ContentslistsavailableatScienceDirect
International
Journal
of
Developmental
Neuroscience
jo u r n al h om ep a g e :w w w . e l s e v i e r . c o m / l o c a t e / i j d e v n e u
Effects
of
early
or
late
prenatal
immune
activation
in
mice
on
behavioral
and
neuroanatomical
abnormalities
relevant
to
schizophrenia
in
the
adulthood
Vivian
T.
da
Silveira
a,
Daniel
de
Castro
Medeiros
b,
Jivago
Ropke
a,
Patricia
A.
Guidine
b,
Gustavo
H.
Rezende
b,
Marcio
Flavio
D.
Moraes
b,
Eduardo
Mazoni
A.M.
Mendes
b,
Danielle
Macedo
c,
Fabricio
A.
Moreira
a,
Antonio
Carlos
P.
de
Oliveira
a,∗aDepartmentofPharmacology,InstituteofBiologicalSciences,UniversidadeFederaldeMinasGerais,Brazil
bCenterforTechnologyandResearchinMagneto-Resonance(CTPMAG),GraduatePrograminElectricalEngineering–UniversidadeFederaldeMinas
Gerais,Brazil
cDrugResearchandDevelopmentCenter,DepartmentofPhysiologyandPharmacology,MedicalSchool,UniversidadeFederaldoCeará,Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received6December2016
Receivedinrevisedform19January2017 Accepted19January2017
Availableonline22January2017
Keywords:
Schizophrenia Neurodevelopment Prenatalimmuneactivation PolyI:C
Magneticresonanceimaging
a
b
s
t
r
a
c
t
Maternalimmuneactivation(MIA)duringpregnancyinrodentsincreasestheriskoftheoffspringto developschizophrenia-relatedbehaviors,suggestingarelationshipbetweentheimmunesystemandthe braindevelopment.HerewetestedthehypothesisthatMIAinducedbytheviralmimetic polyinosinic-polycytidylicacid(polyI:C)inearlyorlategestationofmiceleadstobehavioralandneuroanatomical disordersintheadulthood.Ongestationaldays(GDs)9or17pregnantdamsweretreatedwithpolyI:Cor salineviaintravenousrouteandtheoffspringbehaviorsweremeasuredduringadulthood.Considering theprogressivestructuralneuroanatomicalalterationsinthebrainofindividualswithschizophrenia,we usedmagneticresonanceimaging(MRI)toperformbrainmorphometricanalysisoftheoffspringaged oneyear.MIAonGD9orGD17ledtoincreasedbasallocomotoractivity,enhancedmotorresponsesto ketamine,apsychotomimeticdrug,andreducedtimespentinthecenterofthearena,suggestingan increasedanxiety-likebehavior.Inaddition,MIAonGD17reducedglucosepreferenceintheoffspring. Noneofthetreatmentsalteredtherelativevolumeofthelateralventricles.However,adecreaseinbrain volume,especiallyforposteriorstructures,wasobservedforone-year-oldanimalstreatedwithpolyI:C comparedwithcontrolgroups.Thus,activationofthematernalimmunesystematdifferentGDslead toneuroanatomicalandbehavioralalterationspossiblyrelatedtothepositiveandnegativesymptoms ofschizophrenia.Theseresultsprovideinsightsonneuroimmunonologicalandneurodevelopmental aspectsofcertainpsychopathologies,suchasschizophrenia.
©2017ISDN.PublishedbyElsevierLtd.Allrightsreserved.
1. Introduction
Schizophreniaisasevere,chronicandmultifactorialpsychiatric disorderthataffectsaround0.3–0.7%ofthepopulation(American PsychiatricAssociation,2013).Severalhypotheseshavebeen pro-posed to identifymechanisms involved in the development of this pathological condition, including the neurodevelopmental hypothesis.Accordingtothishypothesis,maternalimmune acti-vation (MIA),withconsequent induction of neuroinflammatory
∗ Correspondingauthorat:DepartmentofPharmacology,InstituteofBiological Sciences,UniversidadeFederaldeMinasGerais,Av.AntônioCarlos6627,31270-901 BeloHorizonte,MG,Brazil.
E-mailaddress:[email protected](A.C.P.deOliveira).
response in the fetus, would affect neurogenesis, brain devel-opment,and neuronal maturation. Thus, long-termsequelscan interfere negatively in responses to bio-psycho-social stress-ors, predisposing to schizophrenia and other mental illnesses in adulthood (Weinberger, 1987, 1995). Among the biological changeslinkedtoschizophreniaarefacilitationofdopaminergic transmissionwithhyperactivityandenhancedresponsivityto psy-chotomimeticdrugs,anhedonia,anxietyand ventriculardilation (VanandKapur,2009).
Systematicreviewshaveshownrobustevidencebetween pre-natalinfectioninducedbydifferentagentsandadultschizophrenia (Khandakeretal.,2012,2013;Avramopoulosetal.,2015),withan alteredpatternoftheinflammatorymediators(Milleretal.,2011; Gaoetal.,2014;Stuartetal.,2015),aswellasprogressive
neu-http://dx.doi.org/10.1016/j.ijdevneu.2017.01.009
roanatomicalchangesinpatientswiththisdisorder(Lawrieetal., 1997;Olabietal.,2011;Samartzisetal.,2014).However,littleis knownaboutthemolecularmechanismsinvolvedinthisintriguing associationandcontroversyexistsintheliterature.Thishas moti-vatedtheestablishmentofneurodevelopmentalanimalmodelsof schizophrenia,includingtheinduction ofMIAby administering virusorviralmimeticparticles,suchaslipopolysaccharide, turpen-tine,IL-6(Meyer,2014)orpolyinosinic-polycytidylicacid(polyI:C) (Macêdoetal.,2012;Meyer,2014).
Poly I:C is a synthetic analog of double-stranded RNA that mimicscertainvirus,suchasthehumaninfluenzavirus.Itsmost well-knownactionistheactivationofToll-likereceptor3(TLR3), whichincreasestheexpressionofdifferentgenesinvolvedinthe innateimmuneresponse andinterferewithleukocyte function-ing,reducing rollingvelocity, increasingadhesion tothevessel walland facilitating extravasationfrom thecirculation to peri-tonealcavityorothertissues(Meyer,2014;Pircheretal.,2014). Thus,theinflammatory mediatorsproduced inresponse tothis activation,associatedtomorphological(Fatemietal.,2012)and endocrine changes in theplacenta (Hsiao and Patterson 2011), leadtoactivationoftheperipheralandcentralimmunesystem of the fetus (Juckel et al., 2011; Ibi et al., 2011; Fatemi et al., 2012).
