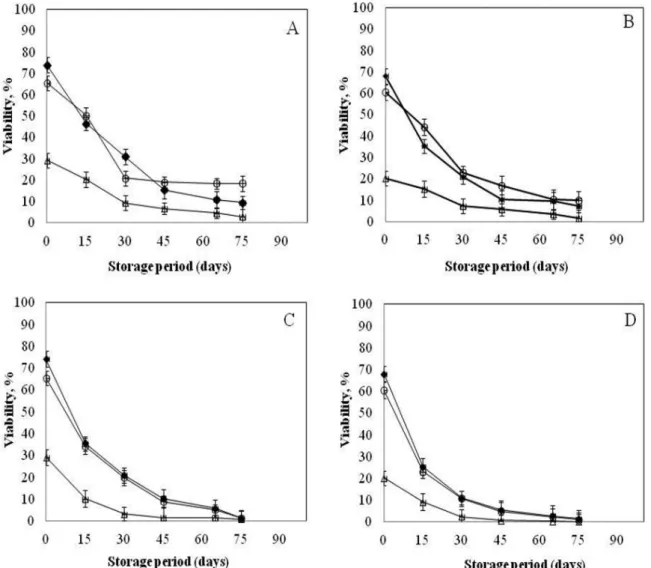
Effectiveness of convective drying to conserve indigenous yeasts with high volatile profile isolated from algerian fermented raw bovine milk (Rayeb)
Texto
Imagem
Documentos relacionados
O único critério localizado que nos permite alguma aproximação é o projeto de Residência em Saúde da Fam1'lia( 2000 ) encaminhado pela Universidade Federal de
Vale dizer, numa variedade de português como a de Porto Alegre, que tende a prevenir a redução vocálica por alçamento, consoantes que não as palatais, dorsais e labiais contribuem
O objetivo do presente estudo será descrever o perfil epidemiológico, do ponto de vista clínico, demográfico e laboratorial, das gestantes encaminhadas ao
Sente-se orgulhoso por fazer parte da empresa porque existe uma boa comunicação entre os colaborado- res?, Pensa que os seus valores são similares aos valores da empresa em
Apresento o meu Relatório de Estágio Profissional enquadrado no 2º ano do 2º Ciclo em Ensino da Educação Física nos Ensinos Básico e Secundário, da
Atividades Reuniões Estratégicas Bloco 1: Análise e discussão; Bloco 2: Metas; Bloco 3: Propostas de ação; Bloco 4: Assembleia Geral; Gincana Família-Escola;
Para todo este processo foi utilizado o software Inkscape, e como já referido todos estes ícones foram exportados em SVG devido a todas as vantagens que este formato tem, como
