Fluorescence in situ hybridization (FISH)
Texto
Imagem

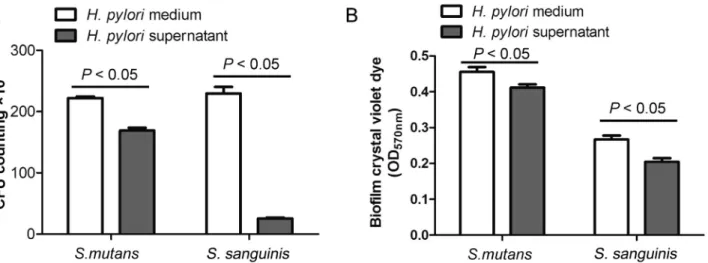
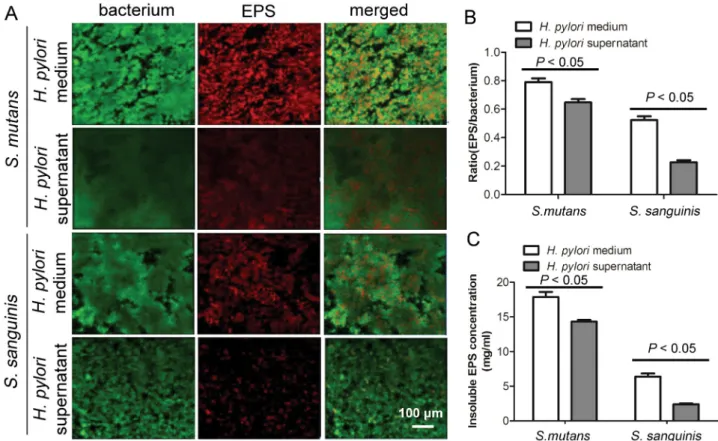
Documentos relacionados
However, gastric metaplasia significantly decreased and complete regression was achieved in 15/28 patients (53.6%) 6 months after eradication of H. pylori , accompanied by
Os requisitos a que uma organização deve obedecer encontram-se descritos na norma através da identificação de diversas etapas baseadas no ciclo de melhoria
Há uma grande preocupação em informar o leitor sobre a vida do escritor, em virtude da suposição de vínculo entre vida e obra, tal como acontecia com a crítica
The degradation rate of the resulting hydrogels was analysed to help choosing the better ratio of the copolymers. 500 μl hydrogels were produced using different combinations of
Outra novidade presente na ISO 17100 é a definição das competências do gestor de projetos de tradução, o qual deve entender como funciona a indústria
Evaluation of Stool Antigen Test, PCR on ORAL Samples and Serology for the Noninvasive Detection of Helicobacter pylori Infection in Children.. Polymerase Chain
O ISD compreende o processo de desenvolvimento humano como algo indissociável dos processos de construção social e cultural do sujeito (BRONCKART, 2007). Sob
RESUMO - Foi desenvolvida uma técnica de criação de ninfas de Deois incompleta ('Walker, 1851) sobre Brachiaria humidicola, em casa telada, em Belém, Part A utilização do método,
