*Correspondence: C. F. Souza. Departmento de Famacologia, Universidade Federal de São Paulo. Rua Botucatu, 862, 04023-062 - São Paulo – SP, Brasil. E-mail: [email protected]
A
vol. 50, n. 4, oct./dec., 2014 http://dx.doi.org/10.1590/S1984-82502014000400011
Effects of long-term fluoxetine treatment on adrenergic plasticity
in rat vas deferens
Cláudio Fontes Souza
1,*, Lúcia Garcez-do-Carmo
1, Isaltino Marcelo Conceição
2,
Thalma Ariani Freitas
2, Roberto Frussa-Filho
11Department of Pharmacology, Federal University of São Paulo, São Paulo, SP, Brazil, 2Laboratory of Pharmacology, Unit of
Mode of Toxin Action, Butantan Institute, São Paulo, SP, Brazil
Chronic antidepressant administration increases neurotrophin levels in the central and peripheral nervous system, leading to an increase of neuronal sprouting, reestablishment of neural networks and neurotransmitter levels. Injured peripheral nerves regenerate at very slow rates. However, the recovery of
the hypogastric nerve in rodents after injury is signiicantly improved with neurotrophin administration. Accordingly, our goal was to determine whether treatment with the antidepressant luoxetine affects
catecholamine levels and neuronal function, after surgical denervation of the rat vas deferens.
Noradrenaline levels in the denervated vas deferens were higher in luoxetine-treated animals than in the
vehicle-treated group, as measured by high performance liquid chromatography. In functional studies of smooth muscle contraction, the responses induced by phenylephrine or ATP, as well as pre-synaptic
α2-adrenoceptor reactivity, were not modiied by chronic treatment with the antidepressant. However, the contraction mediated by neuronal release of noradrenaline induced by tyramine was increased on
days 7 and 21 after denervation in rats treated with luoxetine. These data indicate that luoxetine can
improve functional recovery after rat vas deferens denervation.
Uniterms: Fluoxetine/chronic administration/effects. Denervation. Antidepressants/long term use/
experimental study. Adrenergic neurotransmission. Noradrenaline/neuronal release.
A administração crônica de antidepressivos aumenta os níveis de neurotroinas no sistema nervoso
central, levando a um aumento da arborização neuronal, restabelecendo a rede neural e os níveis de neurotransmissores. Lesões do sistema nervoso periférico mostram uma regeneração muito lenta.
Entretanto, a recuperação após a lesão do nervo hipogástrico em roedores é signiicativamente melhorada após a administração de neurotroinas. Nesse sentido, nosso objetivo foi veriicar se o tratamento com o antidepressivo, luoxetina, interfere nos níveis de catecolaminas e na função neuronal, após a desnervação
cirúrgica do ducto deferente de rato. Nos vasos deferentes desnervados, os níveis de catecolaminas nos
grupos tratados com luoxetina foram maiores que no grupo veículo, quantiicados em cromatograia líquida de alta eiciência (CLAE). Nos estudos funcionais, a contração da musculatura lisa induzida pela fenilefrina ou pelo ATP, assim como a reatividade pré-sináptica α2-adrenérgica, não foram modiicadas
com o tratamento crônico de luoxetina. Contudo, nas contrações mediadas pela liberação neuronal de
norepinefrina induzida por tiramina, observou-se aumento da contração nos dias 7 e 21 após a desnevação
em ratos tratados com luoxetina. Esses dados indicam que a luoxetina pode melhorar a recuperação
funcional do vaso deferente de rato após a desnervação.
