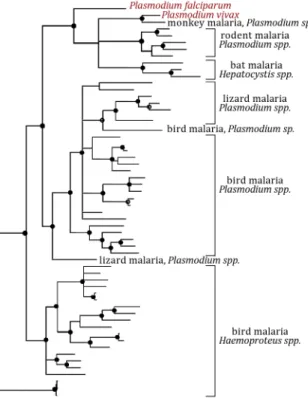
Interactions of Plasmodium blood and liver stages within a single host
187
0
0
Texto
(12)
(13)
(14)
(15)
(16)(17)
(18)(19)
(28)
(30)
(33)
(35)
(36)
(38)
Imagem


Documentos relacionados
Relativamente aos serviços centrais e considerando as diretrizes vindas da casa mãe, propõe-se a elaboração de um plano estratégico, sendo este posteriormente divulgado
The objective of the problem is to determine the optimal fleet size with the minimum total distribution cost, which considers fixed and variable routing costs and direct delivery
A ideia de realizar este projeto de investigação partiu da atual polémica de restruturação da Colina de Santana e o consequente destino dos Hospitais de São José, Santa
Redes de saúde são estruturas organizadas para o desenvolvimento das ações e serviços de saúde que buscam garantir a integralidade do cuidado. Este estudo discorre sobre a rede
A inclusão dos Estudantes com Necessidades Educacionais Especiais é vivenciada nas escolas por meio do enfrentamento de desafios que são próprios da complexidade e da subjetividade
Teve-se como objetivo central auxiliar os alunos na compreensão dos fundamentos da porcentagem e dando ênfase em situações cotidianas colocando-se em prática
Infection rate, infective mosquitoes, larvae melanization and annual transmission potential (ATP) of mosquitoes infected with Dirofilaria immitis , captured in Itacoatiara, State
Given the high prevalence of HBV markers in the different regions of the state of Santa Catarina, the high risk of transmission of HBV infection through transfusions and the high
