1567
RONCUS MELEDAE N. SP. AND NEOBISIUM OCULATUM N. SP., FROM THE
ISLAND OF MLJET, DALMATIA (NEOBISIIDAE, PSEUDOSCORPIONES)
B. P. M. ĆurčIĆ1
, S. E. MAKArOV1, T. rAĐA2, B. S. ILIĆ1, and D. Ž. ANTIĆ1
1 Institute of Zoology, Faculty of Biology, University of Belgrade, 11000 Belgrade, Serbia 2 Speleological Society “Špiljar”, 21000 Split, Croatia
Abstract - Two species of troglobitic false scorpions, Roncus meledae n. sp. and Neobisium oculatum n. sp. (Neobisiidae, Pseudoscorpiones), are described from inside underground habitat, i.e. the Jama Na Žutim Kokom Cave, the Island of Mljet, Dalmatia, Croatia. he analyzed pseudoscorpions prove to difer from all other congeners. It is possible that the subterranean pseudoscorpions analyzed represent relicts of an old north tropical faunal pattern of the Mediterranean.
Key words: Pseudoscorpions, Neobisiidae, new species, Roncus meledae n. sp., Neobisium oculatus n. sp., karst fauna, Dal-matia, Mediterranean.
INTrODuCTION
he Mediterranean is a 3.600 km-long inter-conti-nental sea, from Gibraltar to the coast of Levant, all of it being situated on the same latitudinal belt. Despite this, the oceanographic environment and its biota show a very clear west-to-east gradient which is perhaps better seen as NNW–SSE. As the sea penetrates eastward, it encounters more and more continental and dry climate. he northwest is mountainous and rich in river input and the South-east is dominated by low topography and limited freshwater inlow.
he Adriatic Sea is no part of this west – east gra-dient. Its inclusion in such discussions only confuses. his narrow, shallow sea, extending northwards into a cooler climate, abundantly supplied with nutrients by the river Po, has its own hydrography and biotic history.
Perhaps nowhere else has the history of biogeo-graphic evolution been so closely interwoven with
major environmental changes as in subrecent Medi-terranean. Climate events, sea level changes and, not least, the tectonic conlagrations, have all let their mark, oten recorded in memory and history.
he problems related to the changing sea levels of the Tertiary – the low glacial level of +/-150 m below the present one, have had important biogeographic consequences. he northern half of the Adriatic re-peatedly felt dry. Tectonics and volcanism were also very active during the Tertiary. he coalescence of is-lands, or collapse of island connections, movements of tectonic upliting of new islands and collapse of old ones were associated by destructive tsunamis. he last were probably the reason for several years of climate disruptions.
1568 B. P. M. ĆurčIĆ ET AL.
Palaeoendemic pseudoscorpions are taxa of a tropical stock (Ćurčić, 1988), which were let behind from the Mesozoic or early Cenozoic and survived in isolation ater the ancient distributional continuum was disrupted. It is not easy to analyze the origin and history of the endemic pseudoscorpions of the Di-naric underground habitat because they represent an adaptive and selected fauna. he colonization of the Dinaric subterranean milieu must have begun a long time ago and passed through successive stages during the diferent geological times, including the development of karstic phenomena. herefore, it is probable that the Dinaric area was colonized at the beginning of its existence by false scorpions, which already inhabited Mediterranean forests.
he study of the cave pseudoscorpions inhabiting the Dinaric karst has ofered further proofs of their great age and probably diferent origin. hese species and genera represent the last vestiges of an old fauna that found shelter in the underground domain of the Balkans and elsewhere (Ćurčić, 1986; 1988).
he discovery (1996) of some pseudoscorpions new to science revealed an underground ecosystem, that of the Jama Na Žutim Kokom Cave, Island of Mljet, middle Dalmatia, Croatia. hese species have been described as Roncus meledae n. sp. and Neobisi-um oculatNeobisi-um n. sp. (both taxa inhabit the same cave). Here is an exact study of the newly found species - Roncus meledae n. sp. (Figs. 1 – 8) and Neobisium oculatum n. sp. (Figs. 9 – 24).
Setal designations follow Beier (1963).