Poly I:C neonatal administration in experimental animals induces various biological changes that recapitulates the neu-robiology, symptomatology and epidemiology of schizophrenia (Meyer and Feldon 2012; Forrest et al., 2012).This model has alsobeenpharmacologicallyvalidatedwithclinicallyused antipsy-choticdrugs,suchasclozapine,haloperidolandrisperidone,which reducedtheneuroanatomicalandbehavioralabnormalities result-ingfromMIA(Zuckermanetal.,2003;Ozawaetal.,2006;Meyer et al., 2010; Piontkewitz et al., 2009, 2011, 2012a; Roenker etal.,2011;Richtandet al.,2011;Ribeiroet al.,2013). Numer-ousexperimentalinvestigationssuggest that thepreciseperiod duringwhichprenatalimmuneactivationoccurscaninduce dis-tinctabnormalitieslaterintheadulthood.Forinstance,itseems thatMIAat gestationday9(GD9) causeslong-term alterations thatcorrelatewithpositivesymptomsofschizophrenia,whereas whenchallengedatGD17offspringpresentpredominantly nega-tive/cognitivesymptoms.However,therearesomemissingdata intheliteratureregardingthelong-termconsequencesofprenatal MIA(Macêdoetal.,2012).Forexample,littlehasbeeninvestigated aboutthealterationsbothinearly(GD9)andlate(GD17)gestation inasinglestudy(Meyeretal.,2008;Lietal.,2009,2010).Inaddition, itremainsunclearhowinterventionatspecifictimeperiodsduring pregnancyinfluencethebraindevelopmentandits neuroanatom-icalformationthatcanpredisposetomentaldisorders.
Thus,inordertocontributetoabetterunderstandingofthe influenceofperipheralandcentralinflammationinthe pathogene-sisofschizophrenia,wetestedthehypothesisthattheMIAinduced byexposuretopolyI:CatGD9orGD17inducesdifferentbehavioral andneuroanatomicalabnormalitiesintheadultoffspring.
2. Methods
2.1. Animals
MaleandfemaleC57Bl/6mice(20–25g,6–8weeksofage)were obtainedfromAnimalCareFacilitiesoftheInstituteofBiological Sciences(ICB)−FederalUniversity ofMinasGerais,Brazil. Ani-malswerehousedat25◦Cwith12hlight/darkcycleandfreeaccess
towaterandfood.Allproceduresusedinthisstudyfollowedthe ethicalprinciplesofanimalexperimentationadoptedbytheEthics CommitteeonAnimalUseoftheFederalUniversityofMinasGerais (CEUA−UFMG),andinstitutionallyapprovedunderprotocol
num-ber227/2013.Alleffortsweremadetominimizethenumberof animalsusedandtheirsuffering.
2.2. Breeding
Micewerematedaccordingtothepreviousliterature(Meyer etal.,2008;Bitanihirweetal.,2010).Femaleandmalemicewere maintainedinacageovernight(from18p.m.to8a.m.)in propor-tionofonefemaletoeachmale,andthepresenceofthevaginal plugwasobservedinthenextmorning.Incaseoftheformationof thevaginalplug,thisdaywassetasGD0.
2.3. Activationofthematernalimmunesystem
Eightpregnantdamsweretreatedwithasingleinjectionofpoly I:C(polyinosinic-polycytidylicacidpotassiumsalt;Sigma-Aldrich, SaintLouis,USA)and6pregnantdamsweretreatedwithasingle injectionofsaline(sterilepyrogenfree0.9%NaCl)viaintravenous (i.v.)routeatGD9or17atafinalvolumeof4ml/kg.Thesedayswere chosenduetothecorrespondencetotheearlystagesoffirstandthe secondtrimesterofhumangestation,respectively(Workmanetal., 2013).PolyI:Cwasdissolvedinsterilesalinesolutionontheday ofadministrationandinjectedinthelateraltailveinofthe preg-nantdams.ThedoseofpolyI:C(5mg/kg)waschosenbasedon previousstudies(Meyeretal.,2005;Meyer,2014;Arsenaultetal., 2014).Inordertoperformi.v.injections,femaleswererestrained andthetailwasgentlywarminginwater(40◦C)toinduce
vasodi-lationforabettervisualizationofthevessels.Thesuccessofthe injectionwascheckedinaccordancewiththeitemsdescribedin Bitanihirweetal.(2010).Briefly,thetotalnumberofcirculating white blood cell (WBC) from pregnant females wasperformed exactly24hafteradministrationofpolyI:Corsaline.Blood sam-ples(10l)takenfromthetailwerecollectedanddilutedinratio 1:40withTurk’ssolution(Sigma,St.Louis,MO,USA).Total nucle-atedcellswerecountedinduplicateinahemacytometer(Neubauer chamber)withtheaidofopticalmicroscope,inaccordancewith Thrall(2007).TheresultswereexpressedinWBC/10−6Lofblood. Afterthisprocedure,thefemaleswerehousedindividuallyinan enrichedenvironmentuntilthebirthofoffspring.
2.4. Phenotypecharacterizationoftheoffspring
Twenty-onedaysafterbirth, theoffspring wereweanedand separatedbysexand treatmentgroup,withamaximumoffive animals per cage. For the present study, only the litter males wereevaluated.Malelittermateswereseparatedintothefollowing groups:pre-exposuretopolyI:CatGD9(groupPE9),pre-exposure tosterilesalineatGD9(groupSE9),pre-exposuretopolyI:Cat GD17(groupPE17),pre-exposuretosterilesalineatGD17(group SE17).
Behavioraltestsbeganintheyoung-adultage,fromthetenth weekoflife.Thedecisiontostarttheexperimentsfromthisage wasbasedonpreviousliteraturetomakealongitudinal examina-tioninoffspringexposedtopolyI:Cintheirprenatallife.These studiesdemonstratedthatthemainalterationsareobservedwhen offspringreachlateadolescenceorearlyadulthood, recapitulat-ingclinicalobservationsinpatientswithschizophrenia(Vuillermot etal.,2010;Vernonetal.,2015).
2.5. Experimentalprotocols
2.5.1. Basallocomotionandtimeinthecenterofthearena
Animalsweregentlyplacedatthecenter ofa circulararena (50cmdiameter)enclosedbywalls(40cm)locatedinanisolated room.Thearenawasrecordedbyavideocameracoupledtoa com-puter.Thedistancetraveledandthetimespentinthecentralarea ofthearena(definedasanareawith15cm2)wereautomatically analyzedduring20minbytheAny-maze®software.
2.5.2. Hyperlocomotioninducedbyketamine
Theanimalswerefirsthabituatedinthearenaduring10min andthen receivedan administrationofketamine(60mg/kg)by intraperitoneal route. This dose was established previously by adose-responsecurveperformedbyourgroup.After15minof ketamineadministration,thelocomotionoftheanimalswas mea-suredfor20min.Theanimalswerefilmedandthedistancetraveled isautomaticallyanalyzedbyAny-maze®software.
