant changes in withdrawal thresholds, polymorphonuclear cells counts or MPO activity were observed (Fig. 1A–C). T. toxicaria (0.2, 2 or 20 mg/kg) injected (per os) 1 h prior to zymosan i.art. injection significantly (p<0.05) partially reversed zymosan-induced head withdrawal threshold (Fig. 1A). Further, T. toxicaria (0.2, 2 or 20 mg/kg) significantly (p<0.05) decreased the number of polymorphonuclear cells (Fig. 1B). This decrease in neutrophils was certified by the decrease of MPO activity in the synovial lavage (Fig. 1C).
3.2 Effect of zinc protoporphyrin IX (ZnPP IX), a specific HO-1 inhibitor, on theT. toxicaria
efficacy on the zymosan-induced TMJ inflammatory hypernociception
To investigate the role of HO-1 activity in the anti-inflammatory effect of T. toxicaria, the animals were pretreated (s.c.) with ZnPP IX (3 mg/kg), a specific HO-1 inhibitor. The effects ofT. toxicaria(20 mg/kg) on the zymosan-induced TMJ inflammatory hypernoci-ception (Fig. 2A) and inflammation [polymorphonu-clear cells counts (Fig. 2B)] and MPO activity in the TMJ synovial lavage (Fig. 2C) were not observed in the presence of ZnPP-IX (3 mg/kg).
3.3 Histopathological analysis
neutrophils, which characterized acute inflammation. Oedema was also observed in the synovium (Fig. 3B). Table 1 shows the scores attributed to TMJ’s his-topathological analysis and compares the values between the TMJs of sham and zymosan groups. A significant (p<0.05) increase in the inflammatory parameters was observed in the zymosan group.
T. toxicaria (20 mg/kg) reduced the inflammatory parameters to a normal status with a lower inflamma-tory cell influx in the SM (Fig. 3C). Table 1 shows the
scores attributed to TMJ’s histopathological analysis and compares the values between the TMJ of zymosan and T. toxicaria (20 mg/kg) groups. A significant (p<0.05) reduction in the inflammatory parameters was observed. Further, the effects of T. toxicaria (20 mg/kg) on the TMJ’s histopathological analysis (H&E) in the SM was not observed in the presence of ZnPP-IX (3 mg/kg) (Fig. 3D).
3.4 Immunohistochemical analysis
An immunohistochemical analysis for HO-1 showed an increase in HO-1 expression that was characterized by brown-coloured cells in the SM of the zymosan-induced TMJ inflammatory hypernociception and treated withT. toxicaria(20 mg/kg) (Fig. 4C). The cell types that expressed HO-1 were the synovial cells in the synovial membrane. Decreased immunoreactivity to HO-1 in synoviocites and neutrophils of the zymosan-induced TMJ inflammatory hypernocicep-tion was also observed (Fig. 4B). The negative control group sections were composed of zymosan-induced TMJ inflammatory hypernociception that were not treated with an anti-HO-1 antibody (Fig. 4A). None of the negative controls showed HO-1 immunoreactivity.
3.5 Sub-chronic toxicity ofT. toxicaria
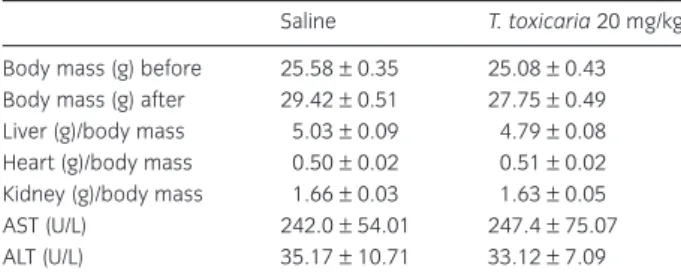
Repeated injections (per os) ofT. toxicaria (20 mg/kg) over 14 consecutive days did not produce any signs of toxicity in mice. The overall body mass and the weights of the liver, kidney and heart (wet basis) were normal. Serum levels of the enzymatic markers of hepatic function (ALT and AST) did not differ from the respective controls (Table 2).