SYSTEMATIC PArT
NEOBISIIDAE J. C. CHAMBErLIN, 1930
RONCUS L. KOCH, 1873
RONCUS MELEDAE
B. ĆurčIĆ & rAĐA, NEW SPECIES (Figs. 1-8; Table 1)
Etymology — he new species is named ater its type
locality sensu lato – the Island of Mljet, middle Dal-matia, Croatia.
Material examined — Holotype male from the Jama Na Žutim Kokom Cave, Island of Mljet, middle Dal-matia, Croatia; 26 March 1996, collected by Tonći rađa (together with Neobisium oculatum n. sp.).
Description — he dorsal side of the cephalothorax is with no eyes and, in general, it is longer than wider (Fig. 1, Table 1). he anterior margin of the carapace is wider than the posterior and the carapace resem-bles an irregular quadrate (Fig. 5). he epistome is low and apically rounded (Figs. 4 and 5). he cara-pace bears 27 setae and these lie in four rows. Four setae constitute the anterior row, seven setae belong to the ‘ocular’ series, ten to the median and inter-median rows and six setae constitute the posterior series. No preocular setae are developed in each pre-ocular recess (Fig. 5).
he setal formula of abdominal tergites I – X can be expressed as 7 – 11 – 10 – 11 – 11 – 11 – 11 – 12 – 11 – 9 and is remarkable for the lower number of setae on tergite I. Sternite II of the male has 17 setae along the posterior sternal margin; sternite III car-ries 8 anterior and 12 posterior and 3 suprastigmal microsetae on either side. he fourth sternite has 10 marginal setae and 3 small setae along each of the stigma. Sternites V – X carry 15 – 16 – 14 – 14 – 13 – 12 posterior setae (Fig. 8). he cheliceral spinneret is represented by an extremely low sclerotic knob on the movable inger (Fig. 7). Immediately below, there are teeth of irregular size which diminish both proxi-mally and distally. On the ixed cheliceral inger, the teeth are smaller. Fixed cheliceral inger with six se-tae, movable cheliceral inger with a single seta only (Fig. 7).
1570 B. P. M. ĆurčIĆ ET AL.
1572 B. P. M. ĆurčIĆ ET AL.
Eight trichobothria are carried on the ixed in-ger and four on the movable chelal inin-gers (Figs. 1 and 2). he chelal palm is 2.875 times as long as the chela. he teeth of the ixed inger (57) are triangu-lar, close-set and occupy almost the whole length of the inger; proximal and distal teeth of this inger are smaller and basal teeth merge into dental lamel-la (Fig. 1). Movable chelamel-lal inger with 67 small and close-set teeth (Fig. 1).
he measurements and morphometric ratios of the diferent structures, as well as the tactile seta ra-tios, are presented in Table 1 and in Figs. 1-8. he tibia IV, metatarsus IV and tarsus IV each carry a long tactile seta (Fig. 3, Table 1).
Remarks — he new species is easily distinguished from its congeners, Roncus pripegala and R. insularis, from Dalmatia, Croatia, in the length of the pedipalps and pedipalpal articles, the ratio of length-to-breadth of pedipalpal femur, ratio of tibial length-to-breadth, in the chelal length-to-breadth ratio, length of chelal palm and length of leg IV (Table 1). From R. pripe-gala, R. meledae n. sp. difers in its pedipalpal articles, in the carapacal setation, tergal and sternal setation, number of teeth on ixed and movable chelal inger, as well as in the less elongated pedipalpal podomeres. R. meledae n. sp. difers considerably from R. insu-laris in the tergal and sternal setation, number of chelal teeth, form of the pedipalps, as in the number of morphometric ratios and linear measurements. Furthermore, the new species is clearly distinct from R. insularis in pedipalpal length, pedipalpal femur length-to-breadth ratio, patellar length-to-breadth ratio, chelal breadth ratio, chelal length-to-breadth ratio, total length of leg IV and in the tibia IV length-to-breadth ratio (Table 1).
Morphometric ratios and linear measurements are presented in Table 1.
