A new method to quantify and compare the multiple components of fitness-A study case with kelp niche partition by divergent microstage adaptations to Temperature
Texto
Imagem
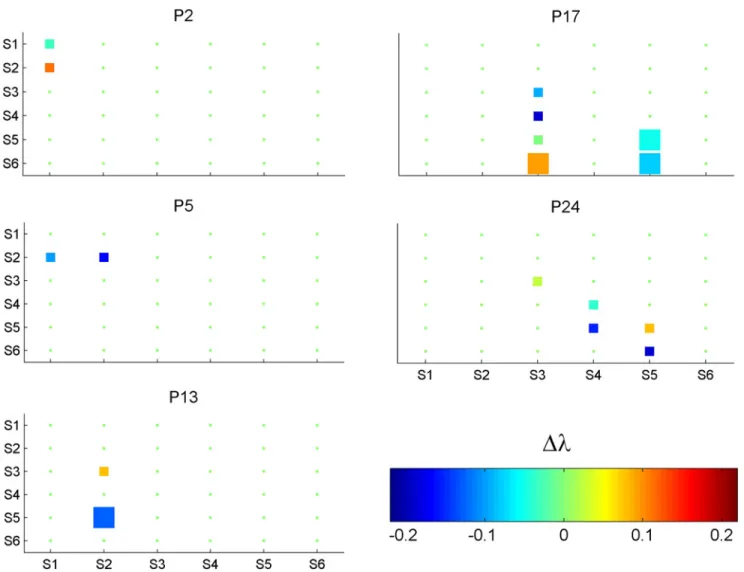

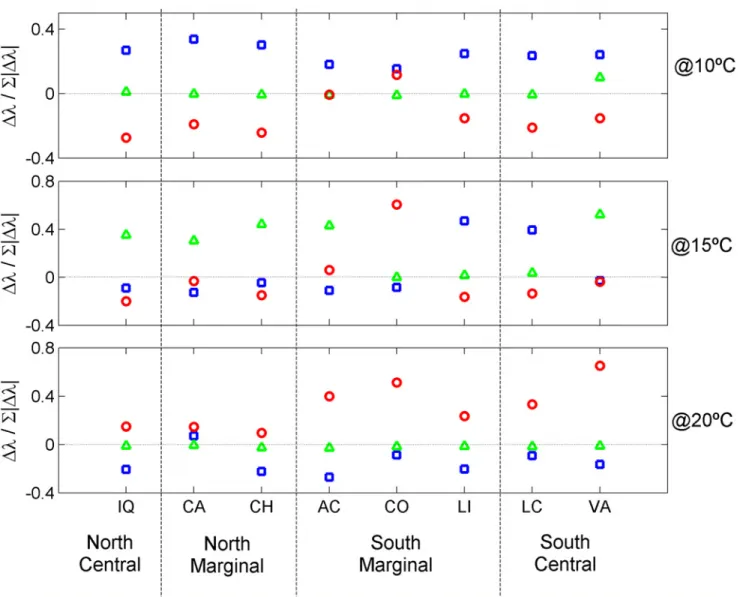
Documentos relacionados
[r]
Despercebido: não visto, não notado, não observado, ignorado.. Não me passou despercebido
[r]
Quando ligar um televisor Sony, a entrada pode ser seleccionada automaticamente carregando no botão de entrada após a configuração (Entrada SYNC) - avance para o passo 5.. 4
Provar que a interseção de todos os subcorpos de E que são corpos de decomposição para f sobre F é um corpo de raízes fe
The fourth generation of sinkholes is connected with the older Đulin ponor-Medvedica cave system and collects the water which appears deeper in the cave as permanent
A reconstruction of the past annual precipitation rates for the last 280 years has been obtained from speleothem annual growth rates, derived from the
O padrão de uso avaliativo da língua, por parte do par de audiodescritores, no roteiro de AD da peça quanto aos termos dos sistemas que compõem a subrede cuja condição de entrada