Unitermos: Fluoxetina/administração crônica/efeitos. Desnervação. Antidepressivos/uso contínuo/estudo
INTRODUCTION
Although the primary actions of antidepressants are usually related to an increase in monoamine levels in the central nervous system (CNS), the exact molecular mechanisms by which these agents exert their therapeutic effects are not well understood (Slattery, Hudson, Nutt, 2004). Several indications, such as brain imaging of
post mortem animals and of depressed human patients show acorrelation between depression and an axonal atrophy or reduction of the neuronal networks in many CNS areas(Slattery, Hudson, Nutt, 2004; Nestler et al., 2002). These neuronal alterations are reversed by chronic administration of antidepressants, which also induce an increase in axonal sprouting (Carlson et al., 2006;
Santarelli et al., 2003). Neurotrophins such as nerve
grow factor (NGF) and brain derivate neurotrophic factor (BDNF) regulate neuronal development and cell survival in the nervous system (Wang et al., 2008; McAllister,
Katz, Lo, 1999). Chronic antidepressant administration induces an increase in levels of neurotrophic factors (Sofroniew, Howe, Mobley, 2001; Castrén, 2005; Kuipers, Bramham, 2006)and in the expression and activation of BDNF receptors in the hippocampus and frontal cortex of rodents (Russo-Neustadt et al., 2000; Balu et al., 2008).
Chronic luoxetine treatment (selective serotonin reuptake inhibitors) increases NGF levels in cerebral areas, such as the prefrontal cortex and hippocampus (Hassanzadeh, Hassanzadeh, 2009). Due to the fact that antidepressants increase neurotrophin levels in the CNS, it is speculated that this process could also occur in the peripheral nervous system (PNS), where neuronal regeneration after a lesion also depends on neurotrophic factors, mainly NGF. In the urogenital tract, which is rich in neurotrophin receptors (Keast, Kepper, 2001), chronic treatment with exogenous NGF facilitates autonomic nerve regrowth (Burgers et al., 1991)and induces an increase neuronal activity (Milner
et al., 1995). The vas deferens is a urogenital organ with a high neuronal network density, formed in great part by sympathetic nerves (Batra, 1974; Wanigasekara, Kepper, Keast, 2003). Therefore, the aim of the present work was to determine whether chronic administration of luoxetine increases noradrenaline levels in the denervated vas deferens, which could indicate an improvement in the functional recovery process after nerve injury.
MATERIAL AND METHODS
Animals and surgical procedures
Male Wistar rats, 5 months old, weighing between
300-350 g and housed under standard conditions (24 oC and a controlled 12-h light/dark cycle), were used. All experimental procedures were approved by the Ethics Committee of the Federal University of São Paulo.
Surgical denervation of both vasa deferentia was performed according toKasuya and colleagues (1969). Briefly, the rats were anesthetized by intraperitoneal injection of ketamine/xylazine, and through a suprapubic incision, the prostatic portion of the vas deferens was exposed. The serous coat of this prostatic region, where the deferential artery and the hypogastric plexus come in contact with the vas deferens, was carefully picked off from approximately 1 cm from the prostatic end. During this procedure, great care was taken to avoid injury to the vas deferens and the vascular tissue. Later, the vasa deferentia were put back in place and the incision was sutured. Sham-operated rats (controls for the denervated rats) were operated in parallel, under the same conditions as the denervated group, with the exception that no denervation was performed. The rats were euthanized by CO2 asphyxiation 7, 21 and 28 days after the surgery. Both vasa deferentia were removed and prepared for in vitro contraction experiments or biochemical
analysis.
Treatment
The rats were treated daily with fluoxetine (10 mg/kg i.p.) or vehicle (saline with 1% dimethyl sulfoxide) for 11 days prior to surgery and up to 24 h before euthanasia.
Contraction experiments (in vitro)
Tissues preparation
The vasa deferentia from denervated or sham-operated animals, treated with fluoxetine or vehicle, were removed for in vitro contraction experiments.
was washed out and the experimental protocols were started 30 min later.
Contractile response to the indirect adrenergic agonist, tyramine
The vas deferens was contracted by a single concentration of the indirect adrenergic agonist tyramine (100 µM) until the maximal contraction was achieved.
Contractile response to electrical stimulation (ES)
The vas deferens was placed between two parallel platinum electrodes and an electrical ield stimulation was performed with a Grass®S88 stimulator (50 V supramaximal voltage, 3 ms duration, 10 Hz frequency). These electrical stimulation parameters induce only neuronal stimulation without direct muscle stimulation, since this contraction was inhibited by tetrodotoxin, as previously described in our laboratory (Santos et al., 2003). Only the phasic component
of the contraction was considered.