2.5.3. Sucrosepreferencetest
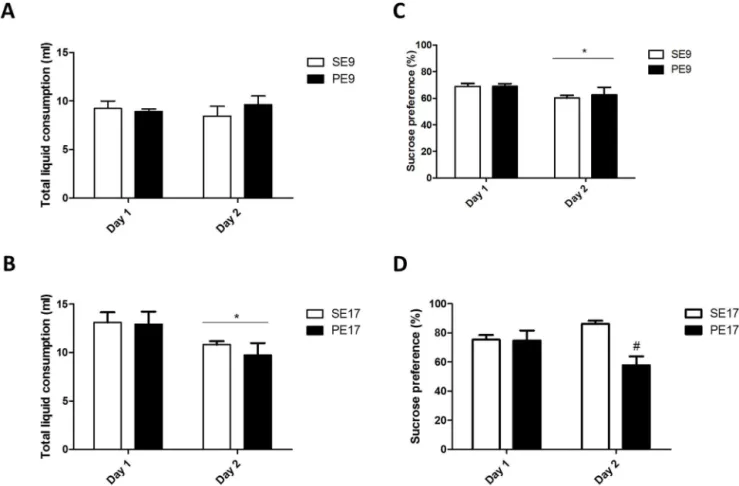
SucrosepreferencetestwasperformedaccordingtoBitanihirwe etal.(2010).Thistestisbasedontheobservationthatrodents typ-icallyprefertoingestsweetwaterinsteadofpurewater,when givingthepossibilityofthischoice.Thereductioninthis prefer-enceischaracterizedasanhedonia.Thus,inordertoevaluatethe possiblechangesofthisinstinctivebehaviorofrodents,the con-sumptionofwateroftheanimalswasmeasuredevery24huntil2 p.m.,for3days(habituationphase).Thisteststartedatthenextday andlastedfor48h(teststage).Duringthetest,theanimalshadfree accesstotwodrinkingtubelocatedatthetopofthecage.Atube wasfilledwithasolutionof0.5%sucrose,theotherwithwater.After 24htofreeexposuretothetubes,thequantityofwaterandsucrose wasmeasured,andthepositionsofthetubeswerechanged.Anew measurementofthevolumeofthesolutioninthetubewasdone after24h.Theresultsofsucrosepreferencewerecalculatedasa percentagescore[sucroseconsumption/(totalliquidconsumption) x100],accordingtoapreviousstudy(Bitanihirweetal.,2010).
2.5.4. Analysisoftheneuroanatomicalabnormalities
ToanalyzetheeffectofMIAexposureatGD9and17on neu-roanatomicalabnormalities in the offspring, we used magnetic resonanceimaging(MRI).Todothis,afterthebehavioral exper-iments,themaleanimalswerereturnedtotheirboxesandkept untiltheybecome1yearold,whentheMRIwasperformed.Asfar asweknow,thisisthefirstworkthatperformedbrain morphome-tricanalysisinthisanimalmodelatthisage.Thisagewaschosen based on epidemiological studies with MRI. MRI was selected becauseitisanon-invasivetechnique.Theanalysiswasconducted in partnership with the Center of Technology and Research in Magneto-Resonance(CTPMAG)ofUFMG,andthemethodologywas chosenaccordingtotheliterature(Lietal.,2009;Gomidesetal., 2014).Magneticresonance imaging experimentswereacquired using4.7TNMRsystem(OxfordSystems)controlledbyaUNITY Inova200imagingconsole(Varian).Theimagingprotocolconsisted ofcoronalT2-weighted(TR=3000ms,TE=50ms)spinecho multi-slicescans,16contiguous1mmthickslices(Fig.6D−9sequential brainimages), in accordancewith previousliterature(Gomides etal.,2014).Mice(n=24)wereanesthetizedwithhalothane(4% induction,1.5%maintenance)togetherwithoxygen(1.5l/min),and thetotalimagingtimewasabout50minpermouse,timerequired toobtaintheimages.Theanestheticandtheoxygenweredelivered viaafacialmasktominimizeartifactmovements.Allbrain mor-phometricmeasurementswereperformedatNIHImageJprogram. Thevolumes(mm3)of ventricles(slices 0.2and
−0.8cm refer-encedfromBregma)andtotalbrain(5sequentialslicesfrom+1.2 to−2.8cmreferencedfromBregma)werecalculatedby summing-uptheareaofeachsection(mm2)multipliedbytheslicethickness (1mm).Inordertoquantifytheventricleenlargement,thedata
Fig.1. EffectofpolyI:Cinjectiononleukocytescountsintheperipheralbloodof pregnantmice.PregnantfemalemiceweretreatedwithpolyI:C(5mg/kg;i.v.)and whitebloodcell(WBC)fromperipheralbloodwerecountedafter24h.Resultsare shownasmean±SEMof6(salinegroup)and8(polyI:Cgroup)animals.***P<0.001 comparedtosalinegroup.
Fig.2. Effectofprenatalimmuneactivationatearlyorlategestation onthe basallocomotoractivityoftheadultoffspring.Adultoffspringfromthe progeni-torexposedtopolyI:Corsalineattheninth(A)orseventeenth(B)gestationalday wereplacedatthearenaandthebasallocomotoractivitywasevaluated.Resultsare shownasmean±SEM.N=6(VehatGD9),8(polyI:CatGD9),5(VehatGD17)and 5(polyI:CatGD17).*P<0.05comparedtorespectivesalinegroup.
weretakenasrelative volume(ventriclevolumenormalizedby totalbrainvolume).
2.6. Statisticalanalysis
Statistical analysis was performed using Prism 5.0 software (GraphPad, CA,USA). The resultswere analyzed withtwo-way ANOVA,consideringprenatalchallenge(vehicleorpolyI:C)and time(GD9orGD17)ofinjectionasexperimentalfactors.Post-hoc comparisonswereperformedwithBonferronitest.
Inordertoperformthemorphometricanalysisofanimalbrains alongtheantero-posterioraxis(fivesequentialslices,highlighted atFig.6Dby*),thedatafromGD9andGD17werepooledforthe vehicle(Veh)andpolyI:C(Poly)groups.Theprenatalchallenge andsliceweretakenasfactorsforthetwo-wayANOVAanalysis and followedby Bonferronipost-hoc comparison test.Thedata arepresentedasmean±standarderrorofthemean(SEM).The significancelevelwassetatp<0.05.
3. Results
3.1. MIAincreasedlocomotoractivityoftheoffspring
MIAinductionbypolyI:Cwasconfirmedbythereductioninthe numberofWBCs(T(15)=13.93;p<0.0001)(Fig.1).
signif-Fig.3. Effectofprenatalimmuneactivationatearlyorlategestationonthe anxiety-likebehavioroftheadultoffspring.Adultoffspringfromtheprogenitorexposedto polyI:Corsalineattheninth(A)orseventeenth(B)gestationaldayswereplaced atthearenaandthetimethatanimalsspentinthecenteroftheapparatuswas evaluated.Resultsareshownasmean±SEM.N=6(VehatGD9),8(polyI:CatGD9), 5(VehatGD17)and5(polyI:CatGD17).*P<0.05and***P<0.001comparedto respectivesalinegroup.