Distribution — It is probable that the distribution of the new subterranean Roncus species from the is-land of Mljet is relict of the Miocene northern areas of once tropical or subtropical regions. Its present area probably preserves the old biogeographical
dis-tribution and is therefore in line with contemporary records worldwide (Ćurčić, 1972, 1988; Ćurčić et al., 1993, 2004, 2010a, b, c, d; 2011a, b, c, h; Hadži, 1937).
SYSTEMATIC PArT
NEOBISIIDAE J. C. CHAMBErLIN, 1930
NEOBISIUM CHAMBErLIN, 1930
NEOBISIUM OCULATUM
B. ĆurčIĆ & rAĐA, NEW SPECIES (Figs. 9-24; Table 2)
Etymology — Ater the presence of two small eyes on each carapacal side.
Material examined — Holotype male and allotype fe-male from the Jama Na Žutim Kokom Cave, Island of Mljet, middle Dalmatia, Croatia; 26 March 1996, collected by Tonći rađa (together with Roncus mele-dae n. sp.)
Table 1. Linear measurements (in millimeters) and morphometric ratios in Roncusmeledae n. sp., R. pripegala B. Ćurčić, and R. insularis
B. Ćurčić from Croatia. he distinctive traits of Roncus meledae n. sp. are in bold numbers. Abbreviations: M = male, MM = males.