A
Sham Indo 0.2 2 20
0 20 40 60 80 100 120 140 160
T. toxicaria (mg/kg)
Zymosan 2 mg *
*
* *
#
H
e
a
d
w
it
hdr
aw
al
t
h
re
sh
ol
d (
g
)
B
Sham Indo 0.2 2 20
0 10,000 20,000 30,000 40,000 50,000
T. toxicaria(mg/kg)
Zymosan 2 mg *
* *
* #
Leukocyt
e cel
ls
(
x
10
3/m
L
)
C
Sham Indo 0.2 2 20 0
50 100 150
*
* *
T. toxicaria(mg/kg)
Zymosan 2 mg *
#
M
P
O
a
ct
iv
ity
(U
/ jo
in
t flu
id
)
Figure 1 Efficacy ofTephrosia toxicariaPers. on zymosan-induced tem-poromandibular joint (TMJ) inflammatory hypernociception and inflam-mation. Zymosan (2 mg; 40μL) or saline was injected intra-articularly (i.art.) into the left TMJ of the rat.T. toxicaria(0.2, 2 or 20 mg/kg,per os) or indomethacin (5 mg/kg, s.c.) 1 h prior to zymosan i.art. injection. (A) Head withdrawal threshold in T. toxicaria Pers.-treated animals: The mechanical nociceptive threshold was measured inT. toxicaria-treated animals before and 4 h after an i.art. injection of zymosan. (B) Leukocyte counting inT. toxicariaPers.-treated animals: 6 h after zymosan injection (i.art.), leukocyte migration was evaluated inT. toxicaria-treated animals by cell counting in TMJ synovial lavage. (C) Myeloperoxidase (MPO) activ-ity from TMJ synovial lavage inT. toxicariaPers.-treated animals. The MPO activity was measured in TMJ synovial lavage 6 h after zymosan injection (i.art.). Data are expressed as mean±SD of 6 mice for each group.
#p<0.05 indicates a significant difference from the sham group, *p<0.05
indicates a significant difference from the zymosan group (analysis of variance, Bonferroni).
4. Discussion
In this work, we demonstrated the antinociceptive and anti-inflammatory efficacy of T. toxicariaPers. in the model of zymosan-induced TMJ inflammatory hypernociception in rats. We also found evidence
that, at least in part, the antinociceptive and anti-inflammatory effects of T. toxicaria in zymosan-induced TMJ inflammatory hypernociception depends upon the integrity of the HO-1 pathway.
Zymosan has been used as a pro-inflammatory agent for some time (Gadó and Gigler, 1991). To our knowledge, we performed the first demonstration of a zymosan-induced TMJ inflammatory hypernocicep-tion describing the histopathological inflammatory parameters involving the articular and periarticular tissues in an experimental TMJ arthritis (Chaves et al., 2011). Acute rat knee joint zymosan arthritis is char-acterized by an increase in vascular permeability, cell migration and pain. Later, a progressive synovitis occurs with mononuclear cell infiltration and a fibro-blast reaction (Keystone et al., 1977; Rocha et al., 1999). Our group has also demonstrated articular car-tilage and subchondral bone degradation during the chronic stages of zymosan arthritis in the rat knee joint (Bezerra et al., 2004).
In the present study, the i.art. 2 mg injection of zymosan decreased the head withdrawal threshold, which was partially increased byT. toxicariatreatment. Regarding the inflammatory parameters, T. toxicaria administration decreased the number of polymorpho-nuclear cells, which was certified by the decrease of the MPO activity in the synovial lavage. Further, the TMJ’s histopathological analysis from rats treated with T. toxicariacorrelated with the decrease in both leuko-cyte influx and MPO activity, since this treatment reduced the inflammatory parameters to a normal status with a lower inflammatory cell influx in the synovial membrane.