Fig.4. Effectofprenatalimmuneactivationatearlyorlategestationontheadult offspringresponsetoketamine.Adultoffspringfromtheprogenitorexposedto polyI:Corsalineattheninth(A)orseventeenth(B)gestationaldayswereinjected withketamine(60mg/kg,i.p.)andthelocomotoractivitywasevaluated.Resultsare shownasmean±SEM.N=6(VehatGD9),8(polyI:CatGD9),5(VehatGD17)and 5(polyI:CatGD17).*P<0.05and**P<0.01comparedtorespectivesalinegroup.
icant main effect of day of treatment (F(1,20)=0.4793;ns) and interactionbetweenfactors(F(1,20)=0.4285;ns)wasobserved. Fur-thermore,MIAreducedthetimespentinthecenterofthearena, suggestinganxiety-likebehavior inadultoffspring (Fig.3).This effectisrevealedbyeffectofprenatalchallengeinthisparameter (F(1,20)=29.21; p<0.0001). Thisoccurredirrespectively of injec-tions at day 9 or 17, since there was no effect of treatment day(F(1,20)=0.9519;ns).Therewasnointeractionbetweenfactors on this response (F(1,20)=0.6590;ns). In addition, the adminis-trationof ketamineincreasedlocomotion inanimalsfrom both treatmentdays (Fig. 4). Therewas a significantmain effect of typeoftreatment(F(1,20)=13.50;p=0.0015),butnotdayof treat-ment(F(1,20)=0.02;ns).Therewasnointeractionbetweenfactors (F(1,20)=1.036;ns).
3.2. MIAonGD17reducedglucosepreferenceoftheoffspring
InordertoinvestigatewhetherlateMIAcouldinduce anhedo-niainadultoffspring,weperformedthesucrosepreferencetest. AdultoffspringexposedtoMIAatGD9didnotdifferfrom con-trolfortotalliquidconsumption(F(1,18)=0.26;ns).Therewasno differencebetweendaysofliquidconsumption(F(1,18)=0.003;ns) andinteractionbetweengroupanddayfactors(F(1,18)=0.79;ns) (Fig.5A).However,whenexposedatGD17andevaluatedfortotal liquidconsumption,amaineffectofthedayofliquid consump-tion(F(1,18)=6.01;p=0.025)wasobserved,althoughmaineffect ofthetreatment(F(1,18)=0.3;ns)andinteraction(F(1,18)=0.15;ns) (Fig.5B)werenotobserved.ForsucrosepreferenceofGD9groups, there was an effect of the day of consumption (F(1,18)=5.73; p=0.028),althoughnoeffectofthetreatment(F(1,18)=0.017;ns) andinteraction(F(1,18)=0.13;ns)wereobserved(Fig.5C).Onthe
other hand, for sucrose preference of GD17 groups, there was aneffectofthetreatment(F(1,18)=7.27;p=0.01)andinteraction (F(1,18)=6.62;p=0.02),althoughaneffectofthedayofconsumption wasnotobserved(F(1,18)=0.31;ns)(Fig.5D).
3.3. MIAreducedbrainvolumeoftheoffspringagedone-year
Finally,weevaluatedtheeffectsoftheMIAattheearly(DG9) and late gestation(DG17) on brainvolume of theanimals and lateralventriclesrelativevolume(%)withoneyearold,through theinvivoMRItechnique.Thetwo-wayANOVAanalysisoftotal brainvolume (Fig.6A) revealedno interactionbetweenfactors (F(1,17)=1.00,ns),maineffectoftimeofinjection(F(1,17)=0.37,ns) orbetweeneachgroup(Bonferroni’sposthoctestp>0.05). How-ever,itwasdetectedasignificanteffectofthemainfactorprenatal challenge(F(1,17)=13.04,p=0.002),whichindicatesareductionof brainvolumeforanimalschallengedwithPolyI:Ccomparedwith controlgroups.Sincetherewasnoeffectoftimeofinjection(GD9 orGD17),wecombinedtheanimalstoreducetheanalysisonlyto thetreatmentfactor.Thenweanalyzedtheeffectsoftreatmenton thebrainvolumealongtheantero-posterioraxis.This morphomet-ricmeasurements(Fig.6B)alsorevealedadecreaseofbrainvolume forpolyI:Canimals(interactionbetweenfactorsF(4,95)=0.41,ns; mainfactorsliceF(4,95)=216.4,p<0.0001;mainfactorprenatal challengeF(1,95)=23.38,p<0.0001), mainlyfor posterior brain structures(Bonferroni’sposthoctestp=0.009for−2.8slice).As shownatFig.6C,earlyorlateMIAdidnotaltertherelativevolume oftheoffspring ventricles(Fig.6C, two-wayANOVAinteraction betweenfactorsF(1,17)=2.152,ns; main factor time of injection F(1,17)=0.78,ns; mainfactor prenatal challengeF(1,17)=1.23,ns; Bonferroni’sposthoctestp>0.05).
4. Discussion
Inthepresent study,wedemonstratedthatMIAexposurein twospecificgestationaldaysleadstobehavioraland neuroanatom-icalchangesintheoffspringintheiradultlifeandinagedmice, respectively.ConfirmingthatpolyI:CdidinduceMIA,weobserved thatwhitebloodcellsweresignificantlydecreased24hafter injec-tion.Thisisinaccordance withPircheretal.(2014).Thisresult couldbeexplainedbytheproinflammatoryeffectsthatthe syn-theticanalogueofviraldsRNAplaysonendothelialcells,suchas reductioninleukocyterollingvelocity,increaseinleukocyte adhe-siontothevesselwall,andincreasedextravasationofleukocytes fromthecirculationtoperitonealcavityorothertissuesconditions tightlyimplicatedinsystemicinflammation(Pircheretal.,2014).
endopheno-Fig.5.Effectofprenatalimmuneactivationatearlyorlategestationonanhedonicbehavioroftheadultoffspring.AdultoffspringfromtheprogenitorexposedtopolyI:Cor salineattheninth(AandB)orseventeenth(CandD)gestationaldayswereplacedindividuallyoncagesovernightandasucrosesolutionandwaterweremadeavailablead libitumevery24h.Anhedonicbehaviorwasevaluatedusing(AandC)thetotalofvolumefluids(waterandsucrosesolution)consumptionand(BandD)percentagescoreof sucroseconsumptionontwoconsecutivesdays.Resultsareshownasmean±SEM.N=6(VehatGD9),5(polyI:CatGD9),5(VehatGD17)and6(polyI:CatGD17).*P<0.05 withrespecttoday1groups;#P<0.05comparedtorespectivesalinegroup.
