Simultaneous allergen inactivation and detoxification of castor bean cake by treatment with calcium compounds
Texto
Imagem
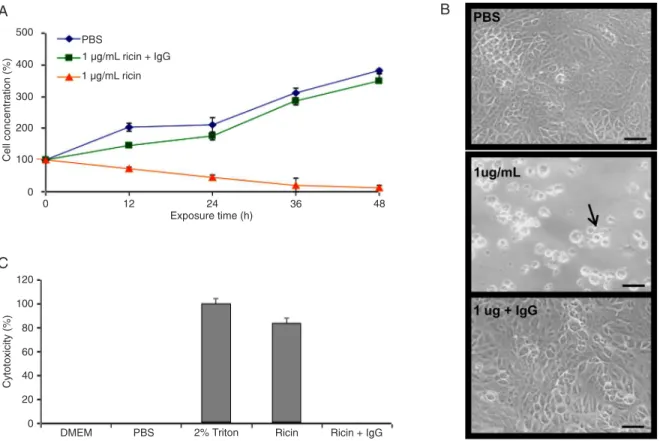
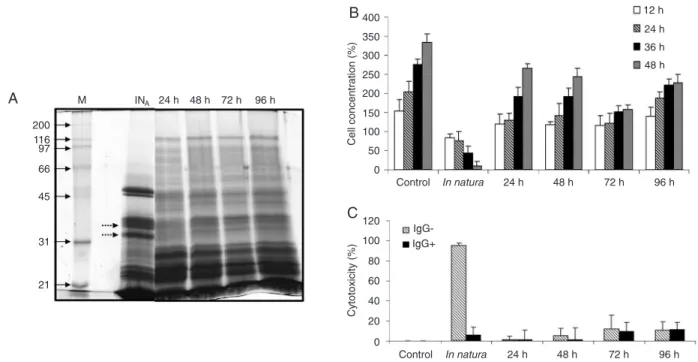
Documentos relacionados
This log must identify the roles of any sub-investigator and the person(s) who will be delegated other study- related tasks; such as CRF/EDC entry. Any changes to
Além disso, o Facebook também disponibiliza várias ferramentas exclusivas como a criação de eventos, de publici- dade, fornece aos seus utilizadores milhares de jogos que podem
Foi neste âmbito que decidi desenvolver uma página de FB para a farmácia onde estagiei, com o objetivo não só de promover os produtos à venda na farmácia, como também
É nesta mudança, abruptamente solicitada e muitas das vezes legislada, que nos vão impondo, neste contexto de sociedades sem emprego; a ordem para a flexibilização como
From the 12 studies included (13 entries) in this meta-analysis, they all started from the same research assumption, in which the elderly with classifications according to the
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Em sua pesquisa sobre a história da imprensa social no Brasil, por exemplo, apesar de deixar claro que “sua investigação está distante de ser um trabalho completo”, ele
Ao longo do monitoramento 643 dias, os coliformes totais apresentaram uma variação, em escala logarítmica, de 6,79 a 9,04 NMP/100g, valores estes elevados, quando comparados com