Analysis of pre-synaptic α2-adrenoceptor activity
The vas deferens was electrically stimulated with a frequency of 0.05 Hz (50 V, 3 ms). When the electrical twitch responses were stabilized, concentration-response curves were obtained for the α2-adrenoceptoragonist clonidine. The pIC50 parameter (negative logarithm of the concentration of clonidine that induces a 50% inhibitory effect) was obtained.
Analysis of post-synaptic reactivity
Three cumulative concentration-response curves for the α1-adrenergic agonist phenylephrine were obtained at 30-min intervals. Afterwards, the contraction induced by a single concentration of ATP (10 mM) was recorded. The parameters obtained were maximum effect (Emax, in grams of tension) and pD2 (negative logarithm of the concentration of the agonist that produces 50% of the maximum effect) for phenylephrine.
Biochemical analysis of noradrenaline content
The vasa deferentia from denervated or sham-operated animals treated with fluoxetine or vehicle were removed for biochemical analysis to determine the noradrenaline content of the tissue (Conceição et al., 2005). Briefly, the vasa deferentia were homogenized in 0.4 N perchloric acid, containing 0.1% sodium metabisulite. After centrifugation (7000 x g, 45 min, 4 °C), the supernatant was aspirated and stored at -80 °C until analyzed by high-performance liquid chromatography (HPLC) with electrochemical detection.
Statistical analyses
The results were expressed as the mean ± standard error. The signiicance of differences between two means was evaluated using a two-tailed Student t-test. Treatment groups were compared using one-way analysis of variance (ANOVA) and a post-hoc analysis of differing groups was carried out using a Tukey-Kramer test. P less than 0.05 indicated a statistically significant deference between groups.
Drugs
Chemicals: adenosine 5’-triphosphate (ATP), clonidine hydrochloride, norepinephrine (L-arterenol bitartrate) and tyramine were from Sigma Chemical Co., USA; luoxetine, from Farmasa, BR; phenylephrine (L-phenyl-ephrine hydrochloride) from RBI, USA; and barium chloride, from Merck, BR. Stock solutions of the drugs were prepared in distilled water, with the exception of norepinephrine and phenylephrine, which were dissolved in 0.01 N HCl. Fluoxetine was dissolved in 1% DMSO in deionized water.
RESULTS
Influence of chronic fluoxetine treatment on the denervation and reinnervation profile of rat vas deferens
The characterization of the denervation and reinnervation of rat vas deferens was obtained indirectly, using in vitro contraction experiments, or directly, by
biochemical noradrenaline quantiication.
In vitro contraction experiments
Neuronal viability and pre-synaptic and post-synaptic receptor reactivity were analyzed in sham-operated and denervated vas deferens obtained from rats chronically treated for 11 days prior to surgery, with luoxetine (10 mg/kg/day) or vehicle. The in vitro
contraction experimentswere done on days 7, 21, and 28 after surgery. Neuronal viability was measured under conditions that promote neurotransmitter release, as contraction induced by the indirect adrenergic agonist tyramine or by electrical stimulation.
Tyramine
groups, compared to the respective sham-operated groups, as expected. The lowest contraction was observed on day 7, followed by a gradual and partial recovery of response through day 28. Considering the denervated groups, in the luoxetine-treated denervated group, the amplitude of the contraction was greater than that of its respective vehicle-treated denervated group. Differences in the amplitude of contraction were signiicant on days 7 and day 21. No differences were noted in the amplitude of contraction in either the fluoxetine-treated or vehicle-treated sham-operated groups. These results indicate that luoxetine treatment can improve the contraction induced by tyramine in denervated vas deferens.
Electrically evoked contractions
The amplitude of the contraction induced by electrical field stimulation (10 Hz) in all groups is shown in Figure 1b. As observed with tyramine, in the denervated groups, the amplitude of the contraction was reduced at all time points tested compared to the respective sham-operated groups. The lowest contraction was observed on day 7, followed by a partial recovery of response through day 28. In the fluoxetine-treated group, there was a tendency towards an increase in the amplitude of contraction, but no statistical differences were obtained when compared to the respective vehicle-treated denervated group. In sham-operated groups, no differences were noted between the luoxetine-treated and vehicle-treated groups.
