Antimicrobial resistance of the upper respiratory tract commensal microbiota in bottlenose dolphins (Tursiops truncatus), under human care
Texto
Imagem
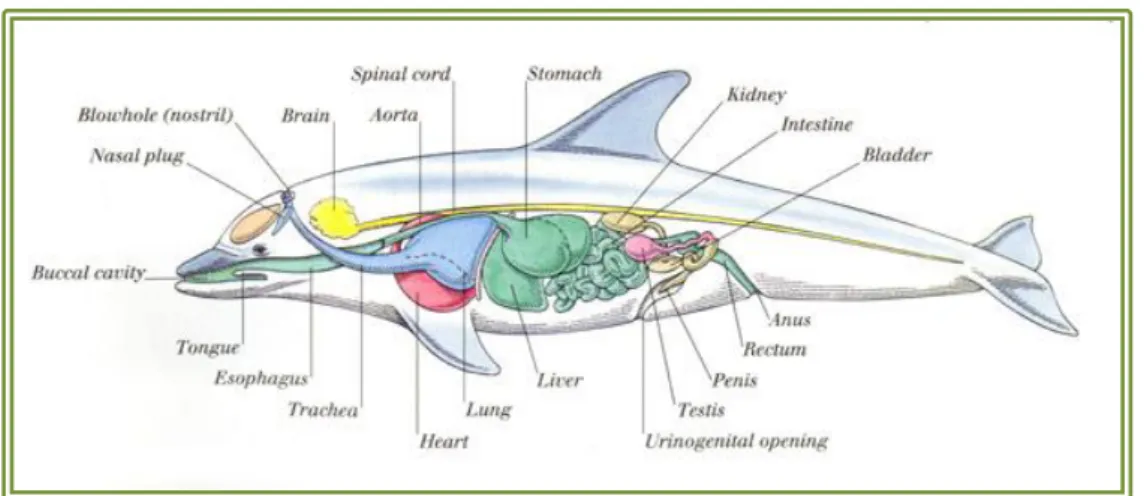
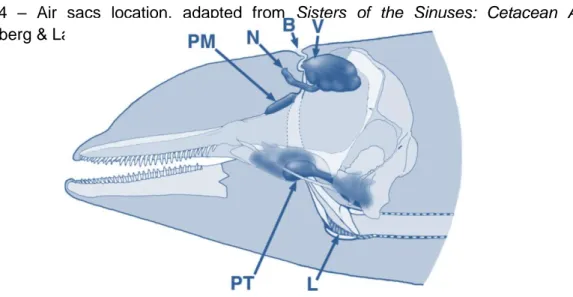
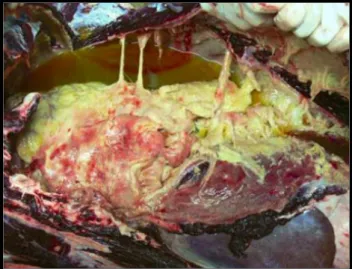

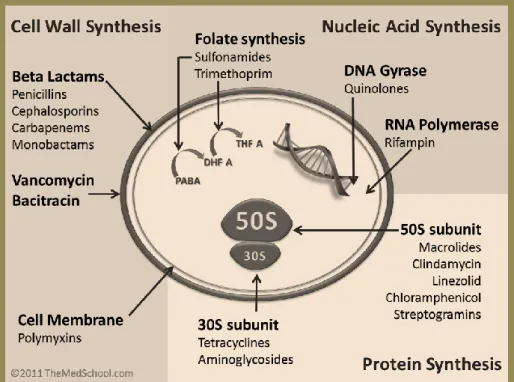
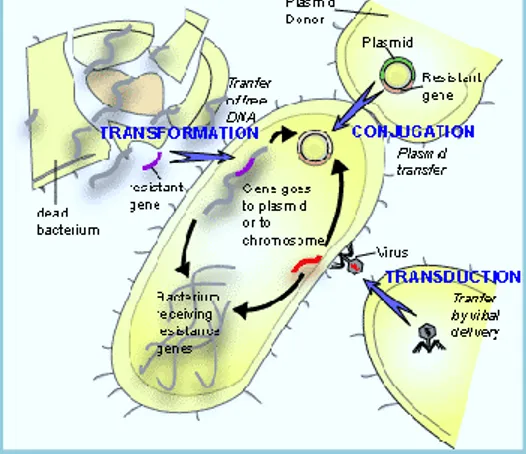
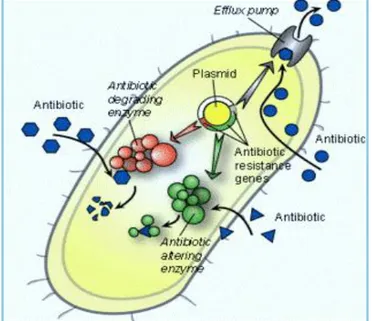
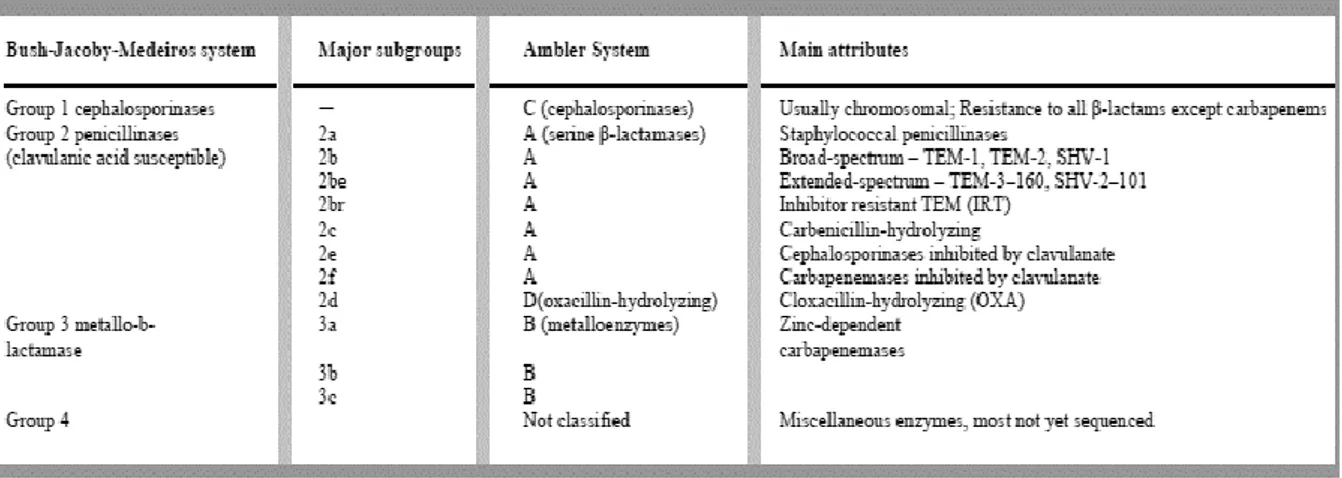


Documentos relacionados
Através dos resultados do presente estudo, observa-se que os benefícios do erfs conseguem ir mais além, demonstra n d o ser superior nos avanços de ganho de força se comparado
“Grupo de Interesse sobre Museologia (ICOFOM) As discussões do grupo tiveram como perspectiva a reflexão sobre aspectos teórico-metodológicos da Museologia (de acordo com
Além de assegurar a distribuição dos medicamentos, o farmacêutico hospitalar desempenha outras atividades, nomeadamente, fazer parte de várias comissões técnicas
Na realização da presente revisão sistemática, foram considerados somente os estudos que trataram do uso de macrófitas aquáticas no tratamento de
Gordon dirige uma grande responsabilidade aos educadores/professores e pais como orientadores do desenvolvimento da criança quanto a aprendizagens de música e através de música
A figura 1 mostra os percentuais de respostas da 1ª questão “Microrganismos são seres:” os resultados coletados mostram que de 327 pessoas, apenas 43 alunos (13,43%) (coluna A)
Observando a carta de controlo podemos verificar que existem dois pontos que se encontram fora do limite de controlo superior, ou seja o número de defeituosos daquela amostra
Formularam-se as seguintes questões de investigação: Quais as dificuldades iniciais que os alunos revelam na compreensão do conceito de função?; Que compreensão revelam os