Beyond larvicidal activity ofT. toxicariaagainstAedes aegypti –the main vector of dengue fever – presented A
Sham Indo
0 20 40 60 80 100 120 140 160
ZnPP-IX 3 mg/kg
T. toxicaria 20 mg/kg
Zymosan 2 mg
* *
# & #
H
e
a
d
w
it
hdr
aw
al
t
h
re
sh
ol
d (
g
)
B
Sham Indo 0
10,000 20,000 30,000 40,000 50,000
Zymosan 2 mg *
*
ZnPP-IX 3 mg/kg
T. toxicaria 20 mg/kg #
& #
Leukocyt
e cel
ls
(
x
10
3/m
L
)
C
Sham Indo 0
50 100 150
*
*
T. toxicaria 20 mg/kg
Zymosan 2 mg #
ZnPP-IX 3 mg/kg
& #
M
P
O
a
c
ti
vit
y (
U
/ jo
in
t f
lu
id
)
Figure 2 Effect of zinc protoporphyrin IX (ZnPP IX), a specific HO-1 inhibi-tor, on theTephrosia toxicaria Pers. efficacy on zymosan-induced tem-poromandibular joint (TMJ) inflammatory hypernociception and inflammation. Animals were pretreated (s.c.) with ZnPP IX (3 mg/kg) alone or followed by an injection (per os) ofT. toxicaria(20 mg/kg) 30 min later. After 1 h, zymosan (2 mg) was injected (i.art.). Also, indomethacin (5 mg/ kg, s.c.) was injected 1 h prior to zymosan i.art. injection. (A) Head with-drawal threshold in ZnPP IX +T. toxicariaPers.-treated animals: The mechanical nociceptive threshold was measured before and 4 h after an i.art. injection of zymosan. (B) Leukocyte counting in ZnPP IX +T. toxi-caria Pers.-treated animals: 6 h after zymosan injection (i.art.), leukocyte migration was evaluated by cell counting in TMJ synovial lavage. (C) Myeloperoxidase (MPO) activity from TMJ synovial lavage in ZnPP
IX +T. toxicariaPers.-treated animals. The MPO activity was measured in
TMJ fluid 6 h after zymosan injection (i.art.). Data are expressed as = mean±SD of 6 mice for each group.#p<0.05 indicates a significant
difference from the sham group, *p<0.05 indicates a significant differ-ence from the zymosan group,&p<0.05 indicates a significant difference
lately by our group (Vasconcelos et al., 2012), some authors have shown that extracts obtained from plants ofTephosiagenus have different actions such as inhi-bition of inflammatory total leukocytes and neutrophil recruitment induced by a variety of inflammatory stimuli in mice (Martinez et al., 2012), wound healing (Lodhi et al., 2006), hepatoprotective effects (Khatri et al., 2009), cancer chemopreventive action (Jang et al., 2003), reduction of oxidative stress in rats (Khan et al., 2001) and inhibition of delayed-type hypersensitivity reactionin vivo(Damre et al., 2003).
Table 1Histopathological analysis of temporomandibular joint in zymosan-induced inflammatory hypernociception: the effects of Tephro-sia toxicariaPers.
Groups Cell influx in the synovial membrane
Sham 0 (0–0)
Zymosan 3 (1–4)*
Indomethacin 0 (0–1)**
T. toxicaria(20 mg/kg) 1 (0–3)**
T. toxicaria(20 mg/kg) + ZnPP-IX 3 (2–4)***
*p<0.05 versus sham. **p<0.05 versus zymosan.
***p<0.05 versusT. toxicaria(20 mg/kg);n= 6 (Kruskal–Wallis, Dunn’s). A
AC
SM
C
PAT
D B
AD
C
C
PAT AC AD
SM
PAT
SM
AD
AC
C C
AC AD
SM
Figure 3Photomicrographs of the histopatho-logical analysis of temporomandibular joints (TMJs): (A) sham group TMJ (40×); (B) TMJ inflammatory hypernociception induced by zymosan 2 mg (40×) showing inflammatory cell influx in the synovial membrane; (C) TMJ of rats pretreated (per os) withTephrosia toxicaria (20 mg/kg) and injected (i.art.) with zymosan 2 mg (40×) showing reduction of the inflamma-tory parameters to a normal status with a lower inflammatory cell influx in the synovial membrane; and (D) TMJ of rats pretreated (s.c.) with ZnPP IX (3 mg/kg), a specific HO-1 inhibi-tor, followed by an injection (per os) ofT. toxi-caria(20 mg/kg) and injected (i.art.) with zymosan 2 mg (40×). ZnPP-IX reverted the effects ofT. toxicaria(20 mg/kg) on the TMJ histopathological analysis (H&E) in the synovial membrane. C, condyle; AC, articular cartilage; AD, articular disc; SM, synovial membrane; PAT, periarticular tissue. Haematoxylin and eosin (H&E) staining.