R. meledae n. sp. R. pripegala R. insularis
M M MM
Character Body
Length (1) 4.02 4.46 3.98-4.735
Cephalothorax
Length (2) 0.97 1.03 0.97-1.16
Breadth (2a) 0.815 0.79 0.82-0.94
ratio 2/2a 1.19 1.30 1.18-1.23
Abdomen
Length 3.05 3.43 2.88-3.635
Chelicerae
Length (3) 0.64 0.65 0.66-0.75
Breadth (4) 0.305 0.30 0.30-0.37
Length of movable inger (5) 0.46 0.47 0.44-0.54
ratio 3/5 1.39 1.38 1.36-1.57
ratio 3/4 2.10 2.17 2.015-2.20
Pedipalps
Length with coxa (6) 5.20 6.585 6.645-7.79
ratio 6/1 1.29 2.95 1.56-1.89
Length of coxa 0.71 0.93 0.81-0.95
Length of trochanter 0.63 0.75 0.74-0.88
Length of femur (7) 1.08 1.35 1.39-1.65
Breadth of femur (8) 0.305 0.31 0.31-0.35
ratio 7/8 3.54 4.35 4.46-4.80
ratio 7/2 1.11 1.31 1.34-1.67
Length of patella (tibia) (9) 0.93 1.155 1.23-1.41
Breadth of patella (tibia) (10) 0.41 0.425 0.38-0.445
ratio 9/10 2.27 2.72 3.02-3.33
Length of chela (11) 1.85 2.40 2.475-2.96
Breadth of chela (12) 0.64 0.65 0.57-0.67
ratio 11/12 2.875 3.69 4.24-4.72
Length of chelal palm (13) 0.93 1.10 1.11-1.37
ratio 13/12 1.45 1.69 1.91-2.13
Length of chelal inger (14) 0.92 1.30 1.36-1.59
ratio 14/13 0.99 1.18 1.13-1.39
Leg IV
Total length 3.625 4.17 4.295-4.96
Length of coxa 0.46 0.58 0.56-0.65
Length of trochanter (15) 0.44 0.48 0.46-0.57
Breadth of trochanter (16) 0.18 0.195 0.20-0.25
ratio 15/16 2.44 2.46 1.93-2.95
Length of femur + patella (17) 0.99 1.11 1.19-1.34
Breadth of femur + patella (18) 0.275 0.30 0.28-0.36
ratio 17/18 3.60 3.70 3.64-4.535
Length of tibia (19) 0.94 1.06 1.11-1.32
Breadth of tibia (20) 0.15 0.15 0.15-0.18
ratio 19/20 6.27 7.07 6.53-8.20
Length of metatarsus (21) 0.295 0.35 0.34-0.40
Breadth of metatarsus (22) 0.11 0.13 0.12-0.14
ratio 21/22 2.68 2.69 2.71-3.17
Length of tarsus (23) 0.50 0.59 0.555-0.71
Breadth of tarsus (24) 0.10 0.11 0.10-0.12
ratio 23/24 5.00 5.36 5.17-7.10
TS ratio - tibia IV 0.54 0.56 0.50-0.585
TS ratio - metatarsus IV 0.17 0.16 0.14-0.24
1574 B. P. M. ĆurčIĆ ET AL.
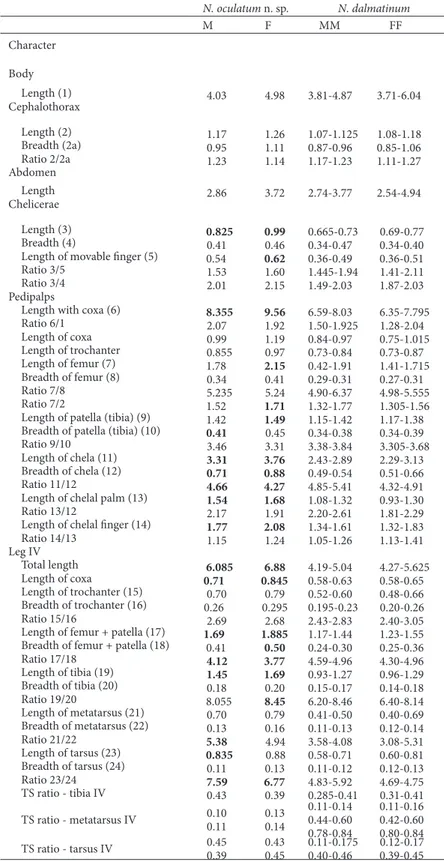
Table 2. Linear measurements (in millimeters) and morphometric ratios in Neobisium oculatum n. sp., and N. dalmatinum B. Ćurčić from Croatia. he distinctive traits of Neobisium oculatum n. sp. are in bold numbers. Abbreviations: M = male, F = female, MM = males, FF = females.
N. oculatum n. sp. N. dalmatinum
M F MM FF Character
Body
Length (1) 4.03 4.98 3.81-4.87 3.71-6.04 Cephalothorax
Length (2) 1.17 1.26 1.07-1.125 1.08-1.18 Breadth (2a) 0.95 1.11 0.87-0.96 0.85-1.06 ratio 2/2a 1.23 1.14 1.17-1.23 1.11-1.27 Abdomen
Length 2.86 3.72 2.74-3.77 2.54-4.94 Chelicerae
Length (3) 0.825 0.99 0.665-0.73 0.69-0.77 Breadth (4) 0.41 0.46 0.34-0.47 0.34-0.40 Length of movable inger (5) 0.54 0.62 0.36-0.49 0.36-0.51 ratio 3/5 1.53 1.60 1.445-1.94 1.41-2.11 ratio 3/4 2.01 2.15 1.49-2.03 1.87-2.03 Pedipalps
Length with coxa (6) 8.355 9.56 6.59-8.03 6.35-7.795 ratio 6/1 2.07 1.92 1.50-1.925 1.28-2.04 Length of coxa 0.99 1.19 0.84-0.97 0.75-1.015 Length of trochanter 0.855 0.97 0.73-0.84 0.73-0.87 Length of femur (7) 1.78 2.15 0.42-1.91 1.41-1.715 Breadth of femur (8) 0.34 0.41 0.29-0.31 0.27-0.31 ratio 7/8 5.235 5.24 4.90-6.37 4.98-5.555 ratio 7/2 1.52 1.71 1.32-1.77 1.305-1.56 Length of patella (tibia) (9) 1.42 1.49 1.15-1.42 1.17-1.38 Breadth of patella (tibia) (10) 0.41 0.45 0.34-0.38 0.34-0.39 ratio 9/10 3.46 3.31 3.38-3.84 3.305-3.68 Length of chela (11) 3.31 3.76 2.43-2.89 2.29-3.13 Breadth of chela (12) 0.71 0.88 0.49-0.54 0.51-0.66 ratio 11/12 4.66 4.27 4.85-5.41 4.32-4.91 Length of chelal palm (13) 1.54 1.68 1.08-1.32 0.93-1.30 ratio 13/12 2.17 1.91 2.20-2.61 1.81-2.29 Length of chelal inger (14) 1.77 2.08 1.34-1.61 1.32-1.83 ratio 14/13 1.15 1.24 1.05-1.26 1.13-1.41 Leg IV