typepredictiveofschizophrenic-typebehaviors(Zuckermanand Weiner,2005;Zavitsanouetal.,2014).However,previousstudies showeddifferentresultsinregardstotheeffectsofNMDA recep-torblockers.Missaultetal.(2014)andMeehanetal.(2016)did notobserveanyeffectsofMK-801inthelocomotoractivityinthe offspringofdamsinjectedwithpolyI:Cincomparisonwith con-trols.Besides,areducedlocomotoractivityoftheoffspringfrom progenitorsinjectedwithpolyI:Cwasalsoreported(Lietal.,2014). Inaddition,inordertoanalyzeanhedonicbehavior,aneffect possiblyrelatedtothenegativesymptomsofschizophrenia,we usedthesamemethodologyofBitanihirweetal.(2010). Accord-ingtothisauthor,thesucrosepreferencetestprovidesasensitive assessment of anhedonicaspects of food.In thepresent study, althoughwe didnot findaneffectofpolyI:Cinjected onGD9, wedidobserveareductioninthesucrosepreferenceonthe sec-onddayoftestbetweenPE17incomparisonwithSE17.Similarto ourresults,Bitanihirweetal.(2010)havealsodemonstratedthat theGD17grouptreatedwithpolyI:Chavereducedpreferencefor sucrose.Onepossiblereasonwhyanhedoniawasevidentonlyin theseconddayofthetestmightbethatpolyI:Cdidnotreducethe motivationtoexploreanovelstimulus(i.e.,sucroseonday1).Only thesecondday,whensucroseisanalreadyfamiliarstimulusand noveltyisnolongerafactor,couldanhedoniabeobserved.
Although the neurochemical basis that mediate the altered behaviorsarenotknow,itispossiblethatMIAresultsinanimpaired dopaminergictransmission.Ithasbeensuggestedthat hyperacti-vationinthesubcorticalmesolimbicdopaminergicsystem,which couldbeassociatedwiththepositivesymptoms,anda hypofunc-tioninthemesocorticalprojectionstotheprefrontalcortex,which
couldprecipitatethenegativesymptoms, arecorebrain abnor-malitiesinschizophrenia(MeyerandFeldon,2009).Inthissense, hyperlocomotionandanhedoniaareassociatedwithanincrease andadecreaseinthedopaminergicsystem,respectively(Grace, 2016).Indeed, thedysfunction in this neurotransmitter system hasbeendescribedintheMIAmodel(Eylesetal.,2012). Prena-talimmuneactivationinducedbyLPSorpolyI:Cmayalter the mesolimbicdopaminergicsystembyincreasingthe immunoreac-tivityoftyrosinehydroxylaseinthenucleusaccumbens(Meyerand Feldon,2009).Besides,MIAreducestheexpressionofD1RandD2R intheprefrontalcortexintheoffspringatadulthood(Meyerand Feldon,2009).Thus, thismodelrecapitulatestheneurochemical andmolecularalterationproposedtounderliecertainsymptoms ofschizophrenia.
Fig.6. Effectofprenatalimmuneactivationatearlyorlategestationonneuroanatomicalstructuresofadultoffspring.TotalbrainvolumeforVehandPolygroupsatthe ninth(GD9)orseventeenth(GD17)gestationaldays(A).Anterior-posteriorbrainvolumeanalysisforpooledVehandPolygroups.Theinsetdepictsillustrativeslicesfrom +1.2to−2.8referencedfromBregma(B).Relativelateralventriclevolume,whichvalueistheratioofeachventriclevolumetowholebrainvolume(C).Representative sequentialslicesfrom+3.2to−4.8mmreferencedfromBregma.The*denotestheanterior-posteriorslicesusedformeasurementoftotalbrainvolume(D).Resultsare shownasmean±SEM.N=5(VehatGD9),8(polyI:CatGD9),4(VehatGD17)and4(polyI:CatGD17).**P<0.01,comparedtorespectivesalinegroup.
Olabietal.,2011;Chiapponietal.,2013)andduetotheabsence ofstudiesthat investigatedthis parameterinaged animals,we decidedtoevaluatewhetheranenlargementofthelateral ven-triclewouldbeobservedinoldanimals,asobservedinhumans. Inthepresentstudy,weusedthesamemethodologyofLietal. (2009).Theseauthors,aswellasothers(Piontkewitzetal.,2009, 2011,2012b)demonstratesignificantenlargementoflateral ven-triclesinoffspringthatprenatalimmunechallengewasperformed inearlygestation.Wedidnotobservechangesinthevolumeof thelateralventricleof theadultoffspring ofanygroups. These discrepanciesmightbeduetothedaysofmaternalimmune acti-vationandtheageusedtoevaluatetheoffspring.Itisknownthat differentmoments of gestation are correlated todifferent data foundintheliterature(Macêdoetal.,2012).Besides,Piontkewitz etal.(2009,2011,2012b)alsousedratsintheexperiments,what couldalsoleadtodifferentresults. Finally,consideringthatthe animalsagedone-yearareconsideredoldanimals,itispossible thatthedifferencesobservedinearlyagesarenotevident any-more.
Ontheotherhand,MIAatGD9orGD17ledtoadecreasein theposteriorbrainvolumeintheoffspringatone-yearold.The neuroanatomicalabnormalitiesobservedbetweenPE9and PE17 indicatesthatMIAindifferentgestationaltimescancompromise theintegrityofthecomplexneuronalcircuitofoffspring.The alter-ationsintheposteriorregionsobservedinthePE9andPE17could partiallyexplainthebehavioraldeficitsobservedintheanimals.As farasweknow,thisisthefirstworkthatperformedbrain mor-phometricanalysisinthisanimalmodelatthisage.Weconsider thisnewknowledgeanimportantresearchobjective,inorderto increasethevalidityandreliabilityofthepredictivevalueofthis animalmodel.
Althoughmuchefforthasbeenmadetounderstandthe com-plexfactors involved in schizophrenia, many questions remain unanswered. In this sense, research on the neurobiology of schizophrenia has increasingly focused on neurodevelopmental aspectsanditsrelationtopro-inflammatorysignals.Cytokines pro-ducedduringmaternalinfectioncanleadtolong-termeffectson fetalsomaticcellsthroughinteractionwithepigeneticmachinery (Bale,2015).Therefore,pregnancyhasbeenoneofthemost stud-iedwindowsofsusceptibilitysincethisisacriticaltimeinneuronal programming.Thus,epidemiologicalstudiesassociatedwith ani-malmodelsareabletodemonstratephylogeneticequivalenceof animalandhumanbehaviorandcanprovideanimportantwayto understandthisdisorder.
Inconclusion,wedemonstratedthatprenatalimmune activa-tioninfluencescertainbehavioralandneuroanatomicalparameters in adult mice. Although the precise mechanisms remain tobe investigated,thisstudyreinforcestheimportanceof neurodevel-opmentalmodelsfor investigating behavioral responsesrelated toschizophreniaandsuggeststhattheoutcomemaydependon interferencesatspecificgestationaldays.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
EMAMMandMFDMarerecipientsofCNPqfellowships.Theauthors aregratefultoMr.GilmarTeodoroBatistaforthetechnicalsupport duringthebreedingoftheanimals.