A B C
Although the mechanisms ofTephosiagenus remain to be determined, one study provided pre-clinical evi-dence that anti-inflammatory activity of an extract of T. toxicariais related to inhibition of pro-inflammatory cytokine (tumour necrosis factor-α and interleukin-1β) production and NO-dependent inhibition of leu-kocyte recruitment in mice. Additionally,in vitrodata demonstrated that the extract inhibits oxidative stress by scavenging free radicals, iron chelating activity and inhibition of iron-dependent and iron-independent lipoperoxidation (Martinez et al., 2012).
In the present study, histopathological analyses of immunohistochemical stains showed HO-1 expression in the synovial cells and neutrophils of the zymosan-induced TMJ inflammatory hypernociception. Heme oxygenase (HO) is the rate-limiting enzyme that catalyses the degradation of heme to liberate carbon monoxide (CO), biliverdin (BVD) and free iron in mammalian cells (Alcaraz et al., 2003). HO-1 is induced by oxidative or nitrosative stress, cytokines and other mediators produced during inflammatory processes, likely as part of a defence system in cells exposed to stress to provide a negative feedback for cell activation and the production of mediators, which could modulate the inflammatory response (Alcaraz et al., 2003). Over the last few years, numerous studies have demonstrated that HO-1 expression and the concomitant production of its metabolites, CO and BVD have anti-inflammatory consequences (Vane and Warner, 2000; Soares et al., 2004; Majewska et al., 2008). Considering this, we demonstrated that the HO/BVD/CO pathway plays antinociceptive effects during acetic acid-evoked nociception (Grangeiro et al., 2011). In fact, heme-induced HO-1 was reported to result in a reduction of cell migration, exudation and pro-inflammatory mediators release in a zymosan-induced air pouch inflammation model
(Vicente et al., 2003). There is evidence that CO stimulates soluble guanylate cyclase activity and increases the cellular levels of cyclic guanosine mono-phosphate (cGMP) (Dearden et al., 1989; Morita et al., 1995). Ferreira et al. (1991) provided experimental support to suggest that elevated levels of cyclic GMP are associated with inhibition of nociceptor hypersen-sitivity. In this regard, our research group demon-strated increased antinociceptive response produced by the combination of agents that increase intracellu-lar cyclic GMP concentrations (Bezerra et al., 2008).
In the present study, after the pretreatment with ZnPP IX, a specific HO-1 inhibitor, the antinociceptive and anti-inflammatory efficacy of T. toxicaria in zymosan-induced TMJ inflammatory hypernocicep-tion in rats was not observed, suggesting that HO-1 activity is involved in the inhibitory effects ofT. toxi-caria, corroborating other data showing that the inhi-bition of HO-1 pathway is associated with worsening of inflammatory response (Alcaraz et al., 2003; Vicente et al., 2003; Bednarz et al., 2007). In this regard, we have recently demonstrated that ZnPP IX treatment potentiated the effect of acetic acid by increasing the number of writhes (Grangeiro et al., 2011). Further, analyses of immunohistochemical stains showed an increase in HO-1 expression in the synovial cells in the SM of the zymosan-induced TMJ inflammatory hypernociception and treated with T. toxicaria. In fact, HO-1 activity results in the inhibi-tion of oxidative damage and apoptosis, with signifi-cant reductions in inflammatory events including oedema, leukocyte adhesion and migration, and pro-duction of inflammatory cytokines (Alcaraz et al., 2003).
Although there has been growing interest in the use of natural products in recent years, especially those derived from plants, most of them have been used broadly by local populations without having adequate information on the efficacy and probable toxicity asso-ciated with their use. Thus, research concerning both efficacy and toxicity profile of these acclaimed medici-nal plants must be largely encouraged (Calixto et al., 2001). Therefore, in the preset study, we also evaluated the sub-chronic toxicity ofT. toxicariaadministration.
The administration of any substance may bring about significant changes in the structure, function and even metabolic pathways. These alterations may produce clinical and laboratory findings that can be used to determine the extent of deleterious effect of foreign compound, including plant extract on the blood. Such laboratory investigations have been reported to be highly sensitive, accurate and reliable (Okonkwo et al., 2004). In this regard, in the present
Table 2Systemic effects ofTephrosia toxicariaPers. (20 mg/kg) in mice.