Post-synaptic and pre-synaptic receptor reactivity
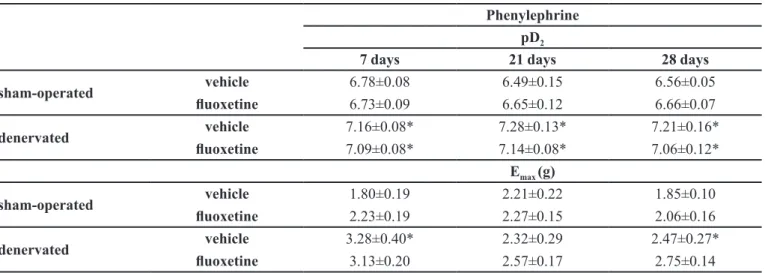
The increase in tyramine-induced contraction observed in the denervated groups after luoxetine treatment could be due to an alteration, induced by fluoxetine, in the synaptic reactivity of the vas deferens. The post-synaptic reactivity of the tissue was studied using exogenous agonists to evaluate the adrenergic and purinergic component of contraction. After denervation, an expected super-sensitivity of the vas deferens to the α1-adrenergic agonist phenylephrine was observed, characterized by an increase in the Emax (in the day 7 and day 28 groups) and by the leftward shift of the concentration-response curves (increase in pD2 value) compared to the respective sham-operated group (Table I). Chronic luoxetine treatment did not induce any alteration in these parameters in either the sham or denervated group, when compared to the respective vehicle-treated group (Table I).
After denervation, an increase in the amplitude of exogenously applied ATP (100 μM) was observed in some groups; however, luoxetine treatment did not induce any differences in the responses when compared
to the respective vehicle-treated group (Table II). The pre-synaptic α2-adrenoceptor reactivity was also analyzed. The α2-adrenoceptor reactivity was evaluated by means of an inhibitory concentration-response curve to the α2-adrenoceptor agonist clonidine over the twitch contractions induced by electrical stimulation (0.05 Hz) in sham-operated vas deferens. No differences were noted in pIC50 value between the vehicle- (9.09±0.10, 8.89±0.07, 8.77±0.18) and luoxetine-treated (8.96±0.07, 8.97±0.03, 8.74±0.04) groups at all time points tested (respectively 7, 21 and 28 days).
Noradrenaline quantification
Noradrenaline levels in rat vas deferens were measured by HPLC with electrochemical detection
FIGURE 1. Histogram showing the maximum effect induced
by (a) a single concentration of tyramine (100µM) and (b) electrical stimulation (3 ms, 50 V, 10 Hz, phasic response) in rat vas deferens. The data are plotted for the sham-operated animals treated with vehicle (white), sham-operated animals treated with luoxetine (dark gray), denervated animals treated with vehicle (light gray) and denervated animals treated with luoxetine (black) for 7, 21 or 28 days after surgery. The bars represent the mean ± S.E.M from four to six experiments. ¤,
indicates a signiicant difference in relation to respective vehicle control group (P<0.05); *, indicates a significant difference in relation to the respective sham-operated group (P<0.05); #, indicates a signiicant difference in relation to the respective day 7 group (P<0.05)
* * # * ¤ * ¤ * # * 0 1 2 3 4
7 days 21 days 28 days
co n tr a c on ( g ) tyramine
Sham -operated / vehicle
Sham -operated / fluoxene
Denervated / vehicle
Denervated / fluoxene (a) 0 1 2 3 4 5 6
7 days 21 days 28 days
co n tr ac on ( g) 10 Hz
Sham -operated / vehicle
Sham -operated / fluoxene
Denervated / vehicle
in sham-operated and denervated groups treated with luoxetine or vehicle. Figure 2 shows that denervation caused a drastic decrease in noradrenaline levels, which was observed in both the treated and untreated groups, reaching the lowest concentration seven days after denervation, followed by a gradual recovery on days 21 and 28. Considering the denervated groups, noradrenaline levels were higher in the fluoxetine-treated group compared to the vehicle-fluoxetine-treated group, at all time points studied. In the sham-operated group, noradrenaline levels were not altered by treatment with luoxetine.
