Contents lists available atScienceDirect
Biomedicine & Pharmacotherapy
journal homepage:www.elsevier.com/locate/biopha
The anticonvulsant e
ff
ect of a polysaccharide-rich extract from
Genipa
americana
leaves is mediated by GABA receptor
Dayanne Terra Tenório Nonato
a, Silvânia Maria Mendes Vasconcelos
c,
Mário Rogério Lima Mota
b, Paulo Goberlânio de Barros Silva
b, Arcelina Pacheco Cunha
d,
Nágila Maria Pontes Silva Ricardo
d, Maria Gonçalves Pereira
a,e, Ana Maria Sampaio Assreuy
a,
Edna Maria Camelo Chaves
a,⁎aSuperior Institute of Biomedical Sciences, State University of Ceara, Av. Dr. Silas Munguba, 1700, Itaperi, 60714-903, Fortaleza, CE, Brazil
bDepartment of Oral Pathology and Clinical Stomatology of Federal University of Ceara, Rua Coronel Nunes Valente, 1127, Rodolfo Teófilo, 607430-970, Fortaleza, CE,
Brazil
cDepartament of Physiology and Pharmacology, Federal University of Ceara, Rua Coronel Nunes Valente, 1127, Rodolfo Teófilo, 607430-970, Fortaleza, CE, Brazil dDepartment of Organic and Inorganic Chemistry, Federal University of Ceara, Rua Humberto Monte, S/N, Campus de PICI, 60440554, Fortaleza, CE, Brazil eFaculty of Education Science and Letters of the Hinterland, Rua José de Queiroz Pessoa, 2554
–Planalto Universitário, 63.900-000, Quixadá, CE, Brazil
A R T I C L E I N F O
Keywords: Plant polysaccharide “Jenipapo” Seizures Black neurons Brain oxidative stress
A B S T R A C T
Background: This study aimed to chemically characterize a polysaccharide-rich extract (PRE) obtained from Genipa americanaleaves and evaluate its neuroprotective effect in the brain morphology and oxidative markers using mice behavioral models.
Methods:Dry powder (5 g) ofG. americanaleaves were submitted to depigmentation in methanol. PRE was obtained by extraction in NaOH and precipitation with absolute ethanol and characterized by infrared spec-troscopy (FTIR) and nuclear magnetic resonance (1H and13C NMR). Swiss mice (25–35 g) received saline (0.9% NaCl) or PRE (1–27 mg/kg) by intraperitoneal (i.p.) route, 30 min before evaluation in behavioral models (open
field, elevated plus maze, sleeping time, tail suspension, forced swimming, seizures induced by pentylenete-trazole-PTZ). Animal’s brain were dissected and analyzed for histological alterations and oxidative stress. Results:FTIR spectrum showed bands around 3417 cm−1and 2928 cm−1, relative to the vibrational stretching of OeH and CeH, respectively.1H NMR spectrum revealed signals atδ3.85 (methoxyl groups) andδ2.4 (acetyl) ppm.13C NMR spectrum revealed signals atδ108.0 andδ61.5 ppm, corresponding to C1 and C5 ofα -L-ara-binofuranosyl residues. PRE presented central inhibitory effect, increasing the latency for PTZ-induced seizures by 63% (9 mg/kg) and 55% (27 mg/kg), and the latency to death by 73% (9 mg/kg) and 72% (27 mg/kg). Both effects were reversed by the association withflumazenil.
Conclusions:PRE, containing a heteropolysaccharide, presents antioxidant and anticonvulsant effect in the model of PTZ-induced seizures via gamma-aminobutyric acid (GABA), decreasing the number of hippocampal black neurons.
1. Introduction
Epilepsy is the second most common neurological disorder after stroke, affecting from 0.5% to 1% individuals at all ages of the world population [1]. The progression of epilepsy underlies a sequential cascade of events that includes cognitive impairment, accompanied by abnormal behavior and generation of reactive oxygen species in the
brain, considered one of the leading causes of generalized epilepsy as-sociated with recurrent seizures [2]. Furthermore, formation of dark neurons and morphological changes may occur depending on duration and intensity of seizures [3].
Pentylenetetrazole (PTZ) is widely accepted as an experimental animal model for the investigation of epileptogenesis and also to test the effectiveness of antiepileptic drugs [4]. Administration of PTZ by
https://doi.org/10.1016/j.biopha.2018.02.074
Received 9 January 2018; Received in revised form 31 January 2018; Accepted 19 February 2018
⁎Corresponding author.
E-mail addresses:[email protected](D.T.T. Nonato),[email protected](S.M.M. Vasconcelos),[email protected](M.R.L. Mota),
[email protected](P.G. de Barros Silva),[email protected](A.P. Cunha),[email protected](N.M.P.S. Ricardo),
[email protected](M.G. Pereira),[email protected](A.M.S. Assreuy),[email protected](E.M.C. Chaves).
Abbreviations:NaOH, sodium hydroxide; PTZ, pentylenetetrazole; PRE, polysaccharide-rich extract; NMR, nuclear magnetic resonance; GABA, gamma-aminobutyric acid; H, hydrogen; C, carbon; FTIR, infrared spectroscopy
0753-3322/ © 2018 Elsevier Masson SAS. All rights reserved.
intravenous or intraperitoneal routes enhances excitatory neuronal re-sponses in the central nervous system via blockade of the inhibitory responses of gamma amino butyric acid (GABA) [5].