Saline T. toxicaria20 mg/kg
study, we performed biochemical analysis in periph-eral blood from animals treated withT. toxicaria. The analysis did not reveal any toxicity signals afterT. toxi-cariatreatment. This finding may be very stimulating sinceT. toxicariawas as effective as indomethacin, used as positive control, without showing any toxicity signals. To the best of our knowledge, this is the first report to explore theT. toxicariatoxicity profile.
In conclusion, we demonstrated the antinociceptive and anti-inflammatory efficacy of T. toxicaria in the model of zymosan-induced TMJ inflammatory hyper-nociception in rats. Additionally, our results strongly suggest thatT. toxicariaefficacy, at least in part, depends upon the integrity of the HO-1 pathway. Further, we verified that T. toxicaria in the effective dose did not produce any signs of toxicity. Taking into account the well-demonstrated antinociceptive and anti-inflammatory efficacy ofT. toxicaria, the designing of alternative compounds to classical anti-inflammatory and analgesic agents is very encouraged to define new pharmacological targets for the inflammatory TMJ painful condition treatment.
Author contributions
J.N.V. collected theTephrosia toxicariaPers.; F.R.L.S., G.M.P.S. and A.M.C.A. extracted the T. toxicaria Pers.; H.V.C. and M.M.B. designed the research; L.C.R., J.C.L., N.C.A., D.R.V. and H.V.C. performed the research; K.M.A.P. performed his-topathological analysis; D.R.V., A.A.R.S., V.P.T.P. and G.C.-F. performed and analysed theT. toxicariaPers. toxicity profile; N.M.B.B. and G.A.C.B. contributed new reagents/analytical tool; D.R.V., H.V.C. and M.M.B. analysed data; and D.R.V., H.V.C. and M.M.B. wrote the paper.
Acknowledgements
The authors thank Adalberto Nascimento de Lima Júnior, Angela Magalhães Vieira and Jordânia Marques de Oliveira for the technical assistance.
References
Alcaraz, M.J., Fernández, P., Guillén, M.I. (2003). Anti-inflammatory
actions of the heme oxygenase-1 pathway.Curr Pharm Des9, 2541–
2551.
Bednarz, N., Zawacka-Pankau, J., Kowalska, A. (2007). Protopor-phyrin IX induces apoptosis in HeLa cells prior to photo-dynamic treatment. Pharmacol Rep59, 474–479.
Bezerra, M.M., Brain, S.D., Girão, V.C.C., Greenacre, S., Keeble, J., Rocha, F.A.C. (2007). Neutrophils-derived peroxynitrite contributes to acute
hyperalgesia and cell influx in zymosan arthritis.Naunyn Schmiedebergs
Arch Pharmacol374, 265–273.
Bezerra, M.M., Brain, S.D., Greenacre, S., Jerônimo, S.M.B., Melo, L.B., Keeble, J., Rocha, F.A.C. (2004). Reactive nitrogen species scavenging, rather than nitric oxide inhibition, protects from articular cartilage
damage in rat zymosan-induced arthritis.Br J Pharmacol141, 172–182.
Bezerra, M.M., Lima, V., Girão, V.C.C., Teixeira, R.C., Graça, J.R.V. (2008). Antinociceptive activity of sildenafil and adrenergic agents in the
writh-ing test in mice.Pharmacol Rep60, 339–344.
Botros, F.T., Schwartzman, M.L., Stier, C.T., Goodman, A.I., Abraham, N.G. (2005). Increase in heme oxygenase-1 levels ameliorates
renovas-cular hypertension.Kidney Int68, 2745–2755.
Bradley, P.P., Christensen, R.D., Rothstein, G. (1982). Cellular and
extra-cellular myeloperoxidase in pyogenic inflammation.Blood60, 618–622.
Bressan, E., Tonussi, C.R. (2008). Antiinflammatory effects of etoricoxib alone and combined with NSAIDs in LPS-induced reactive arthritis. Inflamm Res57, 586–592.
Cairns, B.E. (2010). Pathophysiology of TMD pain – basic mechanisms
and their implications for pharmacotherapy.J Oral Rehabil37, 391–410.
Calixto, J.B., Scheidt, C., Otuki, M., Santos, A.R. (2001). Biological
activ-ity of plant extracts: Novel analgesic drugs.Expert Opin Emerg Drugs6,
261–279.