DISCUSSION
The main finding of the present study was that neurotransmitter levels were higher in denervated vas
TABLE I - pD2 and maximum amplitude of contraction (Emax) in grams of tension from the phenylephrine concentration response-curves obtained in vas deferens from sham-operated and denervated rats treated with vehicle or luoxetine for 7, 21 and 28 days. Each value represents the mean ± standard error from 9 to 12 experiments
Phenylephrine
pD2
7 days 21 days 28 days
sham-operated vehicle 6.78±0.08 6.49±0.15 6.56±0.05
luoxetine 6.73±0.09 6.65±0.12 6.66±0.07
denervated vehicle 7.16±0.08* 7.28±0.13* 7.21±0.16*
luoxetine 7.09±0.08* 7.14±0.08* 7.06±0.12*
Emax (g)
sham-operated vehicle 1.80±0.19 2.21±0.22 1.85±0.10
luoxetine 2.23±0.19 2.27±0.15 2.06±0.16
denervated vehicle 3.28±0.40* 2.32±0.29 2.47±0.27*
luoxetine 3.13±0.20 2.57±0.17 2.75±0.14
* indicates a signiicant difference in relation to the sham-operated group (P<0.05).
TABLE II - Maximum amplitude of contraction (Emax), in grams of tension, from a single concentration of ATP (100 µM) obtained in
vas deferens from sham-operated and denervated rats treated with vehicle or luoxetine for 7, 21 and 28 days. Each value represents the mean ± standard error of 5 to 8 experiments
ATP (100 µM)
Emax
7 days 21 days 28 days
sham-operated vehicle 1.08±0.08 1.30±0.21 0.90±0.11
luoxetine 1.35±0.23 1.08±0.12 1.23±0.13
denervated vehicle 1.68±0.16* 1.77±0.26 1.61±0.26*
luoxetine 1.82±0.18 1.64±0.16* 1.54±0.17
* indicates a signiicant difference in relation to the sham-operated group (P<0.05).
deferens from rats chronically treated with fluoxetine compared to denervated vehicle-treated duct. These results indicate that luoxetine treatment could improve the process of reinnervation that normally occurs after vas deferens denervation.
of post-synaptic receptors or changes in the feedback mechanisms mediated by pre-synaptic receptors. However, control experiments with the α1-adrenergic agonist phenylephrine, purinergic agonist ATP and the α2-adrenergic agonist clonidine showed that the reactivity of the receptors associated with these agonists was not modiied by luoxetine treatment. As a result, the increase in tyramine-induced contraction could be due to a greater availability of noradrenaline. This hypothesis was conirmed by results obtained with electrochemical quantification of noradrenaline levels, where higher noradrenaline levels were observed in luoxetine-treated denervated group compared to the vehicle-treated denervated group. In contrast, contractions induced indirectly by electrical stimulation were not signiicantly different, when comparing these two groups, although a tendency towards an increase in the amplitude of contraction was observed. The capacity of fluoxetine to increase neurotransmitter levels or neurotransmitter release in the experiments with tyramine and in the noradrenaline quantification was very slight, at the threshold of statistical validation. Therefore, one could argue that in the electrical stimulation studies, this weak fluoxetine action was not sufficient to induce a
statistically signiicant effect. One possible explanation of this low effect of luoxetine is the treatment protocol (dose and treatment duration). This protocol was based on published data in the literature. In CNS studies, chronic luoxetine treatment (10 mg/kg/day, 21 days) induces an increase in NGF and BDNF levels in several brain regions (Molteni et al., 2006; Hassanzadeh,
Hassanzadeh, 2009). Although the protocol used in our experiments followed one used to obtain effects in the CNS, this protocol may not be adequate for studies in the PNS. Another possibility is that the mechanism involved in the antidepressant-induced increase in neuronal viability is not as eficient in PNS as in CNS.