The current pharmacological treatment of epilepsy often fails, being in most cases palliative, achieving the effective symptoms relief in about two-third of patients. Additionaly, numerous adverse effects are manifested, such as drowsiness, amnesia, tolerance and physical de-pendence [6]. In this context, biomolecules of plant origin could be considered as an alternative therapy.
The literature has been reported neuroprotective effects of plant polysaccharide extracts, such as antidepressant [7], antioxidant [8], anticonvulsant [9], also in cognitive impairment [10] and cerebral is-chemic injury [11].
Genipa americana L. (Rubiaceae), “jenipapo” or “jenipapeiro”, is widely distributed in Brazil, being its macerated leaves traditionally used by some native tribes to treat fever [12]. Phytochemical analysis of the extract fromG. america leaves revealed the presence offl avo-noids, among others [13], compounds recognized for their antioxidant effects [14,15].
The objective of this study was to chemically characterize the polysaccharide-rich extract obtained from G. americana leaves and evaluate its neuroprotective effects in the brain morphology and oxi-dative markers using mice behavioral models.
2. Materials and methods
2.1. Collection of G. americana leaves and polysaccharide extraction
Leaves were collected at Custodio-Quixada, Ceara (Brazil) and a voucher specimen (n° 46794) was deposited in the Herbarium Prisco Bezerra of Federal University of Ceará, Brazil. For polysaccharide ex-traction, leaves were dried at 40 °C and grounded into powder (5 g), which was suspended in absolute methanol (1:50, w/v), homogenized (2 h, 70 °C) andfiltered (procedure repeated 2×) for partial remotion of methanol-soluble material. Residue was suspended at 0.1 M NaOH (1:50 w/v), homogenized (2 h, 97 °C) and centrifuged (5421 ×g, 30 min, 25 °C) (procedure repeated 3×). Supernatant S1 (highly pig-mented) was discarded and supernatants S2 and S3 were pooled, neu-tralized with 1 M HCl, precipitated with four volumes of 96% ethanol (24 h, 4 °C) and centrifuged (5241 ×g, 30 min, 25 °C). The pellet was dialyzed against distilled water for 48 h, re-centrifuged and the lyo-philized supernatant, containing 54.6% total carbohydrates (including 21.1% uronic acid) and 12% protein [16] was named PRE (poly-saccharide-rich extract; 6.5% yield).
2.2. Polysaccharide chemical characterization
Infrared spectra (FTIR) was carried out on IR Prestige-21- Shimadzu spectrophotometer in the wave region of 4000–400 cm−1, being the polysaccharide sample mixed and grounded with KBr (1:80). The spectrum was obtained from 120 scans at 4 cm−1. The Nuclear Magnetic Resonance (NMR) spectra was obtained in a Brucker Avance-DRX 500 spectrometer equipped with an inverse detection probe and z-gradient accessory working at 499.9 (1H) and 125 MHz (13C), respec-tively. Approximately 30 mg of PRE were dissolved in 800μl D2O and sonicated for 20 min. For dimensional 1H data, 120 transients were collected at 70 °C (2 s acquisition time, 20 ppm spectral window, 32 k data points).1H chemical shifts were referenced to residual D2O atδ 4.78 ppm. For the one-dimensional13C spectrum, 20 k transients were collected at 70 °C (1 s acquisition time, 130 ppm spectral window, 32 k data points).
2.3. Drugs
Thiopental, diazepam (DZP),flumazenil (FLU), imipramine (IMP) and pentylenetetrazole (PTZ) were obtained from Sigma-Aldrich (St.
Louis, MO, USA) and solubilized in sterile saline (0.9% NaCl).
2.4. Animals
Male Swiss mice (25–30 g) were provided by the Central Animal House from the Federal University of Ceará -Brazil, maintained with free access to food and water, at 22 ± 2 °C and 12 h light-dark cycle and allowed to adapt to the laboratory for at least 1 h before experi-ments. The experimental protocols were performed during the light phase of the cycle, following the ethical standards in Directive 86/609/ EEC,“European Convention for the Protection of Vertebrate Animals Used for Experimental and other Scientific Purposes”, 1986, in ac-cordance with the Brazilian College of Animal Experimentation (COBEA) and the Ethics Committee for Animal Use (CEUA/UECE N°. 2451142/2014).
2.5. Experimental design
Animals (8–10) received saline (0.9% NaCl) or PRE (1–27 mg/kg) by intraperitoneal (i.p.) route, 30 min before evaluation in behavioral models (openfield, elevated plus maze, sleeping time, tail suspension, forced swimming). After euthanasia, the entire brain was removed for histological analysis and dissected (prefrontal cortex -PFC, hippo-campus–HC, striatum -ST) for oxidative stress evaluation.
2.6. Openfield
Mice were individually placed in the open-field apparatus consisting of an acrylic box (30 × 30 × 15 cm) with the floor divided into 9 squares. The number of rectangles crossed with all paws (crossing) was counted during 6 min [17]. Animals received saline, PRE or the sedative drug diazepam (2 mg/kg; i.p.) 30 min before evaluation.
2.7. Elevated plus maze
Mice were placed in the center of the high-labyrinth test consisting of two opposing open arms (30 × 5 × 25 cm) and two closed arms (30 × 5 × 25 cm). The number of entries and the time spent on the open and closed arms were registered during 5 min. The increase in the parameters observed in the open arms reveals anxiolytic effect [18]. Animals received saline, PRE or the anxyolitic drug diazepam (1 mg/kg; i.p.) 30 min before evaluation.
2.8. Tail suspension
Mice were suspended 50 cm from the ground by a tapefixed at 1 cm from the tail tip and the immobility time was recorded during 5 min [19]. The decrease in immobility time reveals antidepressant activity. Animals received saline, PRE or the antidepressant drug imipramine (10 mg/kg; i.p.) 30 min before evaluation.
2.9. Forced swimming
Mice were placed individually in acrylic cylinders (35 cm × 24 cm) containing 13.5 cm of water for observation of immobility time during 5 min. Immobility is considered when the animals perform minimal movements necessary to keep his head out of the water [20]. The de-crease in the immobility time reveals antidepressant effect. Animals received saline, PRE or the antidepressant drug imipramine (10 mg/kg; i.p.) 30 min before evaluation.
2.10. Sleep time
Mice received thiopental sodium (40 mg/kg; i.p.) and were left to sleep for observation of the sleep time (the corresponding time between righting reflex loss and recovery) and latency time [21]. The loss of
righting reflex is manifested by the animal inability to return to the prone position when placed in supine position for three consecutive times [22]. Animals received saline or PRE 30 min before the hypnotic sodium pentobarbital at 40 mg/kg (i.p.).
2.11. Rota rod
Mice were placed individually in the Rota Rod apparatus (2.5 cm diameter, raised 25 cm from thefloor, rotation of 12 rpm) and observed during 1 min for evaluation of motor coordination and permanence time in the apparatus [23]. Animals received saline, PRE or the an-xyolitic drug diazepam (1 mg/kg; i.p.) 30 min before evaluation.
2.12. Seizures
Induced by pentylenetetrazole (85 mg/kg; i.p.) 30 min after animals treatment with saline, PRE or diazepam (1 mg/kg; i.p.). The latency to the initiation offirst seizure and the latency to death (anticonvulsant response) were evaluated during 20 min [4]. The involvement of GABA receptors in PRE effect was investigated by the animals treatment with the GABA antagonistflumazenil (2.5 mg/kg, i.p.) 15 min before PRE.
2.13. Histological analysis
After seizures, animals were euthanized and the brains analyzed for
the presence of black neurons, according to routine histological methods (embedded in paraffin and blocks cut into sections of 5μm thickness). For this, tissues were sectioned at the hippocampal coronal plane (bregma 23.30 mm). The total number of dark neurons was analyzed (10 sections/animal) in slides stained with hematoxylin-eosin (H&E). Thefirst of the analyzed sections in the tissue block was chosen randomly, in sequence plus nine sections (50μm between sections). Dark neurons were characterized by neuronal shrinkage, cytoplasmic hyperbasophilia, and nuclear pyknosis [24]. The mean number of neurons was calculated using the formula [25]:
=
N ΣQ
a/f.ΣP A
“ΣQ”is the sum of counted particles appeared in sections,“a/f”is the area associated with each frame,“ΣP”is sum of frame associated points hitting space.
2.14. Brain oxidative stress
After seizures, the homogenates of prefrontal cortex (PFC), hippo-campus (HC) and striatum (ST) were prepared in phosphate buffer so-dium (pH = 7.4) and used to quantify the levels of lipid peroxidation (MDA) by the method of TBARS at 535 nm [26], nitrite by the method of Griess at 540 nm [27], and reduced glutathione (GSH) by DTNB method at 412 nm [28].
2.15. Statistical analysis
The results were expressed as Mean ± Standard Error of the Mean (SEM) and analyzed by ANOVA followed by Newman-Keuls test. The level of significance was set for p < 0.05.
3. Results
3.1. FTIR spectrum of PRE and1H and13C NMR spectrum of PRE
The bands around 3417 cm−1 and 2928 cm−1 relative to the vi-bration stretching of OeH and CeH, respectively [29]. Asymmetric and
symmetrical stretching are observed at 1628 cm−1, corresponding to the C]O bond in carboxylates [30,31]. At 1410 cm−1are deformation
bands of low intensity corresponding to theeCH2bond from the
pyr-anose ring [32]. The bands between 1500 and 700 cm−1are related to the polysaccharides fingerprint [33]. The bands between 1315 and 1240 cm−1corresponding to C
eO group bonds of pyranose ring [34]
and those around 1013 cm−1, to the vibrational stretching of C
eOeC
and CeOeH glycosidic bonds, associated with sugars in the chain
[33,35]. The band at 882 cm−1corresponds toδC
eH inβconfi
gura-tion (Fig. 1A) [30].
The1H NMR spectra reveals two signals atδ3.85 andδ2.4 ppm, derived from methoxyl groups and acetyl groups, respectively, sug-gesting the existence of uronic acid binding in the polymeric chain [36,37]. Signals atδ1.73 andδ1.81 ppm were derived from methyl groups ofα-L-rhamnose [38] (Fig. 1B). The13C NMR spectrum of PRE
reveals signals atδ108.0 andδ61.5 ppm corresponding to C1 and C5 of
α-L-arabinofuranosyl residues with different linkages, respectively
[38,39]. Signals atδ112.0 andδ107.8 ppm were derived fromα-D -galactopyranosyl andα-(1–4)-β-O-Me_D-galactopyranosyl, respectively
[40]. Signal atδ17.0 ppm is ascribed toα-L-rhamnose residues [36,41].
The signal at δ102.3 ppm could be assigned to 4-β-glucopyranosyl units [42] (Fig. 1C). Other chemical displacements (δ) of PRE carbon (65.0–87.2 ppm) and protons (4.0–5.76 ppm) could be assigned to a sugar ring in polysaccharides.