References
Achim,A.M.,etal.,2011.Howprevalentareanxietydisordersinschizophrenia?A meta-analysisandcriticalreviewonasignificantassociation.Schizophr.Bull. 37(July(4)),811–821(ISSN1745-1701.Availableat:<https://www.ncbi.nlm. nih.gov/pubmed/19959704>).
AmericanPsychiatricASSOCIATION(APA),2013.DiagnosticandStatisticalManual ofMentalDisorders−DSM-5,5thed.APA,Washington.
Arnone,D.,etal.,2009.Magneticresonanceimagingstudiesinbipolardisorder andschizophrenia:meta-analysis.Br.J.Psychiatry195(September(3)), 194–201(ISSN1472-1465.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/19721106>).
Arsenault,D.,etal.,2014.ThedifferenteffectsofLPSandpolyI:Cprenatalimmune challengesonthebehavior,developmentandinflammatoryresponsesin pregnantmiceandtheiroffspring.BrainBehav.Immun.38(May),77–90(ISSN 1090–2139.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/24384468>). Avramopoulos,D.,etal.,2015.Infectionandinflammationinschizophreniaand
bipolardisorder:agenomewidestudyforinteractionswithgeneticvariation. PLoSOne10(3),e0116696(ISSN1932–6203.Availableat:<http://www.ncbi. nlm.nih.gov/pubmed/25781172>).
Bale,T.L.,2015.Epigeneticandtransgenerationalreprogrammingofbrain development.Nat.Rev.Neurosci.16(June(6)),332–344(ISSN1471-0048. Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/25921815>). Bitanihirwe,B.K.,etal.,2010.Lateprenatalimmuneactivationinmiceleadsto
behavioralandneurochemicalabnormalitiesrelevanttothenegative symptomsofschizophrenia.Neuropsychopharmacology35(November(12)), 2462–2478(ISSN1740-634X.Availableat:<http://www.ncbi.nlm.nih.gov/ pubmed/20736993>).
Cahn,W.,etal.,2009.Psychosisandbrainvolumechangesduringthefirstfive yearsofschizophrenia.Eur.Neuropsychopharmacol.19(February(2)), 147–151(ISSN1873–7862.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/19056248>).
Canetta,S.,etal.,2016.Maternalimmuneactivationleadstoselectivefunctional deficitsinoffspringparvalbumininterneurons.Mol.Psychiatry21(July(7)), 956–968(ISSN1476–5578.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/26830140>).
Chiapponi,C.,etal.,2013.Age-relatedbraintrajectoriesinschizophrenia:a systematicreviewofstructuralMRIstudies.PsychiatryRes.214(November (2)),83–93(ISSN1872–7123.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/23972726>).
Eyles,D.,Feldon,J.,Meyer,U.,2012.Schizophrenia:doallroadsleadtodopamine oristhiswheretheystart?Evidencefromtwoepidemiologicallyinformed developmentalrodentmodels.TranslPsychiatry2(February),e81(ISSN 2158–3188.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 22832818>).
Faludi,G.,Mirnics,K.,2011.Synapticchangesinthebrainofsubjectswith schizophrenia.Int.J.Dev.Neurosci.29(May(3)),305–309(ISSN1873-474X. Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/21382468>). Fatemi,S.H.,etal.,2012.Theviraltheoryofschizophreniarevisited:abnormal
placentalgeneexpressionandstructuralchangeswithlackofevidencefor H1N1viralpresenceinplacentaeofinfectedmiceorbrainsofexposed offspring.Neuropharmacology62(March(3)),1290–1298(ISSN1873–7064. Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/21277874>). Forrest,C.M.,etal.,2012.PrenatalactivationofToll-likereceptors-3by
administrationoftheviralmimeticpoly(I:C)changessynapticproteins, N-methyl-D-aspartatereceptorsandneurogenesismarkersinoffspring.Mol Brain5,22(ISSN1756–6606.Availableat:<http://www.ncbi.nlm.nih.gov/ pubmed/22681877>).
Gao,L.,etal.,2014.Associationofinterleukin-10polymorphismswith schizophrenia:ameta-analysis.PLoSOne9(3),e90407(ISSN1932–6203. Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/24603720>). Gogtay,N.,etal.,2008.Three-dimensionalbraingrowthabnormalitiesin
childhood-onsetschizophreniavisualizedbyusingtensor-based morphometry.Proc.Natl.Acad.Sci.U.S.A.105(October(3)),15979–15984 (ISSN1091–6490.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 18852461>).
Gomides,L.F.,etal.,2014.Murinemodeltostudybrain,behaviorandimmunity duringhepaticencephalopathy.WorldJ.Hepatol.6(April(4)),243–250(ISSN 1948–5182.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/24799993>). Grace,A.A.,2016.Dysregulationofthedopaminesysteminthepathophysiologyof
schizophreniaanddepression.Nat.Rev.Neurosci.17(August(8)),524–532 (ISSN1471-0048.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 27256556>).
Haijma,S.V.,etal.,2013.Brainvolumesinschizophrenia:ameta-analysisinover 18000subjects.Schizophr.Bull.39(September(5)),1129–1138(ISSN 1745-1701.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 23042112>).
Howes,O.,Mccutcheon,R.,Stone,J.,2015.Glutamateanddopaminein schizophrenia:anupdateforthe21stcentury.J.Psychopharmacol.29
(February(2)),97–115(ISSN1461–7285.Availableat:<https://www.ncbi.nlm. nih.gov/pubmed/25586400>).
Hsiao,E.Y.,Patterson,P.H.,2011.Activationofthematernalimmunesystem inducesendocrinechangesintheplacentaviaIL-6.BrainBehav.Immun.25 (May(4)),604–615(ISSN1090–2139.Availableat:<http://www.ncbi.nlm.nih. gov/pubmed/21195166>).
Ibi,D.,etal.,2011.PolyI:C-inducedneurodevelopmentalanimalmodelfor schizophrenia.NihonShinkeiSeishinYakurigakuZasshi31(November(5–6)) (ISSN1340–2544.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/ 22256608>).
Juckel,G.,etal.,2011.Microglialactivationinaneuroinflammationalanimal modelofschizophrenia–apilotstudy.Schizophr.Res.131(September(1–3)), 96–100(ISSN1573–2509.Availableat:<http://www.ncbi.nlm.nih.gov/ pubmed/21752601>).
Khandaker,G.M.,etal.,2012.Childhoodinfectionandadultschizophrenia:a meta-analysisofpopulation-basedstudies.Schizophr.Res.139(August(1–3)), 161–168(ISSN1573–2509.Availableat:<http://www.ncbi.nlm.nih.gov/ pubmed/22704639>).