At the moment, it is not possible to determine the mechanism involved in this increase in noradrenaline levels in fluoxetine-treated denervated vas deferens. Several lines of evidence suggest that in the CNS the antidepressant-induced increase in neuronal viability is mediated by different signaling pathways, speciically through the activation of the transcriptional factor CREB and an increase in the synthesis of neurotrophic factors (Nibuya, Nestler, Duman, 1996; Altar, 1999). In the PNS, the antidepressant effects on neurotrophin levels are not well studied. Because the urogenital tract (and the vas deferens) is rich in NGF and neurotrophin receptors (Keast, Kepper, 2001), the effect of luoxetine treatment could be mediated by the synthesis of NGF. Of note, the luoxetine effects were obtained only in denervated vas deferens. Fluoxetine treatment did not affect sham-operated groups. These data indicate that fluoxetine can only increase noradrenaline levels during neuronal regeneration, thus suggesting that this effect occurs mainly in situations where the process of regeneration after a lesion has already been initiated.
In summary, our results indicate that chronic fluoxetine treatment could change plasticity during peripheral neuron regeneration, increasing catecholamine levels after denervation, where this could highlight a new potential therapeutic beneit of antidepressants.
ACKNOWLEDGMENTS
Thanks go to Teotila Reuter for technical assistance and CNPq for support of this work. Dr. A. Leyva (USA) helped with English editing of the manuscript.
REFERENCES
ALTAR, C.A. Neurotrophins and depression. Trends
Pharmacol. Sci., v.20, n.2, p.59-61, 1999.
FIGURE 2 - Histogram showing the noradrenaline concentration (pmol/mg tissue) in vas deferens from sham-operated animals treated with vehicle (white), sham-operated animals treated with luoxetine (dark gray), denervated animals treated with vehicle (light gray) or denervated animals treated with luoxetine (black) on days 7, 21 and 28 after surgery. The bars represent the mean ± standard error of the mean of 10 experiments. ¤ indicates a signiicant difference in the relation to respective vehicle control group (P<0.05); * indicates a signiicant difference in relation
to the respective sham-operated group (P<0.05); # indicates a signiicant difference in relation to the respective day 7 group (P<0.05)
*
# *
# * ¤
*
¤ # *
¤ # *
0 50 100 150 200 250 300 350
7 days 21 days 28 days
NA (
ρ
mol/mg of
ssue)
Dosage of noradrenaline
Sham-operated / vehicle
Sham-operated / fluoxene
Denervated / vehicle
BALU, D.T.; HOSHAW, B.A.; MALBERG, J.E.; ROSENZWEIG-LIPSON, S.; SCHECHTER, L.E.; LUCKI, I. Differential regulation of central BDNF protein levels by antidepressant and non-antidepressant drug treatments.
Brain Res., v.1211, p.37-43, 2008.
BATRA, S.K. Sperm transport through vas deferens: review of hypotheses and suggestions for a quantitative model. Fertil Steril., v.25, n.2, p.186-202, 1974.
BURGERS, J.K.; NELSON, R.J.; QUINLAN, D.M.; WALSH, P.C. Nerve growth factor, nerve grafts and amniotic membrane grafts restore erectile function in rats. J. Urol.,
v.146, n.2, p.463-68, 1991.
CARLSON, P.J.; SINGH, J.B.; ZARATE, C.A.; DREVETS, W.C.; MANJI, H.K. Neural circuitry and neuroplasticity in mood disorders: insights for novel therapeutic targets.
NeuroRx, v.3, n.1, p.22-41, 2006.
CASTRÉN, E. Is mood chemistry? Nat. Rev. Neurosci., v.6, n.3, p.241-46, 2005.
CONCEIÇÃO, I.M.; JURKIEWICZ, A.; FONSECA, D.R.; OPPERMAN, A.R.; FREITAS, T.A.; LEBRUN, I.; GARCEZ-DO-CARMO, L. Selective release of ATP from sympathetic nerves of rat vas deferens by the toxin TsTX-I from Brazilian scorpion Tityus serrulatus. Br. J. Pharmacol., v.144, n.4, p.519-27, 2005.
HASSANZADEH, P.; HASSANZADEH, A. Effects of psychotropic drugs on nerve growth factor protein levels in the rat brain. Physiol.-Pharmacol., v.13, n.3, p.244-52, 2009.