3.2. PRE reduces exploratory activity and is devoid of anxiolytic-like effects
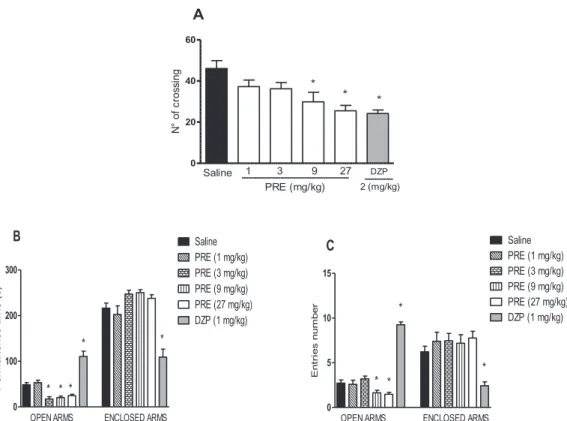
In the openfield test PRE reduced the exploratory activity, seen by the decrease in the number of crossing at 9 mg/kg (29.86 ± 4.67) and 27 mg/kg (25.50 ± 2.66), compared to saline (46.09 ± 3.81). Diazepam (2 mg/kg), used as control, reduced by 52% the number of crossing (Fig. 2A). In the plus-maze test, PRE reduced the time (s) spent in the open arms at all doses [17.44 ± 4.83 (3 mg/kg); 20.33 ± 3, 1 (9 mg/kg); 24.62 ± 3.20 (27 mg/kg) vs. saline (48.50 ± 4.96)] (Fig. 2B). Accordingly, PRE reduced the number of entries on the open arms at 9 mg/kg (1.40 ± 0.24) and 27 mg/kg (1.65 ± 0.20) com-pared to saline (2.90 ± 0.31) (Fig. 2C). Diazepam at 1 mg/kg, dose in which elicits anxiolytic effect, increased either the time (s) spent (110.60 ± 11.43) and the number of entries in the open arms (9.00 ± 0.327).
3.3. PRE increases the immobility time
In the tail suspension test, PRE increased the immobility time at the doses of 9 mg/kg (121.3 ± 16.25) and 27 mg/kg (111.0 ± 11.06) compared to saline (72.00 ± 4.95 s) (Fig. 3A). Similarly, PRE in-creased the immobility time in the forced swimming test at the doses of 3 mg/kg (56.88 ± 9.67), 9 mg/kg (59.60 ± 6.62) and 27 mg/kg (52.25 ± 9.69) compared to saline (28.25 ± 5.74 s) (Fig. 3B). Imi-pramine (10 mg/kg), used as antidepressant, did not alter this behavior.
3.4. PRE is devoid of hypnotic or sedative effects and does not alter motor coordination
Treatment with PRE did not alter the sleep latency [PRE (1 mg/ kg:2175 ± 19,43); PRE (9 mg/kg:2229 ± 4849; PRE (27 mg/ kg:1980 ± 23,75) vs. Saline:2900 ± 56,75] or the sleep duration [PRE (1 mg/kg:1485 ± 398,0); PRE (9 mg/kg:1920 ± 3716; PRE (27 mg/kg:2180 ± 247,4) vs. Saline:2093 ± 326,6] (data not show in
figure). The sleep inducer diazepam reduced the sleep latency (s) (105.6 ± 3.58 vs. saline: 290.0 ± 56.75) and increased the sleep duration (s) (6047 ± 286.1 vs. saline: 2093 ± 326.6). In the Rotarod test, PRE did not alter the animal’s permanence time on the apparatus at any doses [PRE (1 mg/kg:5586 ± 2,67); PRE (9 mg/kg:5200 ± 522; PRE (27 mg/kg:5743 ± 2,57) vs. Saline:5757 ± 2,42]. Diazepam reduced this time (35.43 ± 3.16 vs. Saline:57.57 ± 2.42 s) (data not show infigure).
3.5. PRE inhibits convulsion, the number of black neurons and oxidative stress in the model of seizures induced by PTZ
PRE increased the latency of PTZ-induced seizures at 9 mg/kg and 27 mg/kg by 63% and 55%, respectively compared to saline (65.0 ± 4.64 s). At these doses PRE also increased the latency to death by 73% (9 mg/kg) and 72% (27 mg/kg) compared to saline (154.0 ± 17.27 s). In addition, 20% of the treated animals survived. Flumazenil reversed the protective effects of PRE (27 mg/kg), reducing either the latencies to convulsion (57%) and to death (37%). The re-ference drug diazepam (1 mg/kg) enhanced by 70% animals surveil-lance (Table 1).
The histological evaluation of hippocampus showed that PRE re-duced by 58% the number of black neurons (20.17 ± 4.25) in animals that had been induced seizures by PTZ (47.29 ± 10.20) (Fig. 4A–D). Moreover the evaluation of oxidative stress markers after PRE treat-ment demonstrated reduction in MDA and increase in GSH content at all brain areas (pre-frontal cortex, hippocampus, striatum). However the nitrite content was unaltered by PRE (Table 2).
4. Discussion
This study identified a heteropolysaccharide in the polysaccharide-rich extract (PRE) obtained fromG. americana, that possesses central inhibitory effect and anticonvulsant activity, conferring brain protec-tion against oxidative stress and impairement in the number of hippo-campal black neurons.
