UNIVERSIDADE FEDERAL DE SÃO CARLOS
CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA E EVOLUÇÃO
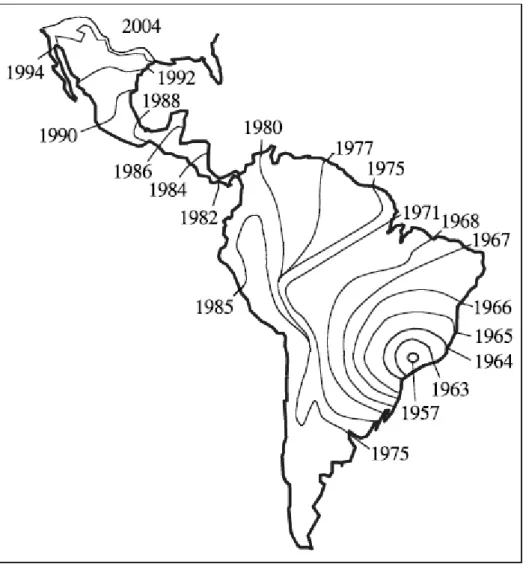
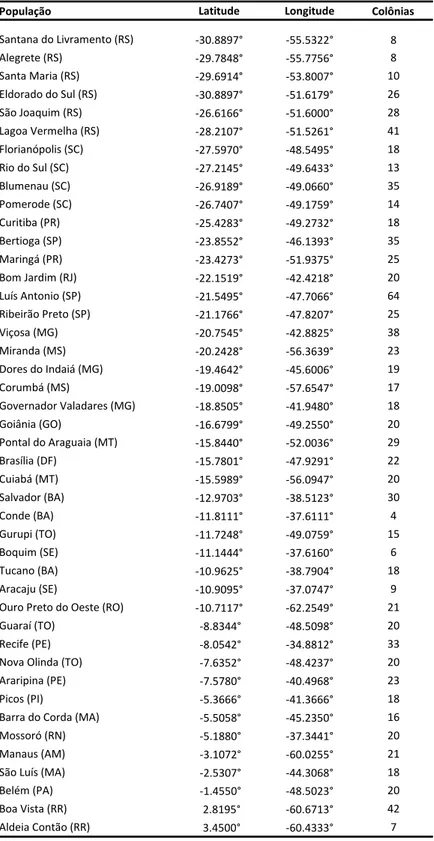
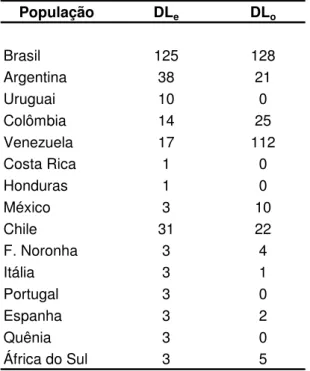
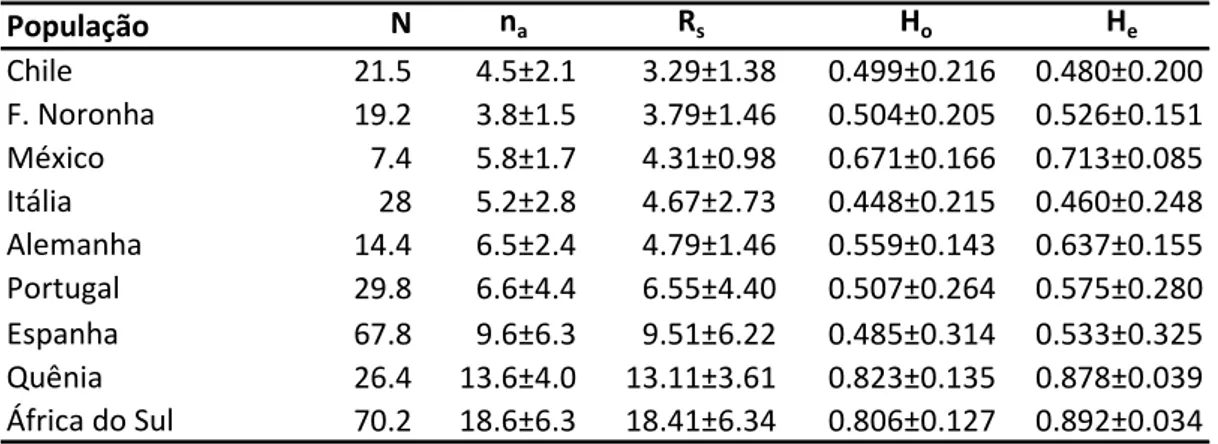
ESTRUTURA GENÉTICA DAS POPULAÇÕES DE ABELHAS AFRICANIZADAS
(
Apis mellifera
L.) E SUAS RELAÇÕES COM AS RAÇAS PARENTAIS
DETERMINADAS POR MEIO DA ANÁLISE DE MICROSSATÉLITES
THAÍS COLLET
UNIVERSIDADE FEDERAL DE SÃO CARLOS
CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA E EVOLUÇÃO
ESTRUTURA GENÉTICA DAS POPULAÇÕES DE ABELHAS AFRICANIZADAS
(
Apis mellifera
L.) E SUAS RELAÇÕES COM AS RAÇAS PARENTAIS
DETERMINADAS POR MEIO DA ANÁLISE DE MICROSSATÉLITES
THAÍS COLLET
Tese de Doutorado apresentada ao
Programa de Pós-Graduação em Genética
e Evolução do Centro de Ciências
Biológicas e da Saúde da Universidade
Federal de São Carlos, como parte dos
requisitos para a obtenção do título de
Doutor em Genética e Evolução, área de
concentração: Genética e Evolução.
Ficha catalográfica elaborada pelo DePT da
Biblioteca Comunitária/UFSCar
C698ep
Collet, Thaís.
Estrutura genética das populações de abelhas
africanizadas (
Apis mellifera
L.) e suas relações com as
raças parentais determinadas por meio da análise de
microssatélites / Thaís Collet. -- São Carlos : UFSCar, 2010.
64 f.
Tese (Doutorado) -- Universidade Federal de São Carlos,
2009.
1. Genética de populações. 2. Abelha - africanizada. 3.
Microssatélites. 4. Estrutura populacional. I. Título.
UNIVERSIDADE FEDERAL DE SAO CARLOS
CENTRO DE
CI~NCIAS
BIOL~GICAS
E
DA
SAODE
PROGRAMA DE
P ~ S - G R A D U A ~ ~
EM
GEN~TICA
E EVOLUGm
. . .
- A . * A . -*T...,,,...
,-,.: