Resposta ao estresse ácido em Saccharomyces
cerevisiae e efeito protetor do íon sódio na
morte celular induzida por ácido
AUTORA: GILZEANE DOS SANTOS SANT’ANA
ORIENTADOR: PROF. Dr. IESO DE MIRANDA CASTRO
Tese apresentada ao Programa de Pós-Graduação do Núcleo de Pesquisas em Ciências Biológicas da Universidade Federal de Ouro Preto, como parte integrante dos requisitos para a obtenção do Título de Doutor(a) em Ciências Biológicas, área de concentração: Biologia Molecular
II RESUMO
Saccharomyces boulardii é usada como probiótico para previnir ou tratar distúrbios
gastrointestinais ocasionados pelo uso excessivo de antibióticos, ou em enterites agudas. A fim de exercer os efeitos benéficos, os probióticos devem ter um mecanismo que os protegem do estresse do ambiente gástrico intestinal. Neste trabalho analisamos as diferenças metabólicas na resposta ao estresse ácido de S. boulardii comparativamente a S. cerevisiae W303, bem como a proteção conferida pela adição do íon sódio nesta condição.
Como esperado, S. boulardii exibiu maior tolerância ao meio gástrico simulado frente às
outras linhagens de S. cerevisiae. Sob estas condições o baixo pH (pH 2) foi o principal
fator responsável pela diminuição da viabilidade celular. Neste trabalho observamos que a adição de baixas concentrações de cloreto de sódio foi mais eficaz do que a adição de outros sais na proteção das células em condições de estresse ácido. O efeito protetor do Na+ na viabilidade de leveduras, sob condições ácidas, foi testado utilizando cepas de S. cerevisiae com deleções nos genes que codificam proteínas envolvidas na homeostase de
sódio, como por exemplo, os genes que codificam para a bomba Na+ATPase (ena1-4∆) e genes codificadores dos canais trocadores Na+/H+ (nha1∆) e Na+/H+ pré-vacuolar (nhx1∆). O efeito protetor de NaCl está relacionado à influência do íon Na+ no potencial elétrico de membrana citoplasmática. Além disso, a ausência ou baixa expressão do gene que codifica para a proteína Ena1-4p sugere uma relação direta com os níveis basais do potencial de membrana citoplasmática das células de leveduras. É provável que a resistência de S. boulardii e do mutante ena1-4∆ ao estresse ácido (na ausência ou presença de NaCl), deve-se ao elevado potencial basal da membrana destas células e ao fato do íon sódio retardar a despolarização da membrana induzida pelo pH 2.0. Além disso, o maior conteúdo de trealose observado em S. boulardii pode ter contribuído para a resposta mais eficiente
III
Saccharomyces boulardii as a probiotic is used to prevent or treat gastrointestinal disorders
caused by excessive use of antibiotics, or in acute enteritis. In order to exert beneficial effects, the probiotic must have a mechanism that protects the gastric intestinal stress
environment. We examined the differences in metabolic response to acid stress in
S. boulardii compared with S. cerevisiae W303 and the protection conferred by the addition
of sodium ion in this condition. As expected, S. boulardii exhibited higher tolerance to
simulated gastric environment facing the other strains of S. cerevisiae. Under these
conditions the low pH (pH 2) was the main factor responsible for the decrease in cell viability. In this work we observed that the addition of low concentrations of sodium chloride was more effective than the addition of other salts in the protection of cells under conditions of acid stress. The protective effect of Na + on the viability of yeast cells under acidic conditions, was tested using using S. cerevisiae null mutants for Na+-ATPases (ena 1-4 null mutant), for Na+/H+ antiporter (nha1∆) and Na+/H+ antiporter prevacuolar (nhx1∆).
The protective effect of NaCl relates with the effect of ion Na+ in electric membrane potential. Additionally, the absence or low expression/activity of Ena proteins seems to be closely related to basal membrane potential of yeast cells. Moreover, the absence or low expression of the gene coding for protein Ena1-4p suggests a direct relationship with the levels of membrane potential of yeast cells. It is likely that the resistance of S. boulardii
and ena1-4 mutant to acid stress (in the absence or presence of NaCl), due to the high
potential of the basal membrane of these cells and because of the ion slowing of membrane depolarization induced by pH 2.0. Furthermore, the highest concentration of trehalose observed in S. boulardii may have contributed to a more efficient response against the
IV
V
RESUMO II
ABSTRACT III
LISTA DE ABREVIAÇÕES X
LISTA DE TABELAS XIII
LISTA DE FIGURAS XIV
Introdução... 1
1 Introdução 2 1.1 A levedura do gênero Saccharomyces.... 2
1.1.1 Saccharomycescerevisiae... 2
1.1.2 Saccharomycesboulardii... 4
1.2 Mecanismos envolvidos em resposta a estresse em leveduras... 7
1.3 Mecanismo de resposta ao estresse ácido... 14
Objetivos... 18
2 Objetivos... 19
VI
2.2 Objetivos Específicos... 19
Material e métodos... 21
3 Material e métodos... 22
3.1 Cepas Saccharomyces utilizadas nos experimentos... 22
3.2 Meios de cultura e condições de cultivo... 23
3.3 Condições de estresse... 24
3.3.1 Teste de Viabilidade... 24
3.4 Reação em cadeia da polimerase em tempo real... 25
3.4.1 Extração e preparação de RNA total de levedura... 25
3.4.2 Obtenção do cDNA... 26
3.4.3 Reação em cadeia da polimerase em tempo real.………...……. 29
3.5 Medidas do pH interno... 29
3.6 Conteúdo intracelular de sódio e potássio... 30
3.7 Atividade H+-ATPásica... 32
3.7.1 Obtenção das membranas citoplasmáticas... 32
3.7.2 Ensaio da atividade H+-ATPásica... 33
3.7.3 Dosagem de proteína... 35
3.8 Determinação do potencial de membrana... 35
3.8.1 Aquisição e análise dos dados em citômetro de fluxo... 35
3.8.2 Curva padrão de potencial com células fixadas... 36
3.9 Medida de cálcio celular total... 37
3.10 Atividade Ca+2-ATPásica... 39
VII
3.11.1 Transformação de levedura... 41
3.11.1.1 Preparo de células de levedura competentes... 41
3.11.1.2 Transformação de levedura... 42
3.11.2 Concentração de cálcio citosólico livre em estresse ácido... 43
3.12 Capacidade tamponante de células e extrato celular...… 44
3.13 Análise Lipídica por Cromatografia Gasosa... 44
3.14 Dosagem de reservas energéticas... 45
3.14.1 Dosagem de trealose e glicogênio... 46
3.14.2 Dosagem de níveis de ATP... 47
3.15 Estatística…………...……… 47
Resultados...…...…...…... 48
4. Resultados... 49
4.1 Tolerância das cepas Saccharomyces ao suco gástrico e a seus componentes... 49
VIII
4.2.1 Variações do pH intracelular em cepas Saccharomyces selvagens e
com deleções nos genes que codificam proteínas envolvidas na
homeostase do sódio... 60
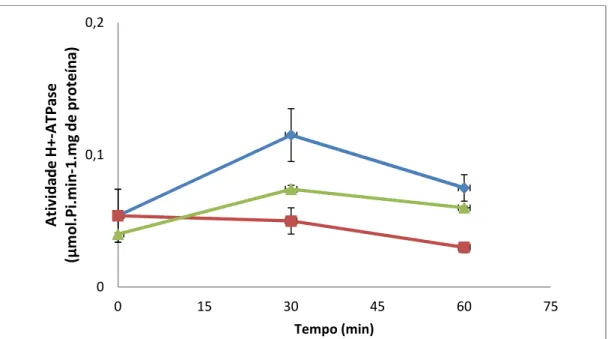
4.3 Participação da H+- ATPase de membrana plasmática em resposta ao estresse gástrico... 63
4.4 Potencial de membrana das células submetidas ao estresse ácido... 65
4.5 Análise de alterações metabólicas que podem esclarecer a maior resistência da cepa Saccharomyces boulardii sob o estresse ácido... 77
4.5.1 Avaliação da homeostase do cálcio em cepas Saccharomyces sob o estresse ácido... 67
4.5.2 Capacidade tamponante das cepas S. boulardii sob o estresse ácido ... 72
4.5.3 Análise qualitativa das alterações na composição lipídica de membrana das cepas Saccharomyces sob o estresse ácido... 74
4.5.4 Análise quantitativa das reservas energéticas durante a exposição ao estresse ácido... 76
Discussão... 79
5 Discussão... 80
Conclusões... 90
6 Conclusões... 91
IX
X
LISTA DE ABREVIAÇÕES
ACT1 Gene que codifica para a actina
ADP Adenosina 5´ bifosfato
AMPc Adesosina 5´ monofosfato cíclico
ANSA 1-Amino- 2- hidroxi- 4 - ácido naftaleno sulfônico
ATP Adenosina 5´ Trifosfato
cDNA DNA complementar
DAA Diarréia associada ao uso de antimicrobianos
DiBAC4(3) Bis-oxonol trimetina (1,3- ácido dibutilbarbitúrico)
DNA Ácido desoxirribonucléico
DNTPs Desoxinucleotídeos 5´ trifosfato
EDTA “Ethylene diamine tetraacetic acid” ou ácido etileno diamino
tetra-acético
EGTA “Ethylene glycol tetraacetic acid” ou ácido etileno glicol
tetra-acético
ENA1 Gene que codifica para a bomba de Na+K+ de membrana
citoplasmática Ena1p
Ena1p Bomba de Na+K+ de membrana citoplasmática
FDA Food and drug administration
GRAS Generally recognized as safe ou Geralmente reconhecido como
seguro
HEPES Ácido N-2-HidroxietilPiper, P.azina-N'-2'-Etanossulfônico
HSP Heat schock proteins ou proteínas de choque térmico
HSP70 Heat schock protein family 70 ou família 70 de proteínas de
XI
microbiana
NHA1 Gene que codifica para o canal antiporter de membrana
citoplasmática Na+K+/H+
Nha1p Canal antiporter de membrana citoplasmática Na+K+/H+
Nhx1p Canal antiporter de membrana pré-vacuolar Na+K+/H+
NSC1 Gene que codifica para o canal de influxo de Na+ e K+
OLE1 Gene que codifica para a desaturase de ácidos graxos Delta(9)
PB Pares de bases
Pdr12p Transportador ABC de membrana citoplasmática
PDR12 Gene que codifica para o transportador ABC de membrana
citoplasmática Pdr12p
PMA1 Gene que codifica para H+-ATPase de membrana citoplasmática
Pma1p H+-ATPase de membrana citoplasmática
PMA2 Gene que codifica uma isoforma da H+-ATPase de membrana
citoplasmática
Pma2p Isoforma da H+-ATPase de membrana citoplasmática
PMC1 Gene que codifica para a Ca+2- ATPase de membrana vacuolar em
leveduras
Pmc1p Ca+2- ATPase de membrana vacuolar de leveduras
PMR2 Gene que codifica para isoformas distintas da bomba de Na+ de
membrana citoplasmática, Ena.
PMSF Phenylmethylsulphonyl fluoride ou fluoreto de fenilmetilsulfonil
XII ROS Espécies reativas de oxigênio
RT-PCR Real time PCR ou PCR em tempo real
SSA1 Proteína chaperonina membro da subfamília SSA de proteínas
HSP70
SSA2 Proteína chaperonina membro da subfamília SSA de proteínas
HSP70
SSA3 Proteína chaperonina membro da subfamília SSA de proteínas
HSP70
SSA4 Proteína chaperonina membro da subfamília SSA de proteínas
HSP70
TCA Ácido tricloroacético
TM Temperatura ótima de anelamento
Trk1p/Trk2p Canal de influxo de K+ na membrana citoplasmática
UFC Unidade formadora de colônia
URL Unidade relativa de luminescência
Vmax Velocidade máxima
XIII
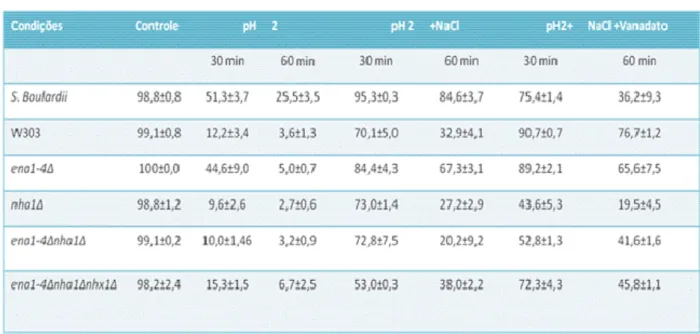
Tabela 1 Viabilidade das cepas Saccharomyces em pH 2na presença ou ausência
de íons Na+. 57
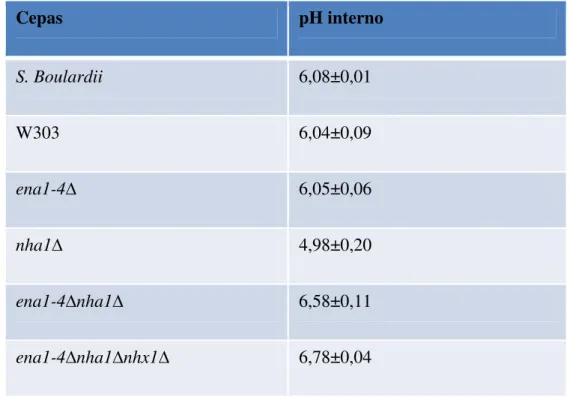
Tabela 2 pH intracelular de cepas Saccharomyces e mutantes envolvidos no
transporte de sódio, submetidos ao estresse ácido (pH 2 + 85 mM de
NaCl). 61
XIV
LISTA DE FIGURAS
Figura 1 Principais sistemas de transporte responsável pela homeostase de K+/Na+
em S. cerevisiae. 13
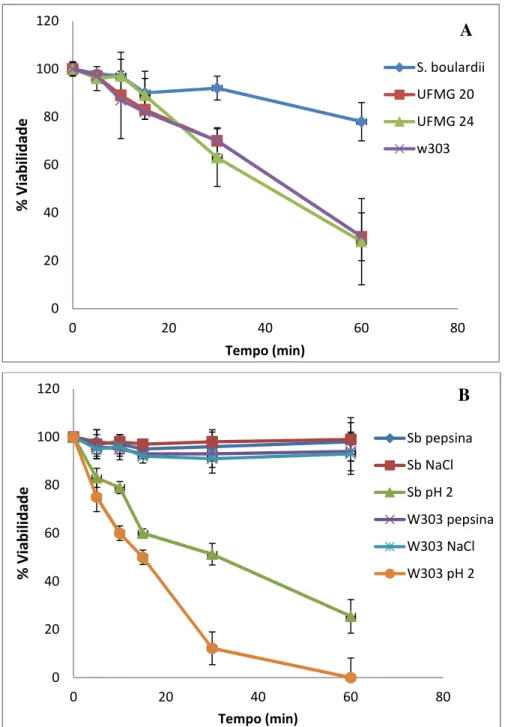
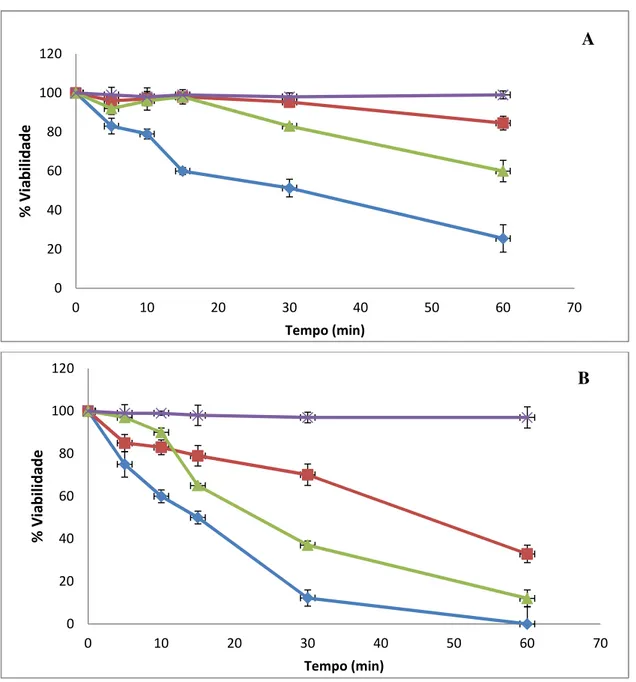
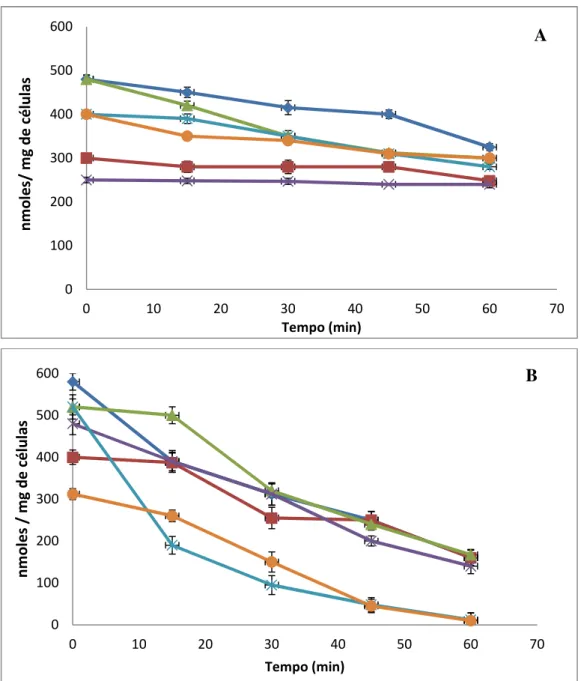
Figura 2 (A) Viabilidade das cepas de Saccharomyces em ambiente gástrico
simulado. (B) Efeito isolado dos componentes do ambiente gástrico
simulado na presença de NaCl. 51
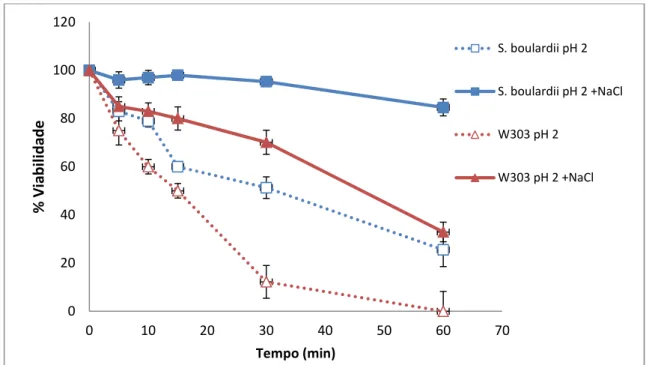
Figura 3 Viabilidade sob estresse ácido na presença de NaCl na viabilidade de S.
boulardii e S. cerevisiae W303 53
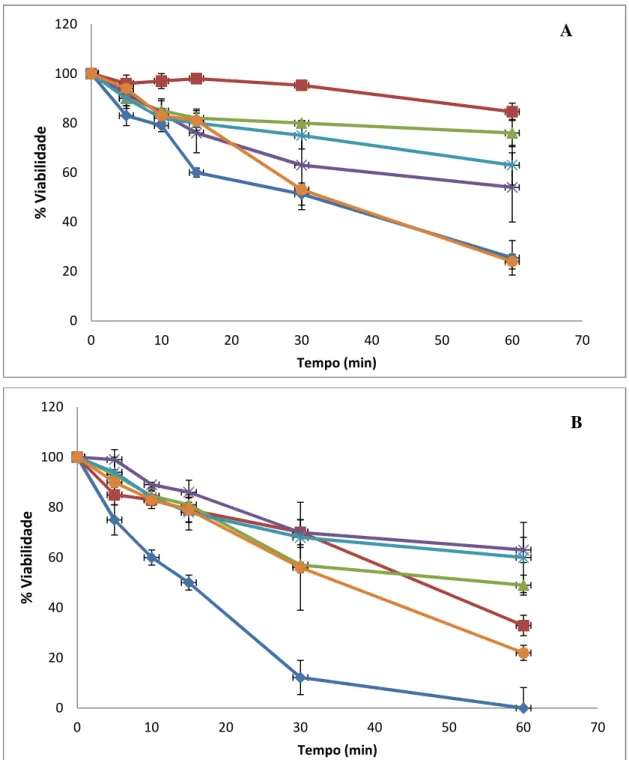
Figura 4 Efeito de sais na tolerância ao pH 2 em S. boulardii (A) e W303 (B). 54
Figura 5 Efeito de 5 µg/mL de cicloheximida na viabilidade de Saccharomyces
crescidas em meio YPD 4% e expostas em estresse ácido. S.boulardii
(Painel A) e S. cerevisiae W303 (Painel B). 58
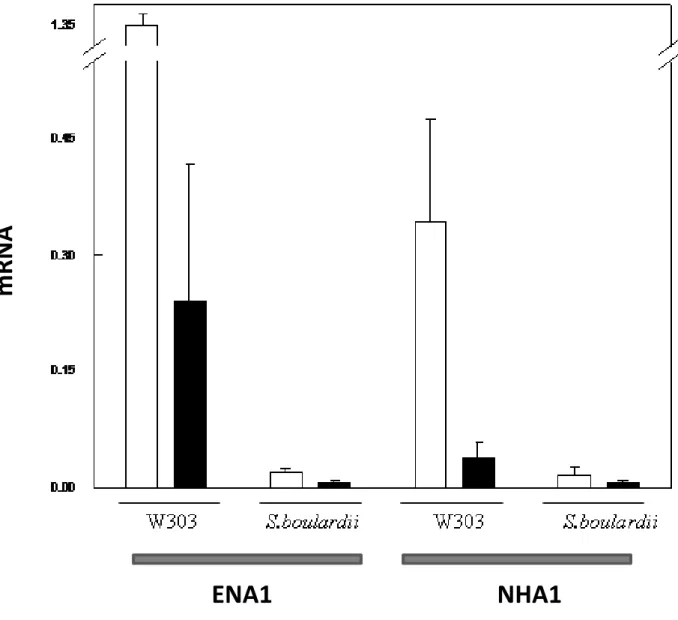
Figura 6 Nível de transcrição dos genes ENA1 e NHA1 em S. boulardii e S.
cerevisiae W303 por RT-PCR. 59
Figura 7 Alterações no conteúdo intracelular de Na+ (Painel A) e K+ (Painel B) em
XV
Figura 9 Medida de cálcio celular total, em cepas Saccharomyces. 69
Figura 10 Atividade Ca+-ATPásica das cepas Saccharomyces. 70
Figura 11 Sinal de cálcio das cepas Saccharomyces crescidas em YPD e submetidas
ao pH 2 71
Figura 12 Capacidade tamponante das cepas Saccharomyces crescidas em meio YPD
e submetidas ao estresse ácido. Painel A: Capacidade tamponante das células; Painel B: Capacidade tamponante dos extratos celulares. 73
Figura 13 Dendograma da análise lipídica da membrana plasmática de S. cerevisiae
W303 e S. boulardii. 75
Figura 14 Conteúdo de trealose, glicogênio e ATP em cepas Saccharomyces crescidas
2 1.- Introdução
A levedura Saccharomyces cerevisiae var. boulardii têm sido amplamente
estudada de acordo com suas propriedades probióticas e mecanismos de ação contra vários patógenos causadores de diarréias. Objetivando alcançar o intestino do hospedeiro em número satisfatório para exercer o papel probiótico, S. boulardii
necessita resistir as condições inóspitas enfrentadas no decorrer do trato digestivo, dentre elas, a resistência ao pH ácido, devido a presença do ácido clorídrico no estômago.
Os mecanismos pelos quais a cepa probiótica S. boulardii apresenta maior
resistência frente às diversas condições de estresse, ainda não estão bem esclarecidos, principalmente no que diz respeito ao estresse ácido.
A utilização de Saccharomyces cerevisiae como modelo de estudo para elucidar
os mecanismos de resposta ao estresse ácido tem sido quase que exclusivamente estudado em estresse com ácidos orgânicos, e há poucos estudos com respostas ao estresse com ácido inorgânico e mecanismos de proteção nestas condições.
As leveduras pertencentes ao gênero Saccharomyces são classificadas como
fungos do filo Ascomicota, ordem Saccharomycetalia, família Saccharomycetacea e
gênero Saccharomyces (Bisby, F. A. et.al., 2006).
1.1.1 - Saccharomyces cerevisiae
A levedura Saccharomyces cerevisiae é reconhecida como um organismo
modelo é um eucarioto simples cujo genoma pode ser facilmente manipulado. Estudos sobre a função de genes de mamíferos são frequentemente feitos em S. cerevisiae, que
ao contrário da maioria dos microrganismos, pode ser tanto haplóide, diplóide ou mesmo poliplóide.
3
mutantes, além de possuir um sistema de transformação de DNA bastante versátil (Goffeau, A. et. al, 1996; Hughes, T. et.al., 2004; Stambuk, B. U. A., 1999). S.
cerevisiae foi o primeiro organismo eucarioto em que o genoma foi completamente
sequenciado e depositado em bancos de dados públicos como http//www.yeastgenoma.org, o que permitiu a clonagem de diferentes genes, sua rápida identificação e caracterização, bem como a melhor compreensão da sua função celular.
S. cerevisiae tornou-se modelo eucarioto ideal, com ferramentas moleculares
apropriadas para manipulação gênica e estudos de expressão, como o transcriptoma, proteoma, metaboloma, que propiciam esclarecimento de diversas rotas metabólicas (Da Silva, M. E. e cols., 1990; Murphy, A. e cols.,1999; Zigova, J., 2000; Pena-Castillo e
Hughes, T., 2007; Jones, G.M. e cols., 2008; Maya, D. e cols.,2008; Schuster, S. e cols.,
2008).
A célula de levedura possui mecanismos de adaptação eficientes para manutenção da alta taxa de crescimento, mesmo após as mudanças promovidas pelo próprio metabolismo, o que está diretamente relacionado à capacidade de regular a expressão de genes apropriados ao regime nutricional em que a célula se encontra. A expressão de genes e a consequente síntese de proteínas são diretamente influenciadas pelas condições nutricionais (KruckeBerg, R., R. e cols., 1998).
A levedura é uma fonte biológica para obtenção de produtos de interesse na indústria farmacêutica e de alimentos, como por exemplo, invertase, hexoquinase, glicose-6-fosfato desidrogenase, glicose oxidase, proteases e outras, além da possível utilização de sua própria biomassa celular como fonte de proteínas na alimentação animal e humana (Pessoa Jr e cols., 1996; Silva, M. E., 1998; Benítez e cols., 1996).
Diferentes modificações genéticas podem gerar efeitos no metabolismo de
S. cerevisiae, obtendo-se assim cepas específicas para a produção de substâncias de
4 1.1.2 - Saccharomyces boulardii
A levedura S. boulardii não patogênica foi isolada da fruta lichia (Litchi
chinensis) na Indochina. Desde o início dos anos oitenta esta levedura tem sido
amplamente estudada quanto a sua ação no tratamento e prevenção a DAA (Diarréia Associada a Antibióticos), para avaliação dos benefícios de S. boulardii no organismo
hospedeiro e descrição do mecanismo de ação (Mccullough, M. J. e cols., 1998).
Um aspecto controverso, relativo à levedura S. boulardii, diz respeito à sua
taxonomia (Mccullough, M. J. e cols., 1998; Van Der AA Kuhle, A., A. e Jespersen, L.,
2003). Inicialmente foi classificada por Cardinali, G. e Martini, A. em 1994, por meio da cariotipagem eletroforética comparativa e análise multivariada do polimorfismo, a cepa S. boulardii não pertencente à espécie S. cerevisiae e também das outras espécies.
O progresso no desenvolvimento da filogenia molecular permitiu a classificação de muitas espécies de leveduras por técnicas como, polimorfismo de microssatélite, amplificação aleatória de polimorfismo e análise polimórfica de fragmento de restrição. Dessa forma, Edwards Ingram e cols. (2007) analisando o genoma pela hibridização
genômica comparativa concluiu que S. cerevisiae e S. boulardii são membros da mesma
espécie, entretanto geneticamente foram detectadas características como trissomia do cromossomo IX e números de cópias dos genes alteradas em S. boulardii que a diferem
de S. cerevisiae.
Estudos de caracterização molecular, desenvolvidos em nosso laboratório, sugerem que S. boulardii é uma cepa da espécie S. cerevisiae. No entanto, S. boulardii e
cepas de S. cerevisiae apresentam diferenças fisiológicas importantes (Fietto, J. L. R. e
cols., 2004). Estas diferenças incluem respostas a situações de estresses, como
resistência a altas temperaturas e ao estresse ácido.
S. boulardii é utilizada como probiótico devido a sua capacidade de atingir
rapidamente altas concentrações no cólon, manter níveis constantes, não colonizar permanentemente o intestino e não translocar facilmente para fora do trato intestinal (Berg, R. e cols., 1993; Boddy, A. V. e cols., 1991). S. boulardii quando introduzida na
5
vários agentes etiológicos da diarréia e bem como das toxinas associadas a estes agentes (Elmer, G.W. e cols., 1987).
A partir de 1960 iniciou-se a comercialização da levedura liofilizada pelo “Laboratoires Biocodex” (Paris, França). Desde então, o uso comercial de S. boulardii
como medicamento no combate às diarréias foi difundido em toda Europa e está disponível no mercado sob diversos nomes comerciais, tais como: Ultra-Levure® (França), Florastor® (Estados Unidos), Precosa® (Dinamarca), Levucell® (Lallemand, Canadá), Perenterol® (Alemanha), Perenteryl® (Chile), Codex® (Itália), Floratil® (Brasil). Atualmente, a levedura é comercializada na Europa, América do Sul e do Norte, Ásia e África (McFarland e Bernasconi, 1993), tendo sido a sua utilização liberada pela FDA (Food and Drug Administration) nos Estados Unidos (Florastor,
2007). Os direitos de comercialização para a América do Sul foram adquiridos pelas
indústrias químicas da MERCK S.A., e no Brasil outras preparações contendo
S. boulardii encontram-se disponíveis (Martins, F. S. e cols., 2005).
A palavra probiótico que significa “para vida” é um termo derivado da língua grega (Neves, M. J., 2005). A terminologia conceitual foi inicialmente proposta por Fuller (1991), após os primeiros ensaios clínicos. Probióticos podem ser definidos como suplementos alimentares que contêm microrganismos vivos, ou componentes microbianos que, quando ingerido em determinado número, apresentam efeito benéfico sobre a saúde e bem-estar do hospedeiro (Salminem e cols., 1999; Dupont, 2001;
FAO/WHO, 2001; Isoulari, E., 2001; Reid, G. et al, 2001; Sanders, D., 2003).
As células probióticas depois de ingeridas devem ser capazes de sobreviver às condições de estresse presentes no trato gastrointestinal, como suco gástrico, presença de sais biliares, enzimas digestivas e manter sua viabilidade e atividade metabólica no intestino para exercerem os efeitos benéficos aos hospedeiros. Quanto aos desafios tecnológicos para a produção industrial de células, estas devem manter-se estáveis e viáveis em níveis satisfatórios durante todo o prazo de validade do produto (Saad, 2006; Araújo, 2007).
6
que S. boulardii inibe a ligação e a enterotoxicidade da toxina A de Clostridium difficili
em íleo de ratos. Dias, R. S. e cols. (1995) comprovaram a capacidade desta levedura de
inibir e neutralizar a enterotoxicidade das toxinas de Escherichia coli e Vibrio cholerae.
Czerucka, D. e cols. (1994) demonstraram que S. boulardii produz uma proteína
de 120 kDa capaz de neutralizar o efeito da toxina do cólera, propondo que este efeito esteja relacionado à habilidade desta proteína de se ligar a sítios preferenciais nas células intestinais do paciente e desta forma regular os níveis intracelulares de AMPc, visto que o aumento de AMPc promovido pela ação da toxina gera, de forma indireta, a perda de água pelas células intestinais.
Outro mecanismo protetor importante é o aumento da resposta imunológica do hospedeiro contra as toxinas diarréicas mediado pelo uso de S. boulardii (Qamar, A. e
cols., 2001). Em trabalho realizado por Brandão, R. L. et al (1998) no nosso laboratório
propõe-se um mecanismo protetor no qual a levedura atuaria competindo pela ligação com a toxina. Os resultados sugerem a adesão da toxina do cólera à levedura pela subunidade B, seguida da internalização da subunidade A, similarmente ao que acontece nas células intestinais. Esta internalização eleva os níveis de AMPc na célula e desencadeia cascatas de fosforilações dependentes de AMPc. Resultados posteriores de Neves, M. J. e cols. (2002), também obtidos pelo grupo de pesquisa no nosso
laboratório, mostraram que a toxina do cólera se liga, de forma específica, a células de
S. boulardii e sugerem que a atividade patogênica desta toxina (e provavelmente de
outras toxinas) pode ser neutralizada por esta ligação. Os estudos de ligação com 25I – toxina do cólera, assim como o sinal de AMPc, dependente da integridade da molécula, sugerem fortemente a presença de receptores mais ou menos específicos na superfície da levedura (similar a situação observada em células de mamíferos).
7
A grande maioria dos probióticos que são comercializados é composta por
bactérias. As leveduras que são utilizadas como probiótico ficam limitadas por
S. boulardii na medicina humana e S. cerevisiae na medicina veterinária. A utilização
de leveduras como probiótico é um investimento rentável e seguro, pois as mesmas podem ser comercializadas liofilizadas, são rapidamente eliminadas pelo organismo após interrupção da terapia e não são afetadas pelo uso de antimicrobianos (Blehaut e
cols., 1989; Boddy, A. V. e cols., 1991).
Resposta a estresses constitui um requerimento funcional para a escolha de um probiótico (Matilla-Sandholm e cols., 2002). Para atingir o intestino, o probiótico
precisa transpor diversas situações de estresses. Após a ingestão por via oral, o microrganismo tem que se manter viável numa temperatura interna do hospedeiro de 37°C. Adicionalmente, o microrganismo entra em contato com o estômago, um ambiente inóspito devido ao pH ácido, e em seguida alcança o intestino encontrando um ambiente totalmente adverso em situação de pH básico.
Dentre os fatores que possam levar a diminuição do número de células probióticas ingeridas pelo hospedeiro tem-se ainda, a ação dos sucos intestinais (sais biliares e pancreáticos) e menor tensão de oxigênio no trato gastrointestinal. Além destas variações, existe a competição entre a microbiota nativa e o probiótico. Neste caso, microrganismos com maior flexibilidade nutricional levam vantagem. Sendo assim, é essencial que microrganismos potencialmente selecionados para serem utilizados como probióticos sejam tolerantes aos produtos secundários (água oxigenada, gazes e ácidos) do metabolismo do hospedeiro e substâncias antagônicas gerados por outros microrganismos (Holzapfel, W. H., 1998).
1.2 - Mecanismos envolvidos em resposta a estresse em leveduras
8
podem reduzir a atividade enzimática, interromper o fluxo metabólico, danificar as estruturas celulares e alterar os gradientes químicos levando a célula a uma condição de instabilidade (Ferreira e Mager, 1993). Porém, as células são capazes de manter a homeostase interna mesmo diante de um ambiente externo variado (Hohmann, S. e Mager, W. H., 1997; Gasch, 2001).
A resposta aos diferentes tipos de estresse é uma característica fundamental na adaptação dos organismos vivos às condições adversas do ambiente. As células de S.
cerevisiae, quando submetidas à condição de estresse, desenvolvem uma rápida resposta
molecular para reparar danos e proteger as estruturas celulares dos efeitos causados pelo estresse (Ruis, H. e Schuller, C., 1995; Swan, T.M. e Watson, K., 1998; Estruch, F., 2000). Essa resposta é caracterizada pela síntese de proteínas específicas, aumento do nível celular de trealose e glicerol, alteração da composição lipídica da membrana plasmática e da atividade da H+ - ATPase, modulação do processo de troca iônica e no caso de estresse oxidativo produção de glutationa e de enzimas superóxido dismutase (Birch, R. M. e Walker, G. M., G. M., 2000).
Os mecanismos de resposta ao estresse são evolutivamente conservados em todos os organismos (eucariotos e procariotos), e as repostas protetoras de organismos vivos foram inicialmente identificados em estudos de reposta a choque térmico. A resposta ao choque térmico é caracterizada pela síntese de proteínas específicas denominadas de proteínas de choque térmico ou Hsp (heat shock proteins), que
9
B, C e D (Estruch, F., 2000; Baumann, F. e cols., 2000). O subgrupo A é o mais
complexo, contendo quatro genes, SSA1, SSA2, SSA3 e SSA4. Embora os genes da
família HSP70 codifiquem proteínas que desempenham funções semelhantes, eles diferem no grau de expressão em condições de estresse (Estruch, F., 2000).
Em leveduras, dois aspectos da resposta ao estresse por choque térmico têm sido bem analisados: a síntese de proteínas de resposta ao estresse ou relacionada ao acúmulo de trealose (Swan, T.M. e Watson, K., 1998). Em culturas de S. cerevisiae
ocorre o acúmulo de trealose em resposta a uma mudança na temperatura de 30°C para 44°C, é quando a concentração de trealose atinge valores intracelulares máximos (Cavalheiro e cols., 1999).
As leveduras quando expostas a um estresse osmótico perdem água dando início a uma série de mecanismos, principalmente relacionados com a síntese de trealose e de glicerol, isto ocorre para resguardar as células da desidratação e proteger as estruturas celulares dos possíveis efeitos da condição de estresse (Estruch, F., 2000; Mager, W. H. e Siderius, M., 2002). Durante muito tempo sustentou-se a hipótese de que a trealose era somente um carboidrato de reserva (Singer, M. A. e Lindquist, S., 1998), entretanto o papel mais importante que ela desempenha nas células de levedura é de estabilizar a membrana celular em condições de estresse (Singer, M. A. e Lindquist, S., 1998; Fujii, S. e cols., 1996). Demonstrada a sua relação com a termotolerância de fungos e
leveduras, o papel do açúcar ganhou importância. Em leveduras a síntese e o acúmulo de trealose estão diretamente relacionados com o aumento de sua sobrevivência quando submetidas a diferentes situações de estresse, como desidratação, congelamento, alta concentração de etanol, oxigenação e CO2 (Shima, J. e cols., 1999; Majara, M. e cols., 1996; Paredes, V. e cols., 2003).
Muitos estudos têm documentado a alteração da composição lipídica celular em resposta a outro tipo de estresse, a exposição ao etanol, por exemplo. Desde que membranas celulares têm sido consideradas como alvos primários do estresse com etanol, muitos pesquisadores têm sugerido uma relação entre a composição de ácidos graxos de membranas lipídicas e a tolerância ao estresse com etanol (You, K. M. e cols.,
10
Chi e Arneborg (1999), que demonstraram que uma linhagem de S. cerevisiae tolerante
ao etanol possuía elevados níveis de ácido oléico e uma significativa redução de deficiência respiratória induzida pelo etanol, em comparação com uma linhagem sensível ao álcool.
Em S. cerevisiae, ácidos graxos insaturados são formados na superfície do
retículo endoplasmático pela enzima desaturase, que é codificada pelo gene OLE1. Essa
enzima converte ácidos graxos saturados a espécies monoinsaturadas, pela introdução de uma dupla ligação entre os carbonos 9 e 10 dos ácidos palmítico (C16:1) e esteárico (C18:0) para formar os ácidos palmitoléico (C16:1) e oléico (C18:1), que são os ácidos graxos insaturados predominantes em S. cerevisiae (Stukey e cols., 1989). As
características dinâmicas e estruturais da membrana podem mudar pelas alterações das condições ambientais, ou seja, mudança na composição molecular da membrana ou adição de moléculas estranhas que interagem com os constituintes da mesma. Isso explica porque mudanças na fluidez da membrana são observadas em resposta a muitos estresses ambientais, e porque as células mantêm um ótimo nível de fluidez dentro da matriz lipídica (Beney, L. e Gervais, P. , 2001).
Células de leveduras crescendo em condições de aerobiose produzem espécies reativas de oxigênio (ROS) como peróxido de hidrogênio, radicais hidroxilas e ânions superperóxidos, provenientes de processos metabólicos normais como respiração e β -oxidação de ácidos graxos (Gordon, C. e cols., 1999; Estruch, F., 2000; Costa, V. e
Moradas-Ferreira, P., 2001). Esses compostos constituem fatores de estresse oxidativo, induzindo um grupo de genes que codificam proteínas antioxidantes (Estruch, F., 2000). Embora as linhagens industriais sejam normalmente resistentes ao estresse oxidativo, esse tipo de estresse pode afetar a produção de biomassa de leveduras de vinificação, processo este realizado em aerobiose (Carrasco, P. e cols., 2001). Linhagens de S.
cerevisiae tolerantes ao estresse oxidativo também apresentam tolerância a outras
formas de estresse, porém essa co-tolerância não é comum a todas as linhagens e depende do tipo e da intensidade do estresse (Lewis, J. G. e cols., 1997).
11
extrusão de sódio do citoplasma e o acúmulo simultâneo de concentrações elevadas de solutos compatíveis para evitar a perda de água (Organismos excluder), em
contrapartida existem os organismos que acumulam altas concentrações de sódio,
conseguindo evitar a intoxicação (Organismos includer) (Ramos e cols., 1999).
S. cerevisiae é uma levedura excluder por manter níveis citoplasmáticos baixos de sódio
através do processo de extrusão ou dirigindo-o para o vacúolo. Em ensaios de resistência ao estresse causado por elevada concentração de sal no meio, linhagens de panificação de S. cerevisiae apresentam resistência a até 1,5 M de NaCl (Lewis, J. G. e
cols., 1997) enquanto que em linhagens de vinificação foram encontradas leveduras
resistentes a 3 M de KCl (Carrasco, P. e cols., 2001).
As diferentes vias metabólicas são coordenadas por uma série de genes que codificam proteínas reguladoras da expressão gênica. A integração entre as várias vias metabólicas mostra que muitos genes estão envolvidos em diferentes respostas celulares e são regulados por um conjunto de proteínas regulatórias que atuam em mais de uma via.
Em S. cerevisiae, mais de 70% dos genes envolvidos na biogênese de
ribossomos foram reprimidos após a adição de 0,5 M de NaCl ao meio de crescimento, porém vários genes envolvidos no metabolismo de glicose, no transporte de elétrons e na resposta ao estresse foram induzidos. A redução da expressão gênica dos genes envolvidos na biogênese dos ribossomos resulta em uma redução na síntese protéica. Essa redução é importante para que a célula redirecione seus fluxos metabólicos em resposta à condição de estresse (Hirasawa, T. e cols., 2005).
Muitos mecanismos de resposta a estresses envolvem a participação de íons, portanto, alterações iônicas são fundamentais para que haja um desencadeamento na ativação das vias de sinalização. O modo de ação para que se estabeleça o efeito positivo dos íons, deve envolver a homeostase iônica. Em leveduras, a manutenção da homeostase de Na+ e K+ é dependente de sistemas transportadores responsáveis pelo influxo e efluxo de sais, tais como a bomba exportadora de sódio Ena1p (Benito B. e
cols., 1997), trocadores de Na+/H+ Nha1p localizado na membrana plasmática, Nhx1p
12
presente na membrana do vacúolo responsável pela homeostase de íons Na+ e K+ e regulação do pH (Cagnac, O. e cols., 2007). Além da captação de Na+ e K+ pelos
transportadores Trk1p e Trk2p (Ko, C. H. e Gaber, R.F., 1991) existem outros canais menos específicos como o NSC1, ainda não identificado geneticamente (Bihler, H. e
cols., 2002). Um modelo dos principais sistemas envolvidos na homeostase de K+/Na+ e
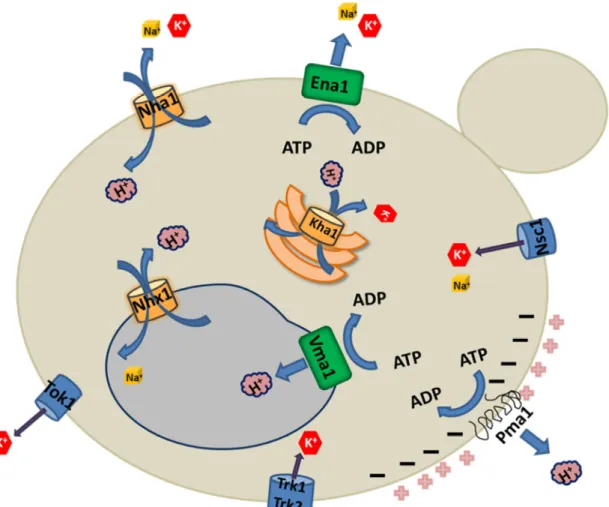
manutenção do potencial de membrana pode ser visualizado na figura 1.
13
Figura 1 – Principais sistemas de transporte para manutenção da homeostase de K+/Na+ em S. cerevisiae. Os transportadores Trk1/2p tem alta afinidade para K+. A Kha1p é um
provável trocador de K+/H+. A extrusão de Na+ é primariamente mediada pela ATPase Ena1p responsável pelo bombeamento de sódio, mas também pelo Nha1p Na+/H+ antiporter. NSC1 é um canal para cátions, não seletivo. A compartimentalização de Na+
é mediada pelo antiporter pré-vacuolar Nhx1p Na+/K+. A membrana pré-vacuolar/vacuolar é energizada pela bomba de próton ATPase tipo –V (Vmap) e a membrana plasmática pela ATPase tipo –P (Pma1p).
14 1.3 – Mecanismos de resposta ao estresse ácido
A maioria das células de eucariotos superiores possui altas concentrações de proteínas em seu citoplasma que funcionam como ácidos ou bases orgânicas. Proteínas contendo maior número de histidina, por exemplo, tamponam o meio intracelular em pH próximo do neutro. Nucleotídeos semelhantes a ATP, além de outros metabólitos de baixo peso molecular contêm grupos ionizáveis que também tamponam o citoplasma (Nelson e Lox, 2004).
Algumas leveduras são capazes de crescer em meio com pH entre 4,5 e 6,5 (Walker, G. M., 1998), porém ácidos orgânicos como ácido sórbico, o ácido benzóico e o ácido acético, os quais têm sido utilizados na preservação de alimentos, inibem o crescimento de leveduras em baixo pH (Piper, P. e cols., 1998; Piper, P. e cols., 2001).
Ácidos orgânicos, quando não dissociados, são capazes de atravessar a membrana plasmática por difusão (Cássio, F. e cols., 1987). Uma vez no citoplasma esses ácidos se
dissociam gerando o próton H+ e o ânion correspondente. Quanto maior o pH do meio, maior o grau de dissociação do ácido. Um ácido orgânico quando se dissocia no meio intracelular diminui o pH citosólico. O acúmulo do ânion no citoplasma pode aumentar a produção de radicais livres, em aerobiose, induzindo o estresse oxidativo (Piper, P. e
cols., 2001).
Em S. cerevisiae a presença de ácido sórbico no meio de crescimento aumenta a
atividade da enzima H+ - ATPase, a qual catalisa a extrusão de prótons H+, e induz a síntese da proteína Pdr12p, que age na membrana plasmática catalisando a extrusão do íon sorbato as custas de ATP ( Piper, P. e cols., 2001). Células de S. cerevisiae
crescendo em pH 7,0 na ausência de sorbato, apresentam uma baixa concentração de Pdr12p na membrana plasmática, porém a adição de sorbato ao meio de cultura aumenta o grau de expressão do gene PDR12.
Mutantes pdr12 são muito sensíveis a sorbato em baixo pH, indicando que a
15
em YEPD (Extrato de Levedura e Glicose) ajustado a pH 4,5 com HCl, porém essa resistência não depende exclusivamente da atividade de Pdr12p.
Outras proteínas envolvidas em diversas funções na célula são importantes na resposta ao estresse ácido. Células mutantes pkf1∆ e pkf2∆ (subunidades α e β da fosfofrutoquinase, respectivamente) rpe1∆ (D-ribulose-5-fosfato 3 epimerase) gdh3∆ (uma isoenzima da gliceraldeído-3-fosfato desidrogenase) e tps2∆ (trealose-6-fosfato fosfatase) são sensíveis a 2 mM de sorbato no meio de crescimento (Cheng, L. e cols.,
1999).
Os ácidos orgânicos inibem a resposta ao estresse de choque térmico e aumentam a frequência de mutações que causam deficiência no metabolismo respiratório de S. cerevisiae, quando cultivada a elevadas temperaturas (Cheng, L. e
cols., 1999). O tratamento de células de S. cerevisiae com ácidos orgânicos inibe a
síntese das Hsp e, consequentemente, a termotolerância (Piper, P. e cols., 1998).
A presença de ácido acético na concentração de 50 mM no meio de crescimento (pH 3,5) semelhantemente ao ácido sórbico, diminui o pH citosólico e aumenta a atividade da H+-ATPase na membrana plasmática de células de S. cerevisiae (Carmelo,
V. e cols., 2005). Linhagens de panificação dessa levedura foram sensíveis a 0,4% de
ácido acético (v/v) em meio YEP ágar (meio sem glicose). Nessas condições, a sobrevivência da maioria das linhagens testadas foi inferior a 10% do total de células inoculadas no meio (Lewis, J. G. e cols., 1997). Células de S. cerevisiae pré-adaptadas
ao estresse ácido, quando presentes em meio mínimo, sem glicose, contendo 1mM de ácido benzóico (pH 4,5), acumulam uma grande quantidade de benzoato no meio intracelular, porém quando a glicose é adicionada ao meio de cultura as células eliminam cerca de 90% do ácido benzóico absorvido. Isso mostra que a célula depende de glicose ou de outra fonte de energia, para gerar ATP e fazer funcionar a H+-ATPase e a proteína Pdr12p, restaurando a homeostase do pH intracelular ( Henriques, M. e cols.,
1997).
Contribuindo para este conjunto de evidências, Ludovico, P. e cols. (2001)
verificou que o tratamento de S. cerevisiae com 20-200 mM de ácido acético, durante
16
concentração de ácido necessária para a ocorrência de um processo ativo de morte celular revelou-se crítica. Com efeito, enquanto que a morte celular observada para concentrações acima de 80 mM não era inibida pela cicloheximida (inibidor da síntese protéica) e estava associada a alterações ultra-estruturais típicas de necrose, a morte induzida por concentrações mais baixas de ácido acético (20-80 mM) foi parcialmente inibida por cicloheximida. Nestas condições foi possível detectar diferentes alterações estruturais típicas de apoptose nomeadamente: i) condensação extensiva de cromatina na periferia do invólucro nuclear observada por microscopia eletrônica de transmissão; ii) exposição de fosfatidilserina na superfície da membrana citoplasmática avaliada por reação com anexina V conjugada com FITC e iii) ocorrência de quebra de DNA avaliada por TUNEL. O fato de o ácido acético ser um subproduto da fermentação alcoólica produzida por S. cerevisiae e, portanto, bastante corriqueiro no seu ambiente
natural, parece indicar que este processo, à semelhança do que acontece nos eucariotos superiores, pode ter um papel fisiológico relevante no ciclo de vida da levedura.
No trabalho de Claret, S. e cols. (2005) foi avaliado o efeito da exposição de
S. cerevisiae em estresse em baixo pH (HCl) e a ativação da via PKC como resposta.
Durante condições de crescimento em baixo pH a via PKC foi ativada, e o sensor Mid2p foi identificado como decisivo na ativação desta via, demonstrando que os componentes Bck1p e Slt2p da cascata MAPK são essenciais em ambiente ácido. Além desses componentes, também foi constatado o envolvimento de RGD1 na via PKC em
pH ácido, comprovando a interação de duas vias, uma envolvendo o sensor Mid2p e o outro envolvendo Rgd1p, ambos convergem na via de integridade celular, traduzindo a sinalização em pH baixo. Demonstraram também, que a interrupção das duas vias envolvidas gera mutantes incapazes de sobreviver sob condições de estresse ácido. Entretanto, em contraste do que foi observado com os mutantes bck1∆ ou slt2∆, o suporte osmótico não eliminou a letalidade dos mutantes rgd1∆ e rgd1∆mid2∆ provocada pelo choque ácido, sugerindo que a ação mediada por Rgd1p na tolerância ao estresse ácido não está limitada a via PKC.
Malakar, D. e cols. (2006) investigaram a função protetora da adição de
17
(10 mM HCl, pH∼2), demonstrando que este estresse provoca redução do espaço periplasmático da membrana plasmática, e substancial aumento no número de vacúolos no citoplasma, entretanto a adição de 1mM de Adomet, nesta condição, além de reduzir o número de vacúolos formados, amenizou as variações de pH interno observadas em estresse ácido.
Resultados preliminares no nosso laboratório demonstraram respostas a diferentes estresses pelo probiótico S. boulardii e que esta cepa responde melhor aos
estresses de temperatura e ao ambiente gástrico simulado, quando comparado a cepas
S. cerevisiae de laboratório (Fietto, J. L. R. e cols., 2004). Neste trabalho, evidenciamos
a resistência de S. boulardii ao estresse ácido, além de observar que a adição de sódio,
um componente do suco gástrico, em pH 2, confere significativa proteção a esta cepa probiótica. Os mecanismos pelos quais o estresse ácido afeta o metabolismo das cepas
S. cerevisiae avaliadas, e as diferenças na resposta de S. boulardii frente à proteção
18
19 2 - Objetivos
2.1 - Objetivo Geral
Caracterizar a resposta da levedura S. boulardii ao pH ácido e o efeito de NaCl,
em baixas concentrações, nesta resposta.
2.2 - Objetivos Específicos
• Comparar a viabilidade de diferentes cepas Saccharomyces cerevisiae frente a
uma simulação do ambiente gástrico;
• Identificar os componentes do ambiente gástrico responsáveis pela perda de viabilidade celular;
• Estudar o efeito do pH ácido (HCl / pH 2) sobre a viabilidade de diferentes cepas S. cerevisiae;
• Avaliar o efeito da adição de diferentes íons sobre a viabilidade celular em pH ácido (HCl / pH 2);
• Investigar possíveis mecanismos envolvidos no efeito protetor de sódio, por meio de medidas da expressão dos genes envolvidos na homeostase deste íon; medidas de acúmulo e extrusão de sódio; medidas de potencial de membrana, em cepas com deleções em genes que codificam proteínas envolvidas na homeostase de sódio;
20
• Verificar a homeostase do cálcio em células submetidas ao estresse ácido (pH 2 / HCl);
• Determinar a capacidade tamponante das células e dos extratos celulares obtidos de células submetidas ao estresse ácido (pH 2 / HCl);
• Analisar a composição lipídica de membrana das cepas Saccharomyces, em
reposta ao estresse ácido;
21
22 3 - Material e Métodos
3.1 - Cepas Saccharomyces utilizadas nos experimentos
Nos experimentos foram utilizadas cepas de Saccharomyces cerevisiae e
mutantes com deleções nos genes envolvidos no transporte de sódio. As cepas utilizadas estão demonstradas na tabela 1:
Cepas Genótipo Procedência
Saccharomyces cerevisiae
var. boulardii
Selvagem Floratil®,Merck S.A.
UFMG 20 Selvagem Martins, F. S. e cols.,
2008
UFMG 24 Selvagem Martins, F. S. e cols.,
2008 W303 MATa leu2-3,112 ura3-1 trp1-1
his3-11,15 ade2-1 can1-100 GAL mal SUC2
Johan M. Thevelein, J.M.
Laboratorium voor Moleculaire
Celbiologie, Katholieke
Universiteit Leuven, Belgium.
LBCM 479 W303 Matα ena1 :: HIS3 :: ena4 José Ramos
Department of Microbiology, ETSIAM, Edificio Severo Ochoa, Campus de Rabanales, University of Cordoba, Spain
LBCM 511 W303 Matα nha :: LEU2 Hana Sychrová, H.
Departments of Membrane
23
Academy of Sciences CR, Prague, Czech Republic
LBCM 500 W303 Matα ena1 :: HIS3 :: ena4 nha1 :: LEU2
Hana Sychrová, H. Departments of Membrane
Transport, Institute of Physiology, Academy of Sciences CR, Prague, Czech Republic
LBCM 480 W303 Matα ena1::HIS3 ::ena4 nha1 :: LEU2nhx1:: TRP1
Hana Sychrová, H. Departments of Membrane
Transport, Institute of Physiology, Academy of Sciences CR, Prague, Czech Republic
3.2 - Meios de cultura e condições de cultivo
O meio de cultura utilizado nos experimentos foi YPD composto por extrato de levedura 1% (p/v), bacto-peptona 2% (p/v) e glicose 2% (YPD). O meio sólido foi acrescido de ágar 1,5% (p/v).
As células da levedura Saccharomyces foram pré-crescidas em tubos de ensaio
24 3.3 - Condições de estresse
As células crescidas em meio YPD 4% até D.O.600nm = 1,0, foram coletadas por centrifugação 3750 rpm, a 4°C, lavadas duas vezes com YP. Após a lavagem, as células foram ressuspensas no mesmo volume de YP para coleta da condição controle. As amostras foram centrifugadas, lavadas e ressuspensas de forma a conversar a D.O. 600nm 1,0 em solução aquosa pH 2 ajustado com HCl 1M, e em solução aquosa contendo 85mM de NaCl, pH 2 ajustado com HCl 1M, a 37°C, sob agitação lenta.
3.3.1 - Teste de Viabilidade
As células submetidas ao estresse ácido e estresse ácido acrescido de 85mM de NaCl foram coletadas em alíquotas de 100 µL nos tempos 0, 30 e 60 minutos. Essas alíquotas foram lavadas 2 vezes com YP, e ao sedimento celular foram acrescentados 100µL de solução corante de azul de metileno (azul de metileno 0,01% em tampão acetato de sódio 0,1M pH 5,0) (Mills, D.R., 1941). Após incubação por 10 minutos à temperatura ambiente, realizou-se a contagem das células em microscópio óptico. No campo de visualização foram contadas 300 células, em triplicata, em cada tempo analisado. A determinação da % de células viáveis foi feita através da seguinte fórmula:
% Células Viáveis = Total de Células – Número de Células Azuis X 100 Total de Células
25
3.4 - Reação em cadeia da polimerase em tempo real
3.4.1 - Extração e preparação de RNA total de levedura
As células de leveduras foram crescidas em 20 mL de meio YPD 4% até atingirem D.O.600nm = 1,0. Após o crescimento, alíquotas da cultura foram coletadas por centrifugação a 3750 rpm por 5 minutos a 4°C para o tempo controle, parte da cultura foi centrifugada e submetida ao estresse ácido em igual volume, a 37°C, sob leve agitação. As amostras foram coletadas em intervalos de 30 minutos, congeladas em nitrogênio líquido e estocadas a -80°C para posterior extração de RNA total.
O sedimento celular congelado em -80°C foi ressuspenso em 2 mL de fenol / água (3,75:1) e 2 mL de TES (10mM Tris:Cl; 10mM EDTA; 0,5% SDS; pH 7,5) e agitado vigorosamente em vórtex. Os tubos foram incubados em banho-maria a 70º C por 1 hora, e agitados vigorosamente em vórtex em intervalos de 5 minutos. Os tubos
foram centrifugados a 3000g, por 10 minutos, a 4º C e a fase superior coletada em tubos de microcentrífuga. Foram adicionados 800 µL de solução fenol e água na proporção de 3,75:1, e após nova agitação os tubos foram centrifugados a 3000g por 10 minutos a 4º
C. O sobrenadante foi coletado e foram adicionados 600 µL de clorofórmio. Procedeu-se à nova agitação em vórtex e em Procedeu-seguida, os tubos foram centrifugados a 3000g por 10 minutos a 4º C. O sobrenadante foi coletado e foram adicionados 50µL de acetato de sódio 3M pH 5,3 e 1 mL de etanol gelado 100%, sendo incubados a –80º C por 15 minutos. Após esta incubação as amostras foram centrifugadas a 15000 rpm por 10
minutos a 4º C, o sobrenadante foi descartado e o sedimento lavado com 500 µL de etanol 70%. As amostras foram centrifugadas e o sobrenadante novamente descartado. O RNA foi ressuspenso em 50 µL de H2O DEPC (200mL água Mili-Q estéril, 200µL DEPC) e armazenado a -80ºC até a utilização.
26
pH 7.0; EDTA 0,1 M) e brometo de etídio. Após 3 horas de corrida a 70 V, o gel foi fotografado sob luz ultravioleta.
3.4.2 - Obtenção do cDNA
Para síntese de cDNA foi utilizado o RNA total extraído anteriormente. 3µg de RNA foi tratado com DNAse para remoção total de DNA genômico das amostras, na presença da enzima RNAse OUT para proteção do RNA.
Eliminação de DNA (Promega)
RNA total (3µg) X µL
Enzima DNAseI 1 µL
Tampão da Enzima 1 µL
RNAse OUT 0,5 µL
Água DEPC 0,01% q.p. 10 µL
Total 10 µL
Manteve-se a mistura acima a 37°C por 30 minutos, em seguida, adicionou-se 1µL de EDTA (25mM) a cada amostra e a mistura foi mantida a 65°C por 10 min e resfriada a 4°C em banho de gelo.
Desnaturação de RNA
RNA total (tratado) 10 µL
Primer oligo dT 1 µL (0,5 µg )
Total 11 µL
27
Mistura para transcrição reversa
Tampão de enzima (5x) 4,0 µL (1X) MgCl2 (25mM) 2,4 µL (3mM) dNTPs (10mM) 1,0 µL (0,5mM)
RNAse OUT 0,5 µL
Enzima IMPROM II reverse 1,0 µL Água DEPC 0,01% q.p. 20 µL
Total 20 µL
As reações foram conduzidas de acordo com a programa de síntese de cDNA descrito abaixo:
25°C por5 minutos 42°C por 60 minutos
70°C por 15 minutos, inativando a enzima pelo calor.
3.4.3 - Reação em cadeia da polimerase em tempo real
Para a reação em cadeia da polimerase em tempo real os reagentes foram acondicionados em tubos apropriados e adicionados de acordo com os procedimentos abaixo descritos:
Iniciadores Sequências
ENA1 direto 5’- AGTTGGCGGTATTGCTTTTCTG - 3’
ENA1 reverso 5’- TTGAGAGGCCATGACGATGAT - 3’
NHA1 direto 5’- CGCGCCGCCACATTT - 3’
28
Amplificou-se também o gene da actina para ser utilizado como controle endógeno, e para isso os seguintes iniciadores foram utilizados:
Iniciadores Sequências
ACT1 direto 5’- GCCGAAAGAATGCAAAAGGA - 3’
ACT1 reverso 5’- TCTGGAGGAGCAATGATCTTGA - 3’
• 5 µL cDNA
• 5 µL de Iniciadores (direto e reverso) (150 µg/mL ou 20 µL)
• 4 µL DNTP 5 mM
• 10 µL Tampão de Amplificação 10X
• 70,5 µL H2O.
A mistura foi aquecida durante 2 minutos a 94° C e centrifugada rapidamente, antes que 0,5 µL de Taq DNA polimerase 2,5U fosse adicionada. A mistura completa
foi homogeneizada e centrifugada novamente e colocadas no aparelho, utilizando-se o seguinte programa:
As reações foram conduzidas em um Instrumento ABI7500 (Applied Biosystems, Foster City, CA) de acordo com as condições descritas abaixo:
• Elevação da temperatura a 50o C por 2 minutos • 50°C por 2 minutos
• 95o C por 10 minutos
O ciclo foi repetido por 39 vezes. • 94o C por 15 segundos
• 59o C por 1 minuto
Abaixamento da temperatura a 4o C.
29
Para a análise dos resultados, foi utilizado o método comparativo 2^ [-∆ (∆C(T))] descrito por Livak, K. J. e Schmittgen, T. D. ((Livak, K. J. e Schmittgen, T. D., 2001).
3.5 - Medidas do pH interno
O pH interno das cepas, que foram expostas às condições de estresse ácido e adição de NaCl, foi avaliado através da técnica de distribuição do [2-14C] - ácido propiônico entre o interior da célula e o meio extracelular ( Viegas, C. A. e Sá-Correia, I., 1991). As células foram crescidas em 20 mL de meio YPD 4%, até atingirem D.O.
600nm 1,0. Após o crescimento as células foram lavadas duas vezes com YP e
30 3.6 - Conteúdo intracelular de sódio e potássio
As leveduras dos mutantes de S. cerevisiae com diferentes deleções para o
transporte de sódio e a levedura S. boulardii foram crescidas em 50 mL de YP glicose
4%, foram coletadas por centrifugação a 3750 rpm por 5 minutos e expostas às condições de estresse ácido já descritas anteriormente. Alíquotas de 4 mL da suspensão de células foram coletadas por filtração a vácuo em filtros previamente pesados, para que fosse possível determinar o peso seco de células. Paralelamente coletaram-se alíquotas de células expostas e não expostas que foram filtradas e lavadas imediatamente com abundância em solução MgCl2 20mM. Os filtros contendo as células foram submersos em 5 mL de solução HCl 1M e após agitação em vórtex foram incubados a 4°C “overnigth”. Para determinar os níveis de Na+ e K+ das alíquotas foi utilizado Espectrofotômetro de Emissão Atômica Varian AA-20 e solução padrão de Na+ e K+ com concentração 100 µg/mL, para ambos (Rodríguez-Navarro, A. e Ramos, R., 1984) .
Para determinar a taxa de sódio total nas amostras utilizou-se a equação matemática a seguir:
Na+ (nmoles/mg de massa seca) = Leitura x (5 x 10-6) 23
X 1000000
P.S. Sendo que:
a) Leitura = quantidade de sódio expresso em mg/L. b) P.S. = valor de peso seco em g.
31
d) 23 = quantidade de Na+ (em mg) presente em 1 mmol (peso molecular).
e) 1000000 = fator de correção desde que o sódio foi expresso em nmoles/mg de célula.
Para determinar a taxa de potássio total nas amostras utilizou-se a equação matemática a seguir:
K+ (nmoles/mg de massa seca )= Leitura x (5 x 10-6) 39,10
X 1000000
P.S.
Sendo que:
a) Leitura = quantidade de potássio expresso em mg/L. b) P.S. = valor de peso seco em g.
c) 5 x 10-6 = volume do extrato de potássio em litros.
d) 39,10 = quantidade de K+ (em mg) presente em 1 mmol (peso molecular). e) 1000000 = fator de correção desde que o potássio foi expresso em nmoles/mg de
32 3.7 - Atividade H+-ATPásica
As células foram crescidas em meio YPD 4% até D.O.600nm = 1,0, coletadas por centrifugação 3750 rpm e lavadas duas vezes com YP, a 4°C e diluídas em YP na concentração de 75 mg/mL de células para a coleta do controle (tempo 0). Em seguida, a suspensão de células foi centrifugada novamente e diluída para exposição ao estresse ácido acrescido de 85mM de NaCl. As células submetidas ao estresse foram coletadas em alíquotas de 5 mL, nos tempos 30 e 60 minutos, filtradas a vácuo e acondicionadas imediatamente em papel alumínio e congeladas em nitrogênio líquido. As amostras foram estocadas no freezer -80°C para obtenção das membranas citoplasmáticas.