Total length 6.085 6.88 4.19-5.04 4.27-5.625 Length of coxa 0.71 0.845 0.58-0.63 0.58-0.65 Length of trochanter (15) 0.70 0.79 0.52-0.60 0.48-0.66 Breadth of trochanter (16) 0.26 0.295 0.195-0.23 0.20-0.26 ratio 15/16 2.69 2.68 2.43-2.83 2.40-3.05 Length of femur + patella (17) 1.69 1.885 1.17-1.44 1.23-1.55 Breadth of femur + patella (18) 0.41 0.50 0.24-0.30 0.25-0.36 ratio 17/18 4.12 3.77 4.59-4.96 4.30-4.96 Length of tibia (19) 1.45 1.69 0.93-1.27 0.96-1.29 Breadth of tibia (20) 0.18 0.20 0.15-0.17 0.14-0.18 ratio 19/20 8.055 8.45 6.20-8.46 6.40-8.14 Length of metatarsus (21) 0.70 0.79 0.41-0.50 0.40-0.69 Breadth of metatarsus (22) 0.13 0.16 0.11-0.13 0.12-0.14 ratio 21/22 5.38 4.94 3.58-4.08 3.08-5.31 Length of tarsus (23) 0.835 0.88 0.58-0.71 0.60-0.81 Breadth of tarsus (24) 0.11 0.13 0.11-0.12 0.12-0.13 ratio 23/24 7.59 6.77 4.83-5.92 4.69-4.75 TS ratio - tibia IV 0.43 0.39 0.285-0.41 0.31-0.41 TS ratio - metatarsus IV 0.10 0.130.11 0.14
setae along each of the stigma. Sternites V – X with 18 – 12 – 13 – 12 – 13 – 11 setae.
Galea is a slight elevation of the inger margin (Figs. 15 and 22). Fixed cheliceral inger with six, movable cheliceral inger with a single seta. Flagel-lum of eight or nine blades, irst eight of nearly equal size and a most proximal blade smaller than the oth-ers. Other lagellar blades pinnate along their ante-rior margin (Figs. 11 and 19).
Apex of pedipalpal coxa with 4 long setae. Pedi-palpal articles smooth and slender. Fixed chelal in-ger with 101 (male) and 104 (female) teeth, while 81 (male) and 86 (female) teeth are born by the movable chelal inger (Figs. 9 and 17).
Trichobothriotaxy as in Figs. 1 and 9.
Tibia IV with a single, basitarsus IV and telotar-sus IV each with two tactile seta.
Morphometric ratios and linear measurements are presented in Table 2.
Remarks — he species Neobisium oculatum n. sp. difers clearly from its congener, N. dalmatinum, in the presence/absence of eyes, setation of tergites I – X and setation of sternites II – X, in the size of the galea, the form of the pedipalps, chelal dentation and in the disposition of trichobothria on both chelal in-gers. Finally, in the new species the basitarsus carries two tactile setae (three N. dalmatinum) and telotar-sus IV has two such setae (as in N. dalmatinum).
Distribution — Dalmatia, on islands, in caves (Croatia).
Acknowledgments — he inancial help of the Serbian
Min-istry of Education and Science (Grant 173038) is gratefully appreciated.
rEFErENCES
Arvanitidis, C.; Eletheriou, A.; Vanden Berghe, E.; Appeltans, W.; van Avesaath, P.H.; Heip, C.H.R.; and J. Mees (2003). Elec-tronic conference on ‘Marine Biodiversity in the
Mediter-ranean and the Black Sea’: summary of discussions, 7 to 20 April, 2003. MARBENA Proceedings, 3. Flanders Marine Institute (VLIZ): Oostende. IV, 74 pp.
