Toward a better understanding of the subtropical Atlantic Forest in the state of Santa Catarina, Brazil: tree sampling accuracy, species richness and frequency of rare species
Texto
Imagem

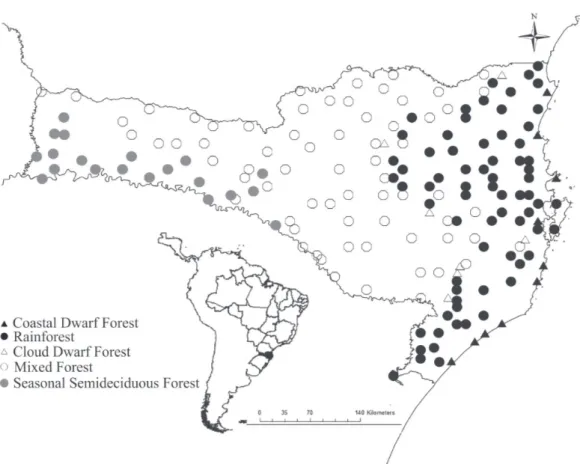



Documentos relacionados
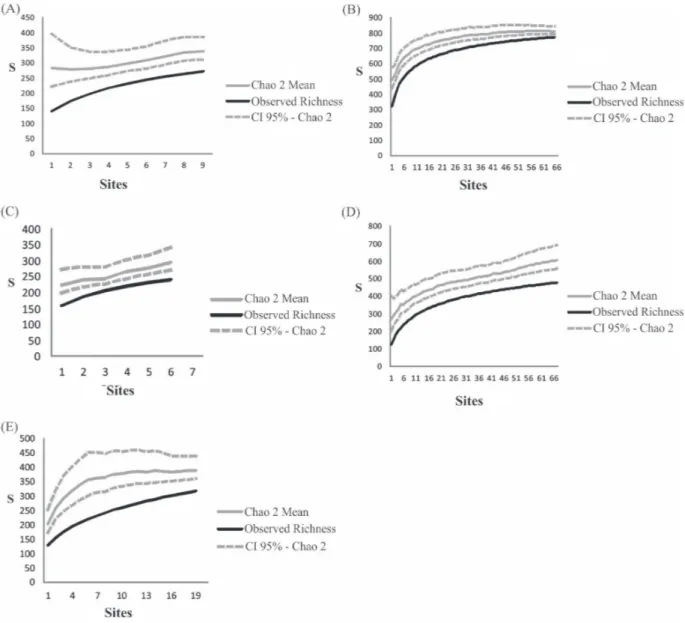
While the former analysis indicates the observed species accumulation curve, the latter is a sample-based nonparametric estimator for addressing expected species richness of
A cada nova edição de edital do Projeto Rondon, segue-se uma série de trabalhos para que a uma IES possa participar, entre estas atividades, destaca-se as principais
competindo já perdeu todos os mapas das festas. A participação em uma festa conseguimos com um corpo leve e desarmado.. Com a referida estrutura, trata- se, pois,
Este trabalho tem como objetivo traçar a trajetória profissional do arquiteto e urbanista Jorge Wilheim (1928-2014) e o papel que desempenhou no planejamento urbano de grandes
Os objetivos dos MP explicitam o eixo de contextualização forte, como por exemplo: - preparar quadros para a atuação estratégica na educação; ampliar a capacidade de
Sem efeito vinculante, mas igualmente importante, é a Declaração sobre o Direito e a Responsabilidade dos Indivíduos, Grupos ou Órgãos da Sociedade de Promover e
O farmacêutico, como gestor do medicamento, tem que conseguir gerir os aspetos administrativos, técnicos e económicos de forma a garantir a gestão do stock de medicamentos e de
No seu entender, o fato de estarem os depósitos vinculados a existência de um contrato de trabalho, impede caracterizá-los, constitucionalmente, com o imposto, taxa ou contribuição