Chaves, H.V., Ribeiro, R.A., de Souza, A.M., Silva, A.A.R., Gomes, A.S., Vale, M.L., Bezerra, M.M., Brito, G.A. (2011). Experimental model of zymosan-induced arthritis in the rat temporomandibular joint: Role of
nitric oxide and neutrophils.J Biomed Biotechnol2011, 2001–2011.
Choi, H.C., Kim, H.S., Lee, K.Y., Chang, K.C., Kang, Y.J. (2008). NS-398,
a selective COX-2 inhibitor, inhibits proliferation of IL-1β-stimulated
vascular smooth muscle cells by induction of HO-1.Biochem Biophys Res
Commun376, 753–757.
Damre, A.S., Gokhale, A.B., Phadke, A.S., Kulkarni, K.R., Saraf, M.N. (2003). Studies on the immunomodulatory activity of flavonoidal
frac-tion ofTephrosia purpurea.Fitoterapia74, 257–261.
Dearden, D.V., Hayashibara, K., Beauchamp, J.L., Kirchner, N.J., Van Koppen, P.A.M., Bowera, M.T. (1989). Fundamental studies of the energetics and dynamics of ligand dissociation and exchange processes
at transition-metal centers in the gas phase: Mn(CO)x+, x=1–6.J Am
Chem Soc111, 2401–2409.
Denadai-Souza, A., Camargo Lde, L., Ribela, M.T., Keeble, J.E., Costa, S.K., Muscará, M.N. (2009). Participation of peripheral tachykinin NK receptors in the carrageenan -induced inflammation of the rat
temporo-mandibular joint.Eur J Pain13, 812–819.
Ferreira, S.H., Duarte, I.D.G., Lorenzetti, B.B. (1991). The molecular mechanism of action of peripheral morphine analgesia: Stimulation of
the cGMP system via nitric oxide release.Eur J Pharmacol201, 121–122.
Gadó, K., Gigler, G. (1991). Zymosan inflammation: A new method
suit-able for evaluating new antiinflammatory drugs.Agents Actions32, 119–
121.
Gegout, P., Gillet, P., Chevrier, D., Guingamp, C., Terlain, B., Netter, P. (1994). Characterization of zymosan-induced arthritis in the rat: Effects
on joint inflammation and cartilage metabolism. Life Sci 55, 321–
326.
Grangeiro, N.M.G., Aguiar, J.A., Chaves, H.V., Silva, A.A.R., Lima, V., Benevides, N.M.B., Brito, G.A.C., da Graça, J.R.V., Bezerra, M.M. (2011). Heme oxygenase/carbon monoxide-biliverdin pathway may be involved in the antinociceptive activity of etoricoxib, a selective COX-2
inhibitor.Pharmacol Rep63, 112–119.
Jang, D.S., Park, E.J., Kang, Y.H., Hawthorne, M.E., Vigo, J.S., Graham, J.G., Cabieses, F., Fong, H.H., Mehta, R.G., Pezzuto, J.M., Kinghorn, A.D. (2003). Potential cancer chemopreventive flavonoids from the
stems ofTephrosia toxicaria.J Nat Prod66, 1166–1170.
Keystone, E.C., Schorlemmer, H.U., Pope, C., Allison, A.C. (1977). Zymosan-induced arthritis: A model of chronic proliferative arthritis
following activation of the alternative pathway of complement.Arthritis
Rheum20, 1396–1401.
Khan, N., Sharma, S., Alam, A., Saleem, M., Sultana, S. (2001).Tephrosia
purpurea ameliorates N-diethylnitrosamine and potassium
bromate-mediated renal oxidative stress and toxicity in Wistar rats.Pharmacol
Toxicol88, 294–299.
Khatri, A., Garg, A., Agrawal, S.S. (2009). Evaluation of hepatoprotective
activity of aerial parts ofTephrosia purpureaL. and stem bark ofTecomella
undulata.J Ethnopharmacol122, 1–5.
Lee, H., Oh, S., Woo, J.K., Kim, W., Pelt, C.S.V., Price, R.E., Cody, D., Tran, H., Pezzuto, J.M., Moriarty, M., Hong, W.K. (2005). Chemopreventive effects of deguelin, a novel Akt inhibitor on tobacco-induced lung
Lodhi, S., Pawar, R.S., Jain, A.P., Singhai, A.K. (2006). Wound healing
potential ofTephrosia purpurea(Linn.) Pers. in rats.J Ethnopharmacol
108, 204–210.
