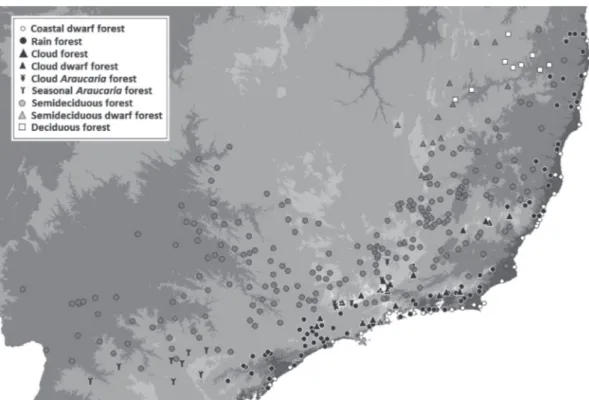
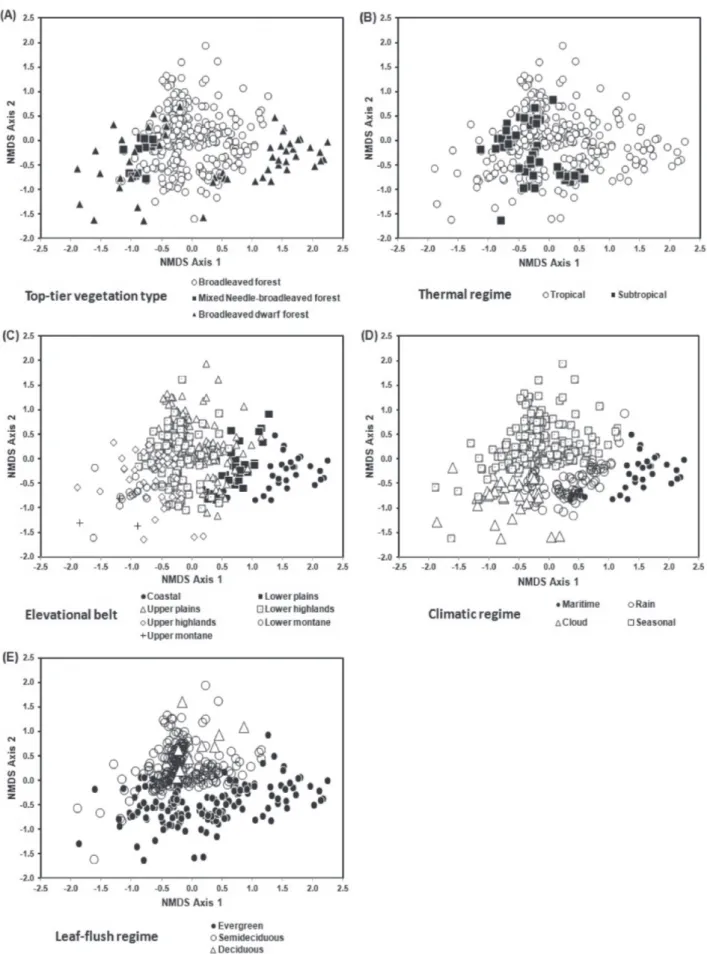
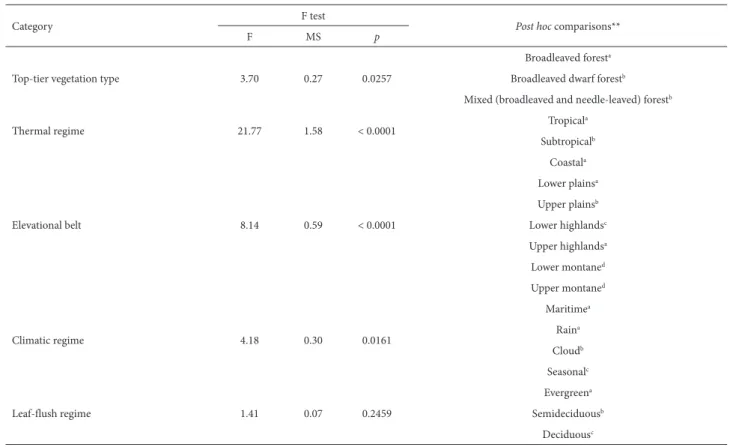
Tree species composition in areas of Atlantic Forest in southeastern Brazil is consistent with a new system for classifying the vegetation of South America
Texto
Imagem
Documentos relacionados
Uma vez criado o banco de dados de peptídeos, precisa-se agora obter os espectros experimentais, conhecidos como tandem mass spectrum - tms, para que a busca
Then the characteristics of cable-stayed bridge, the characteristics of wind field and wind induced vibration of bridges are studied to prepare for the large-span cable-stayed
Este ganho de eficiência é maior quando há pequenas variações na média do processo, já grandes variações na média do processo a eficiência tende a diminuir,
Ao atentar para a quantidade de laboratórios em funcionamento no IFRN Campus Natal Zona Norte, que no total correspondem a 10, e que a responsabilidade
28,29 Here, the spatial patterns show the formation of regions occupied by groups of distinct species, bound by interfaces of empty spaces, whose dynamics is similar to
Os objetivos dos MP explicitam o eixo de contextualização forte, como por exemplo: - preparar quadros para a atuação estratégica na educação; ampliar a capacidade de
A presente Oferta será rescindida em caso de: (i) inadimplência, (ii) descumprimento das obrigações contratuais, (iii) migração para outro plano não participante da oferta,