The central inhibitory effect of PRE was demonstrated by several behavioral parameters, such as the reduced exploratory activity in the openfield, the lack of anxiolytic and antidepressant activities in the plus maze, tail suspension and forced swimming tests. In general, drugs possessing central inhibitory effects, such as anxiolytics (barbiturates and benzodiazepine), produce sedation as its major adverse effect [43]. In fact, ethanolic extracts of Ziziyphus nummularia[44] andPaederia scandens [45] showed central depressant action, accompanied by crease in the sleep duration. In contrast, PRE, despite of being in-hibitory, did not change the animal motor coordination or demonstrate hypnotic or sedative effect in the sleep time test.
Pharmacological studies support that either the reduction in in-hibitory synaptic activities via GABAergic mechanisms, or the increase in excitatory synaptic activities via glutamatergic mechanisms, would elicit seizures [2,5]. In our study, PRE protected PTZ-induced seizures by the increase in the latencies of seizure and death, showing 20% of animal survival. In addition,flumazenil (GABA receptor antagonist) reversed PRE protective effects, suggesting the involvement of GABA receptors in this protective effect. Other studies had demonstrated protective effect of aqueous extracts in the model of seizures induced by PTZ, such as those obtained from Feretia apodanthera [46], Galium spurium[47] andPaederia scandens[45].
Hippocampus is one of the most vulnerable brain area for lesions resultant from excitotoxicity [48] via NMDA and non-NMDA glutamate receptors (AMPA, kainate), and that the formation of black layer in hippocampal pyramidal neurons could be prevented by the blockage of glutamate receptors [49]. Black neurons, or apoptotic neurons, had been previously reported in models of ischemia, hypoglycaemia, ex-posure to excitatory amino acids and epilepsy, [50,51,52]. In our study a single dose of PTZ (85 mg/ kg) induced an increase in the number of black neurons in mice hippocampus, which was inhibited by PRE. Other studies had shown that PTZ administration at lower doses, either daily, alternate or consecutive days, stimulate black neurons development [53,54].
The production of free radicals is associated with damage of cellular structures in pathological conditions of the central nervous system, such as Parkinson's disease, stroke, dementia [55] and in epilepsy induced by PTZ [56,57]. PRE reduced the oxidative stress in the prefrontal cortex, hippocampus and striatum of animals that had been induced seizures by PTZ, seen by the increased levels of GSH and reduced levels of MDA. Similar results had been already described for other plant aqueous Fig. 2.PRE reduces exploratory activity and is devoid of anxiolytic-like effects. Mice received i.p. 0.9% NaCl (Saline), PRE or DZP 30 min before evaluation. (A) Openfield test (B,C); Plus Maze test. Mean ± SEM (n = 8). ANOVA and Newman-Keuls test.*p < 0.05 vs. Saline. PRE: polysaccharide-rich extract ofG. americana; DZP: diazepam.
Fig. 3.PRE increases immobility time. Mice received i.p. 0.9% NaCl (Saline), PRE or imipramine (10 mg/kg) 30 min before evaluation. (A) Tail suspension test; (B) Forced Swimming Test (FST). Mean ± SEM (n = 8). ANOVA and Newman-Keuls test.*p < 0.05 vs. Saline. PRE: polysaccharide-rich extract ofG. americana.
Table 1
PRE protects PTZ-induced seizures.
aTreatment
(mg/kg)
Seizure latency (s) Seizured animals (%)
death latency (s) survival animals (%)
Saline 65.0 ± 4.64 100 154.0 ± 17.27 0 PRE (1) 85.4 ± 9.48 100 340.8 ± 66.01 0 PRE (9) 175.0 ± 17.95* 100 582.4 ± 86.70* 20 PRE (27) 144.9 ± 18.50* 100 567.7 ± 88.89* 20 FLU + PRE
(27)
83.2 ± 9.29# 100 231.3 ± 59.16# 0
DZP (1) 122.5 ± 9.44* 100 1007 ± 98.98* 70
a30 min prior PTZ (85 mg/kg). Mean ± SEM (n = 8
–10). ANOVA and Newman-Keuls test.
* p < 0.05 vs. Saline.
extracts in the mice brain in the model of seizures induced by PTZ [46] and in a model of reperfusion ischemia [8]. The results presented in this study, together with preliminary data demonstrating the lack of toxicity of PRE, open perspectives for its therapeutic use in seizures. In con-clusion PRE contains a heteropolysaccharide presenting antioxidant and anticonvulsant effect in the model of PTZ-induced seizures via GABA, decreasing the number of hippocampal black neurons.
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgments
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico–CNPq (grant n° 307291/ 2013-8) and Fundação Cearense de Apoio ao Desenvolvimento Científico e tecnológico (FUNCAP). Prof. Msc. Vaneicia Santos Gomes (Faculdade de Educação, Ciências e Letras do Sertão Central - FECLESC) for identification of the plant specimen. Assreuy AMS and Vasconcelos SMM are senior investigators of CNPq.
References
[1] E. Trinka, J. Hofler, A. Zerbs, Causes of status epilepticus, Epilepsia 4 (2010) 127–138.
[2] J. Mehla, K.H. Reeta, P. Gupta, Y.K. Gupta, Protective effect of curcumin against seizures and cognitive impairment in apentylenetetrazole-kindled epileptic rat model, Life Sci. 87 (2010) 596–603.
[3] P. Baracskay, Z. Szepesi, G. Orban, G. Juhasz, A. Czurko, Generalization of seizures parallels the formation of“dark”neurons in the hippocampus and pontine reticular formation after focal–cortical application of 4-aminopyridine (4-AP) in the rat, Brain Res. 1228 (2008) 217–228.