Khandaker,G.M.,etal.,2013.Prenatalmaternalinfection,neurodevelopmentand adultschizophrenia:asystematicreviewofpopulation-basedstudies.Psychol. Med.43(February(2)),239–257(ISSN1469–8978.Availableat:<http://www. ncbi.nlm.nih.gov/pubmed/22717193>).
Lawrie,S.M.,etal.,1997.Qualitativecerebralmorphologyinschizophrenia:a magneticresonanceimagingstudyandsystematicliteraturereview. Schizophr.Res.25(May(2)),155–166(ISSN0920-9964.Availableat:<http:// www.ncbi.nlm.nih.gov/pubmed/9187015>).
Lazar,N.L.,Neufeld,R.W.,Cain,D.P.,2011.Contributionofnonprimateanimal modelsinunderstandingtheetiologyofschizophrenia.J.PsychiatryNeurosci. 36(July(4)),E5–E29(ISSN1488–2434.Availableat:<https://www.ncbi.nlm. nih.gov/pubmed/21247514>).
Levman,J.,Takahashi,E.,2015.Multivariateanalysesappliedtofetal,neonataland pediatricMRIofneurodevelopmentaldisorders.NeuroimageClin.9,532–544 (ISSN2213-1582.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 26640765>).
Li,Q.,etal.,2009.Prenatalimmunechallengeisanenvironmentalriskfactorfor brainandbehaviorchangerelevanttoschizophrenia:evidencefromMRIina mousemodel.PLoSOne4(7)(ISSN1932–6203.Availableat:<http://www. ncbi.nlm.nih.gov/pubmed/19629183>).
Li,Q.,etal.,2010.Voxel-basedanalysisofpostnatalwhitemattermicrostructurein miceexposedtoimmunechallengeinearlyorlatepregnancy.Neuroimage52 (August(1))(ISSN1095–9572.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/20399275>).
Li,W.Y.,Chang,Y.C.,Lee,L.J.,2014.Prenatalinfectionaffectstheneuronal architectureandcognitivefunctioninadultmice.Dev.Neurosci.36(5), 359–370(ISSN1421–9859.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/24942872>).
Macêdo,D.S.,etal.,2012.Animalmodelsofprenatalimmunechallengeandtheir contributiontothestudyofschizophrenia:asystematicreview.Braz.J.Med. Biol.Res.45(March(3)),179–186(ISSN1414-431X.Availableat:<http:// www.ncbi.nlm.nih.gov/pubmed/22392187>).
Marsman,A.,etal.,2013.Glutamateinschizophrenia:afocusedreviewand meta-analysisof1H-MRSstudies.Schizophr.Bull.39(January(1))(ISSN 1745-1701.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/21746807>). Meehan,C.,etal.,2016.Effectsofimmuneactivationduringearlyorlategestation onschizophrenia-relatedbehaviourinadultratoffspring.BrainBehav.Immun. (ISSN1090–2139.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 27423491>)[Epubaheadofprint].
Meyer,U.,Feldon,J.,2009.Prenatalexposuretoinfection:aprimarymechanism forabnormaldopaminergicdevelopmentinschizophrenia.
Psychopharmacology(Berl.)206(November(4)),587–602(ISSN1432–2072. Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/19277608>). Meyer,U.,Feldon,J.,2012.Topoly(I:C)ornottopoly(I:C):advancingpreclinical
schizophreniaresearchthroughtheuseofprenatalimmuneactivationmodels. Neuropharmacology62(March(3)),1308–1321(ISSN1873–7064.Available at:<http://www.ncbi.nlm.nih.gov/pubmed/21238465>).
Meyer,U.,etal.,2005.Towardsanimmuno-precipitatedneurodevelopmental animalmodelofschizophrenia.Neurosci.Biobehav.Rev.29(6),913–947(ISSN 0149-7634.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/15964075>). Meyer,U.,etal.,2008.Adultbrainandbehavioralpathologicalmarkersofprenatal
immunechallengeduringearly/middleandlatefetaldevelopmentinmice. BrainBehav.Immun.22(May(4)),469–486(ISSN1090–2139.Availableat:< http://www.ncbi.nlm.nih.gov/pubmed/18023140>).
Meyer,U.,etal.,2010.Chronicclozapinetreatmentimprovesprenatal infection-inducedworkingmemorydeficitswithoutinfluencingadult hippocampalneurogenesis.Psychopharmacology(Berl.)208(March(4)), 531–543(ISSN1432–2072.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/20041229>).
Meyer,U.,2014.Prenatalpoly(I:C)exposureandotherdevelopmentalimmune activationmodelsinrodentsystems.Biol.Psychiatry75(February(4)), 307–315(ISSN1873–2402.Availableat:<http://www.ncbi.nlm.nih.gov/ pubmed/23938317>).
Miller,B.J.,etal.,2011.Meta-analysisofcytokinealterationsinschizophrenia: clinicalstatusandantipsychoticeffects.Biol.Psychiatry70(October(7))(ISSN 1873–2402.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/21641581>). Missault,S.,etal.,2014.Theriskforbehaviouraldeficitsisdeterminedbythe
neurodevelopmentalmodel.BrainBehav.Immun.42(November),138–146 (ISSN1090–2139.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 24973728>).
Olabi,B.,etal.,2011.Arethereprogressivebrainchangesinschizophrenia?A meta-analysisofstructuralmagneticresonanceimagingstudies.Biol. Psychiatry70(July(1)),88–96(ISSN1873–2402.Availableat:<http://www. ncbi.nlm.nih.gov/pubmed/21457946>).
Ozawa,K.,etal.,2006.Immuneactivationduringpregnancyinmiceleadsto dopaminergichyperfunctionandcognitiveimpairmentintheoffspring:a neurodevelopmentalanimalmodelofschizophrenia.Biol.Psychiatry59 (March(6)),546–554(ISSN0006-3223.Availableat:<https://www.ncbi.nlm. nih.gov/pubmed/16256957>).
Piontkewitz,Y.,Assaf,Y.,Weiner,I.,2009.Clozapineadministrationinadolescence preventspostpubertalemergenceofbrainstructuralpathologyinananimal modelofschizophrenia.Biol.Psychiatry66(December(11)),1038–1046(ISSN 1873–2402.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/
19726031>).
Piontkewitz,Y.,Arad,M.,Weiner,I.,2011.Risperidoneadministeredduring asymptomaticperiodofadolescencepreventstheemergenceofbrain structuralpathologyandbehavioralabnormalitiesinananimalmodelof schizophrenia.Schizophr.Bull.37(November(6)),1257–1269(ISSN 1745-1701.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 20439320>).
Piontkewitz,Y.,etal.,2012a.Effectsofrisperidonetreatmentinadolescenceon hippocampalneurogenesis,parvalbuminexpression,andvascularization followingprenatalimmuneactivationinrats.BrainBehav.Immun.26 (February(2)),353–363(ISSN1090–2139.Availableat:<https://www.ncbi. nlm.nih.gov/pubmed/22154704>).