Majewska, M., Zajc, K., Dulak, J., Szczepanik, M. (2008). Heme oxy-genase (HO-1) is involved in the negative regulation of contact
sensi-tivity reaction.Pharmacol Rep60, 933–940.
Martinez, R.M., Zarpelon, A.C., Zimermann, V.V.M., Georgetti, S.R., Baracat, M.M., Fonseca, M.J.V., Vicentini, F.T.M.C., Moreira, I.C.,
Andrei, C.C., Verri, W.A. Jr., Casagrande, R. (2012).Tephrosia sinapou
extract reduces inflammatory leukocyte recruitment in mice: Effect on
oxidative stress, nitric oxide and cytokine production.Rev Bras
Farma-cogn22, 587–597.
Moretti, C., Grenand, P. (1982). The ‘nivrées’, or ichthyotoxic plants of
French Guyana.J Ethnopharmacol6, 139–160.
Morita, T., Perrella, M.A., Lee, M.E., Kourembanas, S. (1995). Smooth muscle cell-derived carbon monoxide is a regulator of vascular cGMP. Proc Natl Acad Sci U S A92, 1475–1479.
Okonkwo, J.E., Iyadi, K.C., Effiong, C.O. (2004). Effect of chronic
admin-istration of haematological parameters of rats.Niger J Physiol Sci19,
10–13.
Rocha, F.A.C., Aragao, A.G.M. Jr., Oliveira, R.C., Pompeu, M.M.L., Vale, M.R., Ribeiro, R.A. (1999). Periarthritis promotes gait disturbance in
zymosan-induced arthritis in rats.Inflamm Res48, 485–490.
Soares, M.P., Seldon, M.P., Gregoire, I.P., Vassilevskaia, T., Berberat, P.O., Yu, J., Tsui, T.Y., Bach, F.H. (2004). Heme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell
acti-vation.J Immunol172, 3553–3563.
Tarus, P.K., Machocho, A.K., Lanmgat-Thoruwa, C.C., Chabra, S.C.
(2002). Flavonoids from Tephrosia aequilata.Phytochemistry 60, 375–
379.
Udeani, G.O., Gerhaüuser, C., Thomas, C.F., Moon, R.C., Kosmeder, J.W., Ksinghorn, A.D., Moriarty, R.M., Pezzuto, J.M. (1997). Cancer chemo-preventive activity mediated by deguelin, a naturally occurring
rote-noid.Cancer Res57, 3424–3428.
Vanderlei, E.S.O., de Araújo, I.W.F., Quinderé, A.L.G., Pontes, B.P., Eloy, Y.R.G., Rodrigues, J.A.G., Silva, A.A.R., Chaves, H.V., Jorge, R.J.B., Menezes, D.B., Evangelista, J.S.A.M., Bezerra, M.M., Benevides, N.M.B. (2011). The involvement of the HO-1 pathway in the anti-inflammatory action of a sulfated polysaccharide isolated from the red
seaweedGracilaria birdiae.Inflamm Res60, 1121–1130.
Vane, J.R., Warner, T.D. (2000). Nomenclature for COX-2 inhibitors. Lancet356, 1373–1374.
Vasconcelos, J.N., Lima, J.Q., Lemos, T.L.G., Oliveira, M.C.F., Almeida, M.M.B., Andrade-Neto, M., Mafezoli, J., Arriaga, A.M.C. (2009).
Estudo químico deTephrosia toxicariaPers.Quim Nova32, 382–386.
Vasconcelos, J.N., Santiago, G.M.P., Lima, J.Q., Mafezoli, J., Lemos, T.L.G., Silva, F.R.L., Lima, M.A.S., Pimenta, A.T.A., Braz-Filho, R., Arriaga,
A.M.C., Cesarin-Sobrinho, D. (2012). Rotenoids fromTephrosia toxicaria
Pers. with larvicidal activity againstAedes aegypti, the main vector of
dengue fever.Quim Nova35, 1097–1100.
Vicente, A.M., Guillen, M.I., Habib, A., Alcaraz, M.J. (2003). Beneficial effects of heme oxygenase-1 up-regulation in the development of
experimental inflammation induced by zymosan.J Pharmacol Exp Ther