KASUYA, Y.; GOTO, K.; HASHIMOTO, H.; WATANABE, H.; MUNAKATA, H.; WATANABE, M. Nonspecific denervation supersensitivity in the rat vas deferens “in vitro”. Eur. J. Pharmacol., v.8, n.2, p.177-84, 1969.
KEAST, J.R.; KEPPER, M.E. Differential regulation of trkA and p75 in noradrenergic pelvic autonomic ganglion cells after deafferentation of their cholinergic neighbours. Eur. J. Neurosci., v.13, n.2, p.211-20, 2001.
KUIPERS, S.D.; BRAMHAM, C.R. Brain-derived neurotrophic factor mechanisms and function in adult synaptic plasticity: new insights and implications for therapy. Curr. Opin. Drug Discov. Devel., v.9, n.5, p.580-86, 2006.
MCALLISTER, A.K.; KATZ, L.C.; LO, D.C. Neurotrophins and synaptic plasticity. Annu. Rev. Neurosci., v.22, p.295-318, 1999.
MILNER, P.; CROWE, R.; FERNYHOUGH, P.; DIEMEL,
L.T.; TOMLINSON, D.R.; BURNSTOCK, G. Nerve
growth factor treatment of adult rats selectively enhances innervation of urinogenital tract rather than vascular smooth muscle. Int. J. Dev. Neurosci., v.13, n.5, p.393-01, 1995.
MOLTENI, R.; CALABRESE, F.; BEDOGNI, F.; TONGIORGI, E.; FUMAGALLI, F.; RACAGNI, G.; RIVA, M.A. Chronic treatment with fluoxetine up-regulates cellular BDNF mRNA expression in rat dopaminergic regions. Int. J. Neuropsychopharmacol., v.9, n.3, p.307-17, 2006.
NESTLER, E.J.; BARROT, M.; DILEONE, R.J.; EISCH, A.J.; GOLD, S.J.; MONTEGGIA, L.M.Neurobiology of depression. Neuron, v.34, n.1, p.13-25, 2002.
NIBUYA, M.; NESTLER, E.J.; DUMAN, R.S. Chronic
antidepressant administration increases the expression of cAMP response element binding protein (CREB) in rat hippocampus. J. Neurosci., v.16, n.7, p.2365-72, 1996.
RUSSO-NEUSTADT, A.A.; BEARD, R.C.; HUANG, Y.M.; COTMAN, C.W. Physical activity and antidepressant treatment potentiate the expression of speciic brain-derived neurotrophic factor transcripts in the rat hippocampus.
Neuroscience, v.101, n.2, p.305-12, 2000.
SANTARELLI, L.S.M.; GROSS, C.; SURGET, A.; BATTAGLIA, F.; DULAWA, S.; WEISSTAUB, N.; LEE, J.; DUMAN, R.; ARANCIO, O.; BELZUNG, C.; HEN, R.Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science, v.301, n.5634, p.805-809, 2003.
SANTOS, W.C.; SMAILI, S.S.; JURKIEWICZ, A.; PIÇARRO, I.; GARCEZ-DO-CARMO, L. Dual effect of agmatine in the bisected rat vas deferens. J. Pharm. Pharmacol., v.55, n.3, p.373-80, 2003.
SLATTERY, D.A.; HUDSON, A.L.; NUTT, D.J. Invited review: the evolution of antidepressant mechanisms. Fundam. Clin. Pharmacol., v.18, n.1, p.1-21, 2004.
SOFRONIEW, M.V.; HOWE, C.L.; MOBLEY, W.C. Nerve growth factor signaling, neuroprotection, and neural repair.
WANG, J.W.; DAVID, D.J.; MONCKTON, J.E.; BATTAGLIA, F.; HEN, R. Chronic luoxetine stimulates maturation and synaptic plasticity of adult-born hippocampal granule cells.
J. Neurosci., v.28, n.6, p.1374-84, 2008.
WANIGASEKARA, Y.; KEPPER, M.E.; KEAST, J.R. Immunohistochemical characterisation of pelvic autonomic ganglia in male mice. Cell. Tissue Res., v.311, n.2, p.175-85, 2003.