Piontkewitz,Y.,Arad,M.,Weiner,I.,2012b.Tracingthedevelopmentofpsychosis anditsprevention:whatcanbelearnedfromanimalmodels.
Neuropharmacology62(March(3)),1273–1289(ISSN1873–7064.Available at:<https://www.ncbi.nlm.nih.gov/pubmed/21703648>).
Pircher,J.,etal.,2014.HepatitisCvirusinducedendothelialinflammatory responsedependsonthefunctionalexpressionofTNF␣receptorsubtype2. PLoSOne9(11),e113351(ISSN1932–6203.Availableat:<http://www.ncbi. nlm.nih.gov/pubmed/25419735>).
Ratnayake,U.,etal.,2014.PrenatalexposuretotheviralmimeticpolyI:Calters fetalbraincytokineexpressionandpostnatalbehaviour.Dev.Neurosci.36(2), 83–94(ISSN1421–9859.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/24863806>).
Ribeiro,B.M.,etal.,2013.Evidencesforaprogressivemicroglialactivationand increaseiniNOSexpressioninratssubmittedtoaneurodevelopmentalmodel ofschizophrenia:reversalbyclozapine.Schizophr.Res.151(December(1–3)), 12–19(ISSN1573–2509.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/24257517>).
Richtand,N.M.,etal.,2011.Effectsofrisperidoneandpaliperidonepre-treatment onlocomotorresponsefollowingprenatalimmuneactivation.J.Psychiatr.Res. 45(September(9)),1194–1201(ISSN1879-1379.Availableat:<https://www. ncbi.nlm.nih.gov/pubmed/21440257>).
Roenker,N.L.,etal.,2011.Effectofpaliperidoneandrisperidoneonextracellular glutamateintheprefrontalcortexofratsexposedtoprenatalimmune activationorMK-801.Neurosci.Lett.500(August(3)),167–171(ISSN 1872–7972.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 21699956>).
Samartzis,L.,etal.,2014.Whitematteralterationsinearlystagesofschizophrenia: asystematicreviewofdiffusiontensorimagingstudies.J.Neuroimaging24 (March–April(2)),101–110(ISSN1552–6569.Availableat:<http://www.ncbi. nlm.nih.gov/pubmed/23317110>).
Steeds,H.,Carhart-Harris,R.L.,Stone,J.M.,2015.Drugmodelsofschizophrenia. Ther.Adv.Psychopharmacol.5(February(1)),43–58(ISSN2045.Availableat: <https://www.ncbi.nlm.nih.gov/pubmed/25653831>).
Steen,R.G.,etal.,2006.Brainvolumeinfirst-episodeschizophrenia:systematic reviewandmeta-analysisofmagneticresonanceimagingstudies.Br.J. Psychiatry188(June),510–518(ISSN0007-1250.Availableat:<https://www. ncbi.nlm.nih.gov/pubmed/16738340>).
Stuart,M.J.,Singhal,G.,Baune,B.T.,2015.Systematicreviewoftheneurobiological relevanceofchemokinestopsychiatricdisorders.Front.Cell.Neurosci.9,357 (ISSNAvailableat:<http://www.ncbi.nlm.nih.gov/pubmed/26441528>). Thrall,M.A.,2007.Hematologiaebioquímicaclínicaveterinária.Roca,SãoPaulo
(592p.).
VanHaren,N.E.,etal.,2008.Progressivebrainvolumelossinschizophreniaover thecourseoftheillness:evidenceofmaturationalabnormalitiesinearly adulthood.Biol.Psychiatry63(January(1)),106–113(ISSN1873–2402. Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/17599810>). VanOS,J.,Kapur,S.,2009.Schizophrenia.Lancet374(August(9690)),635–645
(ISSN1474-547X.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 19700006>).
Vernon,A.C.,etal.,2015.Longitudinalinvivomaturationalchangesofmetabolites intheprefrontalcortexofratsexposedtopolyinosinic-polycytidylicacidin utero.Eur.Neuropsychopharmacol.25(December(12)),2210–2220(ISSN 1873–7862.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/26475576>). Vorhees,C.V.,etal.,2015.Prenatalimmunechallengeinrats:effectsof
polyinosinic-polycytidylicacidonspatiallearning,prepulseinhibition, conditionedfear,andresponsestoMK-801andamphetamine.Neurotoxicol. Teratol.47(January–February),54–65(ISSN1872–9738.Availableat:<https:// www.ncbi.nlm.nih.gov/pubmed/25450663>).
Vuillermot,S.,etal.,2010.Alongitudinalexaminationoftheneurodevelopmental impactofprenatalimmuneactivationinmicerevealsprimarydefectsin dopaminergicdevelopmentrelevanttoschizophrenia.J.Neurosci.30(January (4)),1270–1287(ISSN1529–2401.Availableat:<https://www.ncbi.nlm.nih. gov/pubmed/20107055>).
Weinberger,D.R.,1987.Implicationsofnormalbraindevelopmentforthe pathogenesisofschizophrenia.Arch.Gen.Psychiatry44(July(7)),660–669 (ISSN0003-990X.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/ 3606332>).
Weinberger,D.R.,1995.Fromneuropathologytoneurodevelopment.Lancet346 (August(8974)),552–557(ISSN0140-6736.Availableat:<http://www.ncbi. nlm.nih.gov/pubmed/7544856>).
Workman,A.D.,etal.,2013.Modelingtransformationsofneurodevelopmental sequencesacrossmammalianspecies.J.Neurosci.33(April(17)),7368–7383 (ISSN1529–2401.Availableat:<http://www.ncbi.nlm.nih.gov/pubmed/ 23616543>).
Yee,N.,etal.,2011.Differentialeffectsofmaternalimmuneactivationandjuvenile stressonanxiety-likebehaviourandphysiologyinadultrats:noevidencefor thedouble-hithypothesis.Behav.BrainRes.224(October(1)),180–188(ISSN 1872–7549.Availableat:<https://www.ncbi.nlm.nih.gov/pubmed/ 21679729>).
Zavitsanou,K.,etal.,2014.Effectofmaternalimmuneactivationonthe kynureninepathwayinpreadolescentratoffspringandonMK801-induced hyperlocomotioninadulthood:ameliorationbyCOX-2inhibition.BrainBehav. Immun.41(October),173–181(ISSN1090–2139.Availableat:<https://www. ncbi.nlm.nih.gov/pubmed/24878170>).
Zuckerman,L.,Weiner,I.,2005.Maternalimmuneactivationleadstobehavioral andpharmacologicalchangesintheadultoffspring.J.Psychiatr.Res.39(May (3)),311–323(ISSN0022-3956.Availableat:<https://www.ncbi.nlm.nih.gov/ pubmed/15725430>).