Beier, M. (1963). Ordnung Pseudoscorpionidea (Aterskorpi-one). In : Bestimmungsbücher zur Bodenfauna Europas,
Vol. 1. - Akademie Verlag, Berlin, 1-313.
Ćurčić, B. P. M. (1972). Nouveaux pseudoscorpions cavernicoles de la Serbie et de la Macédoine. Acta Mus. Mac. Sc. Nat. Skopje12, 141-161.
Ćurčić, B. P. M. (1988). Cave-Dwelling Pseudoscorpions of the Di-naric Karst. - Acad. Sci. Art. Slov., Cl. IV, Hist. Nat., Opera 26, Inst. Biol. Ioannis Hadži, 8, Ljubljana, 1-192.
Ćurčić, B. P. M., Lee, V. F., and S. E. Makarov (1993). New and little-known cavernicolous species of Chthoniidae and Neobisiidae (Pseudoscorpiones, Arachnida) from Serbia.
Bijdr. Dierk., 62, 167-178.
Ćurčić, B. P. M., Dimitrijević, R. N., and A. Legakis (2004). he Pseudoscorpions of Serbia, Montenegro, and the repub-lic of Macedonia. Monographs, 8. - Institute of Zoology – Faculty of Biology – university of Belgrade, Hellenic Zoological Society, Committee for Karst and Speleology – Serbian Academy of Sciences and Arts, Institute of Nature Conservation of the republic of Serbia, Belgrade-Athens, 1-400.
Ćurčić, B. P. M., Dimitrijević, R. N., and N. B. Ćurčić (2010a). Ne-obisium deltshevi (Neobisiidae, Pseudoscorpiones), a new endemic cave-dwelling pseudoscorpion from East Serbia.
Arch. Biol. Sci., Belgrade,62 (1), 191-198.
Ćurčić, B. P. M., Dimitrijević, R. N., Rađa, T., Ćurčić, N. B., and
M. Milinčić (2010b). Chthonius (Chthonius) onaei n. sp. (Chthoniidae, Pseudoscorpiones), a new epigean species from Croatia. Arch. Biol. Sci., Belgrade,62 (2), 494-499.
Ćurčić, B. P. M.,Rađa, T., Ćurčić, S. B. and N. B. Ćurčić (2010c). On Roncus almissae n. sp., R. krupanjensis n. sp., and R. radji n. sp., three new pseudoscorpions (Pseudoscorpi-ones, Neobisiidae) from Croatia and Serbia, respectively.
Arch. Biol. Sci., Belgrade,62 (2), 503-513.
Ćurčić, B. P. M., Makarov, S. E., Rađa, T., Ćurčić, S. B., Ćurčić, N. B., and M. Pecelj (2010d) On three new cave pseudoscor-pions (Pseudoscorpiones, Neobisiidae) from Mt. Mosor, Dalmatia (Croatia). Arch. Biol. Sci., Belgrade,62 (3), 813-828.
Ćurčić, B. P. M., Dimitrijević, R. N., Makarov, S. E., Milinčić, M., Pecelj, M., and T. Rađa (2011a). Two new pseudoscorpions from the uN Administered Province of Kosovo and Croa-tia. Arch. Biol. Sci., Belgrade, 63(1), 235-244.
Ćurčić, B. P. M., Ćurčić, S. B., Ćurčić, N. B., and B. S. Ilić (2011b).
1576 B. P. M. ĆurčIĆ ET AL.
false scorpion from Montenegro. Arch. Biol. Sci., Belgrade,
63(1), 245-250.
Ćurčić, B. P. M., Rađa, T., Makarov, S. E., Ćurčić, S. B., Ilić, B. S., and R. N. Dimitrijević (2011c). A cavernicolous pseu-doscorpion of the genus Chthonius (Chthonius) C. L. Koch from Dalmatia. Arch. Biol. Sci., Belgrade, 63(2), 493-498.
Ćurčić, B. P. M., Rađa, T., Dimitrijević, R. N., Makarov, S. E., and
M. Milinčić (2011h). A new cave pseudoscorpion from Serbia (Pseudoscorpiones, Chthoniidae). Arch. Biol. Sci.,
Belgrade, 63(4), 1257-1263.