[4] R.S. Fisher, Animal models of epilepsies, Brain Res. Brain Res. Rev. 14 (1989) 245–278.
[5] T. Azanchi, H. Shafaroodi, J. Asgarpanah, Anticonvulsant activity ofCitrus aur-antiumblossom essential oil (Neroli): involvment of the GABAergic system, Nat. Prod. Commun. 11 (2014) 1615–1618.
[6] J. Bidwell, T. Khuwatsamrit, B. Askew, J.A. Ehrenberg, S. Helmers, Seizure re-porting technologies for epilepsy treatment: a review of clinical information needs and supporting technologies, Seizure 32 (2015) 109–117.
[7] J. Wang, S. Flaisher-Grinberg, S. Li, H. Liu, L. Sun, Y. Zhou, et al., Antidepressant-like effects of the active acidic polysaccharide portion of ginseng in mice, J. Ethnopharmacol. 132 (2010) 65–69.
[8] S. Ai, X. Fan, L. Fan, Q. Sun, Y. Liu, X. Tao, et al., Extraction and chemical char-acterization ofAngelica sinensispolysaccharides and its antioxidant activity, Carbohydr. Polym. 94 (2013) 731–736.
[9] M.L. Calabrò, D. Raneri, S. Tommasini, R. Ficarra, S. Alcaro, A. Gallelli, et al., Enantioselective recognition of 2,3-benzodiazepin-4-one derivatives with antic-onvulsant activity on several polysaccharide chiral stationary phases, J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 21 (2006) 56–62.
Fig. 4.PRE reduces the number of hippocampal black neurons. Mice received i. p. 0.9% NaCl or PRE (27 mg/kg) 30 min before PTZ (85 mg/kg). (A–C) Black neurons stained with hematoxylin and eosin (H &E) (40×). (D) Graphical representation. Mean ± SEM (n = 8). ANOVA and Newman-Keuls test.*p < 0.05 vs. Saline + PTZ (pentylenete-trazole). PRE: polysaccharide-rich extract of G. americana.
Table 2
PRE inhibits oxidative stress in brain areas of animals convulsioned by PTZ.
Brain areas Parameters Experimental groups
Salinea PRE 1a PRE 9a PRE 27a
mg/kg
Prefrontal cortex MDA 0.15 ± 0.01 655.9 ± 24.09 456.1 ± 56.44* 426.7 ± 65.91* GSH 393.0 ± 74.81 999.2 ± 113.0* 891.2 ± 140.2* 1341 ± 260.2* NO 0.12 ± 0.002 0.13 ± 0.007 0.13 ± 0.003 0.13 ± 0.008 Hippocampus MDA 679.1 ± 62.48 477.9 ± 55.86 499.5 ± 34.43* 435.4 ± 43.56*
GSH 251.7 ± 30.36 1372 ± 194.4* 992.4 ± 140.6* 1128 ± 227.6* NO 0.15 ± 0.01 0.14 ± 0.01 0.17 ± 0.03 0.16 ± 0.01 Striatum MDA 630.2 ± 52.11 633.2 ± 47.61 402.3 ± 40.55* 427.7 ± 51.43*
GSH 437.2 ± 64.39 1053 ± 97.45* 931.1 ± 48.75* 1052 ± 155.5* NO 0.14 ± 0.007 0.18 ± 0.03 0.18 ± 0.01 0.14 ± 0.004
a30 min prior PTZ (85 mg/kg). Mean ± SEM (n = 8). ANOVA and Newman-Keuls test.
* p < 0.05 vs. Saline. PRE: polysaccharide-rich extract ofG. americana; PTZ: pentilenotetrazole.
[10] X. Lin, Z. Huang, X. Chen, Y. Rong, S. Zhang, Y. Jiao, et al., Protective effect of Millettia pulchrapolysaccharide on cognitive impairment induced by d-galactose in mice, Carbohydr. Polym. 101 (2014) 533–543.
[11] Z.Y. Zhou, Y.P. Tang, J. Xiang, P. Wua, H.M. Jin, Z. Wang, et al., Neuroprotective effects of water-solubleGanoderma lucidumpolysaccharides on cerebral ischemic injury in rats, J. Ethnopharmacol. 131 (2010) 154–164.
[12] P.G. Delprete, L.B. Smith, R.M. Klein, Flora Ilustrada Catarinense: Rubiáceas, Herbário Barbosa Rodrigues, (2005) Itajaí.
[13] J.S.F. Alves, L.A. Medeiros, M.F. Fernandez-Pedrosa, R.M. Araújo, S.M. Zucolotto, Iridoids from leaf extract ofGenipa americana, Rev. Bras. Farmacogn. 5 (2017) 641–644.
[14] B.H. Havsteen, The biochemistry and medical significance of theflavonoids, Pharmacol. Ther. 96 (2002) 67–202.
[15] S. Martin, R. Andriantsitohaina, Cellular mechanism of vasculo-protection induced by polyphenols on the endothelium, Annales de cardiologie et d’angéiologie 51 (2002) 304–315.
[16] R.O.S. Souza, P.L. Sousa, R.R. Paula, P.B. Menezes, T.L. Sampaio, L.D. Tessarolo, et al., Trypanocidal activity of polysaccharide extract fromGenipa americanaleaves, J. Ethnopharmacol. 10 (2018) 311–317.
