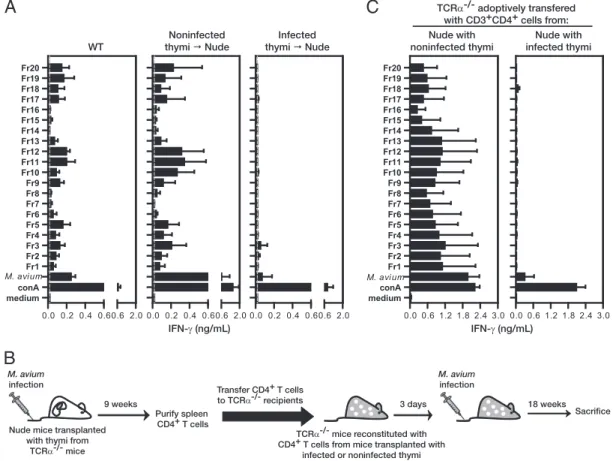
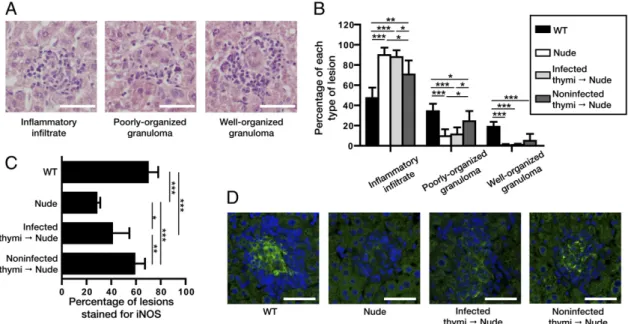
Dissemination of Mycobacteria to the Thymus Renders Newly Generated T Cells Tolerant to the Invading Pathogen
Texto
Imagem
Documentos relacionados
From the 12 studies included (13 entries) in this meta-analysis, they all started from the same research assumption, in which the elderly with classifications according to the
In this connection, the present study aims to present an epidemiological analysis of accidents involving scorpions in Brazil, in the period from 2000 to
Para tanto foi realizada uma pesquisa descritiva, utilizando-se da pesquisa documental, na Secretaria Nacional de Esporte de Alto Rendimento do Ministério do Esporte
Os inquiridos mostram que não compram produtos ou serviços apenas porque são falados por influenciadores digitais pelos quais eles nutrem confiança, o que mostra que, mesmo
Therefore, both the additive variance and the deviations of dominance contribute to the estimated gains via selection indexes and for the gains expressed by the progenies
Os resultados evidenciaram não só uma maior dificuldade na produção de palavras polissilábicas nos dois grupos (alunos com audição normal e alunos com
Objetivou-se com esse trabalho, avaliar o desempenho produtivo de caprinos (Capra hircus) da raça Canindé submetidos a um sistema de exploração misto em regime de manejo
This study aimed at assessing the following in healthy adults: (1) the distribution of aortic PWV values, as well as the diameter, IMT and relative distensibility of the carotid