3.7.1 - Obtenção das membranas citoplasmáticas
As amostras congeladas foram transferidas para tubos de ensaio (0,5 mm de diâmetro), contendo 1,5 g de pérolas de vidro, 0,5 mL de tampão de extração (Tris 0,1
M, EDTA 5 mM, β mercaptoetanol 1mM, pH 8,0) e homogeneizadas duas vezes por 90 segundos com intervalos de resfriamento em gelo como descrito por Becher Dos Passos, J. e cols., (1992). Em seguida, foi adicionado 1 mL de tampão de extração (Sorbitol
0.33 M, Tris 0.1 M, EDTA 5 mM, β mercaptoetanol 1 mM, pH 8.0) e as amostras foram novamente homogeneizadas por 30 segundos. Desta forma uma boa parte das células foi lisada, possibilitando o isolamento de frações de membranas.
Para a obtenção das membranas citoplasmáticas foi feito uma centrifugação fracionada do homogenato, a 1.000g por 3 minutos, sendo o sobrenadante resultante centrifugado a 15.000g por 30 minutos. Após a centrifugação o sedimento obtido foi ressuspenso em 1 mL de tampão glicerol (Glicerol 20 %, Tris 10 mM, EDTA 0,1 mM, β mercaptoetanol 1 mM, pH 7,5).
33
membrana citoplasmática da levedura fica retida na interface das duas fases de sacarose (Serrano, R., 1983), portanto, a interface do gradiente foi coletada, diluída em água Mili-Q a 40C e homogeneizada. Esta nova suspensão foi centrifugada a 100.000g por 30 minutos. O sedimento final obtido, contendo a membrana citoplasmática, foi ressuspenso em 300µL de tampão glicerol, sendo utilizado para determinação da atividade H+- ATPásica (Becher Dos Passos, J. e cols., 1992).
3.7.2 - Ensaio da atividade H+-ATPásica
As amostras contendo frações de membrana citoplasmática relativas aos diferentes tempos de incubação foram testadas em relação à atividade H+-ATPásica, em duplicatas. Para cada uma delas, fez-se um branco correspondente a cada amostra, branco em triplicata para o substrato (branco ATP) e um branco geral.
No branco geral foram adicionados 250 µL de tampão de incubação (Hepes 50 mM, NaN3 10mM - que inibe a ATPase mitocondrial e molibdato de amônio 5mM – que inibe fosfatases ácidas, pH 6,5), nos brancos correspondentes as amostras foram
adicionados 210 µL deste tampão. Nos brancos para ATP foram adicionados 220 µL e
nos testes foram adicionados 180 µL do tampão de incubação.
Como já descrito por Dufour, J. P. e Goffeau, A., A., 1978, a atividade ATPásica requer Mg2+, Mn2+ ou Co3+ como cofatores enzimáticos, sendo o íon Mg2+ o mais efetivo. Desta forma, foram adicionados aos tubos testes e aos brancos ATP, 10 µL de uma solução de MgCl2 50mM (concentração final 1mM). E, finalmente foram adicionados 40 µL das amostras, nos testes correspondentes e nos brancos das amostras. As amostras foram pré-incubadas a 30°C, em banho-maria por 5 minutos. Em seguida, iniciou-se a reação pela adição de ATP 25 mM (concentração final 1 mM). Após este período de incubação a reação foi interrompida com 250 µL de ácido
tricloroacético 10% (p/v) e nos tubos brancos ATP foram adicionados 20µL de solução
34
5 mM em todos os tubos, seguidos da adição de 50 µL de reativo de cor (ANSA 2,5 mg, Na2 S2O5 1,5g, Na2 SO3 50 mg, 10 mL de água Milli-Q) recentemente preparado. As amostras foram deixadas a temperatura ambiente por 30 minutos e, posteriormente foi realizada a leitura a 710 nm em um espectrofotômetro (Beckman DU-68) (Becher Dos Passos, J. e cols. 1992). Como padrão utilizou-se Na3HP04.
A atividade específica expressa em µmoles Pi/min./mg proteína foi calculada por meio da seguinte equação:
Abs. 710nm – a1 × 4 × 6,25
Ativ. enz.específica (µmoles Pi min-1 mg-1) = b1 × 1000
Abs. 546nm – a2 × 20 ×10 b2
Sendo que:
a) Abs. 710nm = Valor da absorbância relativa à hidrólise de ATP. b) a1 = Intercepto da curva padrão de fosfato.
c) b1 = Inclinação da curva padrão de fosfato.
d) 4 = Fator de correção (µg fosfato/ 250µL → µg fosfato/mL) desde que proteína foi expressa em mg/mL.
e) 6,25 = Fator de diluição da amostra (40 µL da amostra diluídos para 250 µL final do tampão de incubação).
f) Abs. 546nm = Valor em absorbância referente à dosagem de proteína. g) a2 = Intercepto da curva padrão de proteínas.
h) b2 = Inclinação da curva padrão de proteínas.
i) 20 =Fator de diluição da amostra para dosagem de proteínas (20µL da amostra +
35
j) 10 = Tempo de reação. 3.7.3 - Dosagem de proteína
A dosagem de proteínas foi feita de acordo com o método de Lowry, O. H. modificado (Lowry, O. H. e cols., 1951), utilizando soro albumina bovina como padrão.
3.8 - Determinação do potencial de membrana
O método para determinação do potencial de membrana foi baseado na metodologia descrita por Krasznai, Z. e cols., (1995), com algumas modificações.
Utilizando-se técnicas de citometria de fluxo para detectar a distribuição do marcador bis-oxonol trimetina (1,3-ácido dibutilbarbitúrico) (Di-BaC-4(3)) através da membrana citoplasmática, que obedece a lei de Nernst, na qual explica as alterações do potencial de membrana após aumento ou reduções das concentrações iônicas nos líquidos extra e intracelulares.
3.8.1 - Aquisição e análise dos dados em citômetro de fluxo
As células foram crescidas em 50mL de meio YP glicose 4% a 300 rpm, a 30°C até atingirem D.O.600nm = 1,0. As células foram coletadas e expostas às condições de estresse de acordo com o item 3.3.
36
mesma suspensão de células em tubos de poliestireno 22 x 75 cm (Vacutainer – BD, E.V.A.). Os tubos foram incubados em temperatura ambiente durante 30 minutos ao abrigo da luz. Após incubação, as amostras foram lidas e analisadas em citômetro de fluxo FACS-calibur (Becton Dickinson) utilizando o programa “BD CellQuestTM Pro Software”.
Para determinação do valor do potencial de membrana das células utilizou-se a equação de Nernst, descrita a seguir:
Ѱ= [(RT/F) ln(Di/De)]
Sendo que :
a) Ѱ = Potencial de membrana
b) R = Constante Universal dos Gases – 8,315 J/mol c) T = Temperatura absoluta – 298 K
d) F = Constante de Faraday – 96480 J/Vmol e) Di = Concentração interna de DiBA-C4-(3) f) De = Concentração externa de DiBA-C4-(3)
3.8.2 - Curva padrão de potencial com células fixadas
As cepas de Saccharomyces utilizadas para construção da curva padrão de
DiBA-C4-(3), foram crescidas em 10 mL de YP glicose 4% sob agitação a 30°C até atingirem D.O.600nm = 1,0. Após terem sido coletadas e lavadas com YP, as células foram ressuspensas em PBS 1X filtrado para concentração final de células de 5 x 10-6 células/mL e mantidas em gelo. Para fixação foi adicionada a 10 mL da suspensão de células o mesmo volume de solução de fixação (1M de NaOH, 0,33M de Paraformaldeído, 0,05M cacodilato de sódio e 0,11M de NaCl – pH 7,2). S. cerevisiae
W303 foi fixada em banho de gelo e S. boulardii fixada em temperatura ambiente,
37
essencial para posterior construção da curva padrão de DiBA-C4-(3). A suspensão de células fixadas foi então centrifugada a 3750 rpm por 5 minutos a 4°C, depois de descartado o sobrenadante, foram lavadas duas vezes com PBS1X filtrado e ressuspensas no mesmo tampão conservando a concentração de 5x10-6 células/mL.
Para construção da curva padrão de potencial, adicionou-se diferentes concentrações de DiBA-C4-(3) à suspensão de células previamente fixadas. Os tubos contendo a mistura de DiBA-C4-(3) e células foram incubadas em temperatura ambiente durante 30 minutos ao abrigo da luz. Após incubação, as amostras foram lidas e analisadas em citômetro de fluxo FACS-calibur (Becton Dickinson) utilizando o programa “BD CellQuestTM Pro Software”.
Os pontos da curva padrão de potencial foram feitos em concentrações de 0, 300, 400, 500, 700 e 1000 nM de DiBA-C4-(3) e foram obtidos em triplicatas técnicas.
3.9 - Medida de cálcio celular total
As células de S. cerevisiae e S. boulardii foram crescidas em YP glicose, foram
38
de Ca2+ das alíquotas foi utilizado Espectrofotômetro de Absorção Atômica Varian
AA-20 e solução padrão de Ca2+ com concentração de 1 µg/mL (Miseta, A. e cols., 1999).
Para determinar a taxa de cálcio celular total foi utilizada a equação matemática descrita a seguir:
Ca+2 (mmoles/Kg de massa seca) = Leitura x (5 x 10-3) 40
X 1000
P.S.
Sendo que:
a) Leitura = quantidade de cálcio expresso em mg/L. b) P.S. = valor de peso seco em kg.
c) 5 x 10-3 = volume do extrato de cálcio em litros.
d) 40 = quantidade de Ca2+ (em mg) presente em 1 mmol (peso molecular).