[17] K.C. Montgomery, The relation between fear induced by novel stimulation and exploratory behavior, J. Comp. Physiol. Psychol. 8 (1955) 254–260. [18] R.G. Lister, The use of a plus-maze to measure anxiety in the mouse,
Psychopharmacology 92 (1987) 180–185.
[19] L. Steru, R. Chermat, B. Thierry, P. Simon, Tail suspension test: a new method for screening antidepressants in mice, Psychopharmacology 85 (1985) 367–370. [20] R.D. Porsolt, A. Bertini, M. Jalfre, Behavioral despair in mice: a primary screening
test for antidepressants, Arch. Int. Pharmacodyn. Ther. 229 (1977) 327–336. [21] R. Ferrini, G. Miragoli, B. Taccardi, Neuro-pharmacological studies on SB 5833, a
new psychotherapeutic agent of the benzodiazepine class, Arzneimittelforschung 12 (1974) 2029–2032.
[22] R. Mattei, R.F. Dias, E.B. Espínola, E.A. Carlini, S.B. Barros, Guarana
(Paulliniacupana): toxic behavioral effects in laboratory animals and antioxidant activity in vitro, J. Ethnopharmacol. 2 (1998) 111–116.
[23] N.W. Dunham, T.S. Miya, A note on a simple apparatus for detecting neurological deficit in rats and mice, J. Am. Pharm. Assoc. Am. Pharm. Assoc. 46 (1957) 208–209.
[24] A.V. Zaitsev, Morphofunctional changes infield CA1 of the rat hippocampus after pentylenetetrazole and lithium-pilocarpine induced seizures, Zh. Evol. Biokhim. Fiziol. 6 (2014) 463–469.
[25] F. Karimzadeh, M. Hosseini, D. Mangeng, H. Alavi, G.R. Hassanzadeh, M. Bayat, M. Jafarian, H. Kazemi, A. Gorji, Anticonvulsant and neuroprotective effects of Pimpinella anisum in rat brain, BMC Complement. Altern. Med. 12 (2012) 76. [26] N.T. Huong, K. Matsumoto, R. Kasai, K. Yamasaki, H. Watanabe, In vitro anti-oxidant activity of vietnamese ginseng saponin and its components, Biol. Pharm. Bull. 21 (1998) 978–981.
[27] L.C. Green, S.R. Tannenbaum, P. Goldman, Nitrate synthesis in the germfree and conventional rat, Science 212 (1981) 56–58.
[28] J. Sedlak, R.H. Lindsay, Estimation of total, protein-bound, and nonprotein sulf-hydryl groups in tissue with Ellman’s reagent, Anal. Biochem. 25 (1968) 192–205. [29] H.H. Sun, W.J. Mao, Y. Chen, S.D. Guo, H.Y. Li, X.H. Qi, Y.L. Chen, J. Xu, Isolation, chemical characteristics and antioxidant properties of the polysaccharides from marine fungusPenicilliumsp. F23-2, Carbohydr. Polym. 78 (2009) 117–124. [30] H. Cheng, S. Feng, X. Jia, Q. Li, Y. Zhou, C. Ding, Structural characterization and
antioxidant activities of polysaccharides extracted fromEpimedium acuminatum, Carbohydr. Polym. 92 (2013) 63–68.
[31] C. Zhao, L. Gao, C. Wang, B. Liu, Y. Jin, Z. Xing, Structural characterization and antiviral activity of a novel heteropolysaccharide isolated fromGrifola frondosa against enterovirus 71, Carbohydr. Polym. 144 (2016) 382–389.
[32] F.C.A. Buriti, K.M.O. Santos, V.G. Sombra, J.S. Maciel, M.A. Daniele,
D.M.A. Teixeira Sá, Characterisation of partially hydrolysed galactomannan from Caesalpinia pulcherrimaseeds as a potential dietaryfibre, Food Hydrocolloids 35 (2014) 512–521.
[33] D. Karnik, J. Jung, S. Hawking, L. Wicker, Sugar beet pectin fractionated using isopropanol differs in galacturonic acid, protein, ferulic acid and surface hydro-phobicity, Food Hydrocolloids 60 (2016) 179–185.
[34] L.C. Bichara, P.E. Alvarez, M.V.F. Bimbi, H. Vaca, H.C. Gervasi, S.A. Brandán, Structural and spectroscopic study of a pectin isolated from citrus peel by using FTIR and FT-Raman spectra and DFT calculations, Infrared Phys. Technol. 76 (2016) 315–327.
[35] F. Xu, K. Liao, Y. Wu, Q. Pan, L. Wu, H. Jiao, D. Guo, B. Li, B. Liu, Optimization
characterization, sulfation and antitumor activity of neutral polysaccharides from the fruit ofBorojoa sorbiliscuter, Carbohydr. Polym. 151 (2016) 364–372. [36] K. Alba, A.P. Laws, V. Kontogiorgos, Isolation and characterization of acetylated
LM-pectins extracted from okra pods, Food Hydrocolloids 43 (2015) 726–735. [37] Y. Tamaki, T. Konishi, M. Fukuta, M. Tako, Isolation and structural characterisation
of pectin from endocarp ofCitrus depressa, Food Chem. 107 (2008) 352–361. [38] F.F. Simas-Tosin, R.R. Barraza, M.D. Ferreira, M.F.P. Werner, C.H. Baggio,
E.R. Carbonero, G.L. Sassaki, M. iacomini, P.A.J. Gorin, Glucuronoarabinoxylan from coconut palm gum exudate: chemical structure and gastroprotective effect, Carbohydr. Polym. 107 (2014) 65–71.
