Parental legacy in insects: variation of transgenerational immune priming during offspring development.
Texto
Imagem
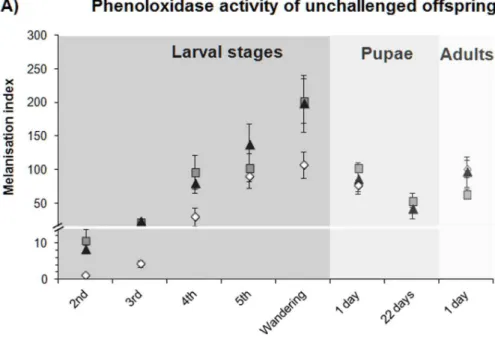
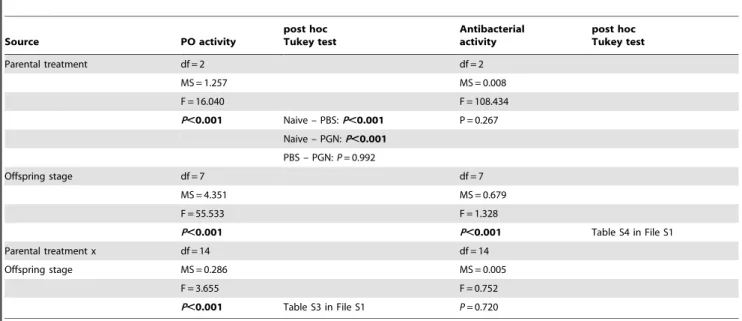
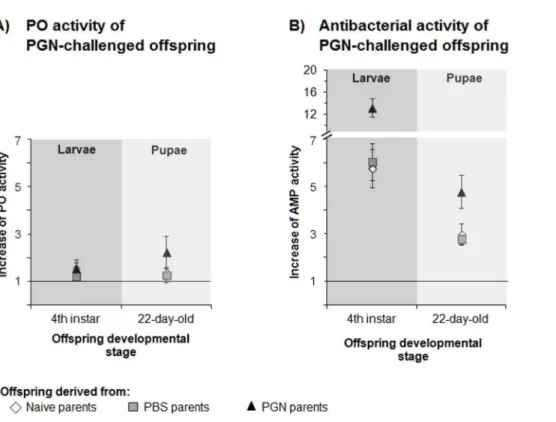
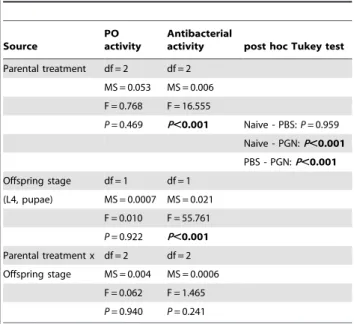
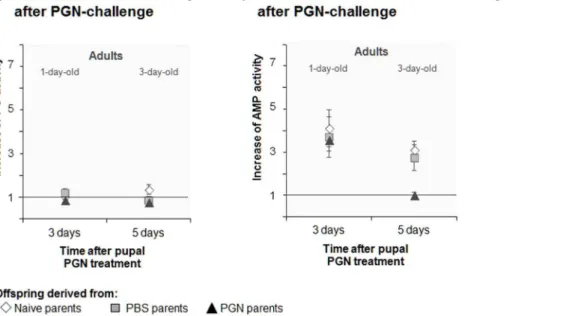
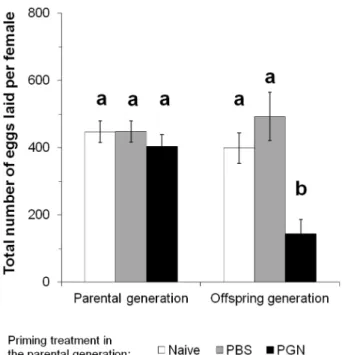
Documentos relacionados
H„ autores que preferem excluir dos estudos de prevalˆncia lesŽes associadas a dentes restaurados para evitar confus‚o de diagn€stico com lesŽes de
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
O consultor deverá informar o cliente, no momento do briefing inicial, de quais as empresas onde não pode efetuar pesquisa do candidato (regra do off-limits). Ou seja, perceber
Effect of prenatal immune activation at early or late gestation on the anxiety- like behavior of the adult offspring.. Adult offspring from the progenitor exposed to poly I:C or
The objective of this study was to evaluate the genetic diversity and the parental contribution of Piaractus mesopotamicus in the production of offspring in the
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
C’est ce que fit Vati- can II dans Gaudium et spes dont nous fêtons cette année le 50 ème