[39] C.T. Scoparo, L.M. Souza, N. Dartora, G.L. Sassaki, A.P. Santana-Filho, M.F.P. Werner, D.G. Borato, C.H. Baggio, Chemical characterization of hetero-polysaccharides from green andblack teas (Camellia sinensis) and their anti-ulcer effect, Int. J. Biol. Macromol. 86 (2016) 772–781.
[40] W.Q. Li, S.S. Shao, L. He, J. Cheng, S. Han, Y. Liu, Structure and chain conformation of a neutral intracellular heteropolysaccharide from mycelium ofPaecilomyces ci-cadae, Carbohydr. Polym. 136 (2016) 728–737.
[41] C.K. Nandan, R. Sarkar, S.K. Bhanja, S. Mondal, S.S. Islam, Structural character-ization of a heteropolysaccharide isolated from the rhizomes ofCurcuma zedoaria (Sati), Carbohydr. Polym. 86 (2011) 1252–1259.
[42] Q. Guo, S.W. Cui, Q. Wang, X. Hu, J. Kang, R.Y. Yada, Structural characterization of a low-molecular-weight heteropolysaccharide (glucomannan) isolated from Artemisia sphaerocephalaKrasch, Carbohydr. Res. 350 (2012) 31–39. [43] A.H. Leão, A. Cabral, G.S. Izídio, A.M. Ribeiro, R.H. Silva, Diazepam effects on
aversive memory retrieval and extinction: role of anxiety levels, Pharmacol. Biochem. Behav. 141 (2015) 42–49.
[44] M. Goyal, D. Sasmal, CNS depressant and anticonvulsant activities of the alcoholic extract of leaves ofZiziyphus nummularia, J. Ethnopharmacol. 151 (2014) 536–542. [45] T. Yang, B. Kong, J.W. Gu, Y.Q. Kuang, L. Cheng, W.T. Yang, J.M. Cheng, Y. Ma,
X.K. Yang, Anticonvulsant and sedative effects of paederosidic acid isolated from Paederia scandens(Lour.) Merrill in mice and rats, Pharmacol. Biochem. Behav. 111 (2013) 97–101.
[46] G.S. Taiwe, F.C.O. Moto, E.R.M. Ayissi, G.T. Ngoupaye, J.S.K. Njapdounke, G.C.N. Nkantchoua, N. Kouemou, J.P.O. Omam, A.K. Kandeda, S. Pale, D. Pahaye, E. Ngo Bum, Effects of a lyophilized aqueous extract ofFeretia apodantheraDel. (Rubiaceae) on pentylenetetrazole-induced kindling, oxidative stress, and cognitive impairment in mice, Epilepsy Behav. 43 (2015) 100–108.
[47] N. Orhan, D.D. Orhan, M. Aslan, I.E. Orhan, UPLC–TOF-MS analysis ofGalium spuriumtowards its neuroprotective and anticonvulsant activities, J. Ethnopharmacol. 14 (2012) 220–227.
[48] W. Pulsinelli, Pathophysiology of acute ischaemic stroke, Lancet 29 (1992) 533–536.
[49] Z.S. Kherani, R.N. Auer, Pharmacologic analysis of the mechanism of dark neuron production in cerebral cortex, Acta Neuropathol. 116 (2008) 447–452. [50] E. Kövesdi, J. Pál, F. Gallyas, The fate of‘dark’neurons produced by transient focal
cerebral ischemia in a non-necrotic and non-excitotoxic environment: neurobiolo-gical aspects, Brain Res. 1147 (2007) 272–283.
[51] N. Auer, H. Kalimo, Y. Olsson, B.K. Siesjo, The temporal evolution of hypoglycemic brain damage. I. Light- and electronmicroscopicfindings in the rat cerebral cortex, Acta Neuropathol. 67 (1985) 13–24.
[52] R.S. Sloviter, D.W. Dempster,“Epileptic”brain damage is replicated qualitatively in the rat hippocampus by central injection of glutamate or aspartate but not by GABA or acetylcholine, Brain Res. Bull 15 (1985) 39–60.
[53] M.I. Naseer, I. Ullah, M.H. Al-Qahtani, S. Karim, N. Ullah, S.A. Ansari, M.O. Kim, F. Bibi, Decreased GABABR expression and increased neuronal cell death in de-veloping rat brain after PTZ-induced seizure, Neurol. Sci. 34 (2013) 497–503. [54] D.S. Vasil’ev, N.L. Tumanova, I.A. Zhuravin, K.K. Kim, Y.N. Lukomskaya,
L.G. Magazanik, A.V. Zaitsev, Morphofunctional changes infield CA1 of the rat hippocampus after pentylenetetrazole and lithium-pilocarpine induced seizures, Zh. Evol. Biokhim. Fiziol. 6 (2014) 463–469.
[55] R.M. Adibhatla, J.F. Hatcher, Altered lipid metabolism in brain injury and dis-orders, Subcell. Biochem. 49 (2008) 241–268.
[56] N. Patsoukis, G. Zervoudakis, C.D. Georgiou, F. Angelatou, N.A. Matsokis, N.T. Panagopoulos, Effect of pentylenetetrazol-induced epileptic seizure on thiol redox state in the mouse cerebral cortex, Epilepsy Res. 62 (2004) 65–74. [57] V. Bashkatova, V. Narkevich, G. Vitskova, A. Vanin, The influence of anticonvulsant