w ww.e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Anti-angiogenic
effects
of
ethanolic
extract
of
Artemisia
sieberi
compared
to
its
active
substance,
artemisinin
Zohreh
Abdolmaleki
a,
Hossein-Ali
Arab
a,∗,
Saeid
Amanpour
b,
Samad
Muhammadnejad
baDepartmentofPharmacology,FacultyofVeterinaryMedicine,UniversityofTehran,Tehran,Iran bTumorModelResearchCenter,CancerInstituteofIran,TehranUniversityofMedicalSciences,Tehran,Iran
a
r
t
i
c
l
e
i
n
f
o
Articlehistory: Received8August2015 Accepted3November2015 Availableonline10February2016
Keywords: Angiogenesis
Humanumbilicalendothelialcells Chickchorioallantoicmembrane Artemisiasieberi
Artemisinin
a
b
s
t
r
a
c
t
Angiogenesisplaysakeyroleintumorgrowth,invasionandmetastasisofcancerdiseasesand there-fore,theinhibitionofangiogenesiscanprovideanimportanttherapeuticapproachincancerdiseases. Thisstudywasdesignedtocomparetheanti-angiogenicactivitiesoftheethanolicextractofArtemisia sieberiBesser,Asteraceae, anditsactivesubstance,artemisinininbothinvitroandinvivomodels. Tocomparecytotoxicity levelofethanolic extractof A.sieberi withartemisinin, different concen-trations(1–100g/ml)weretestedusingMTTassayonhumanumbilicalveinendothelialcells.The anti-angiogenicpropertiesofserialconcentrationsofethanolicextractofA. sieberiandartemisinin wereexaminedonhumanumbilicalveinendothelialcellsusingathree-dimensionalangiogenesisassay (invitromodel)andinthechickchorioallantoicmembraneassayasinvivomodel.Theeffectsofethanolic extractofA.sieberiandartemisininwerealsotestedontheexpressionofVEGFR-1,VEGFR-2andCD34 genesusingreal-timePCR.EthanolicextractofA.sieberiandartemisininsignificantly(p<0.001)inhibited theangiogenesisinthehumanumbilicalveinendothelialcellsculturewhilsttheethanolicextractofA. sieberishowedhighereffectinaconcentration-dependentfashion(p<0.001).Thechickchorioallantoic membraneangiogenesiswasalsocompletelyinhibitedbyethanolicextractofA.sieberiat concentra-tionof33ng/100l/egg.ThegeneexpressionanalysisshowedthattheethanolicextractofA.sieberi andartemisininreducedthetranscriptionofVEGFR-1,VEGFR-2andCD34genesina concentration-dependentmanner.ThisstudydemonstratedthattheethanolicextractofA.sieberiisstronglyableto inhibittheangiogenesisinhumanumbilicalveinendothelialcellsandchickchorioallantoicmembrane modelscomparedtotheartemisinin.
©2016SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Allrightsreserved.
Introduction
Angiogenesis, the process of new blood vessel formation frompre-existingvasculatures,isvitalprocessintheembryonic development,female reproduction cycleand woundhealing. It alsoplaysacrucial rolein thepathogenesisofvarious diseases including rheumatoid arthritis, diabetic retinopathy, psoriasis, juvenilehemangiomaandtumorgrowthandmetastasis(Folkman, 1995;Koch,1998;FerraraandAlitalo,1999).Incancerdisease,this processisaninevitableprocessforthedevelopmentandgrowthof solidtumorsbeyond2–3mm3.Formationofthenewbloodvessels
isrequiredtosustainthedisseminationoftumorcells;otherwiseit mayresultinaninactivatedanddormanttumordisease(Caoetal., 2011).Manyproandanti-angiogenicfactorsareknowntocontrol theangiogenesisprocess.Themembersofthevascularendothelial
∗ Correspondingauthor. E-mail:[email protected](HA.Arab).
growthfactor(VEGF)andfibroblastgrowthfactorgenefamiliesare recognizedaspro-angiogenesisagents(Caoetal.,2008;O’Reilly etal.,1997),whiletransforminggrowthfactor-(TGF-)and endo-statinareintroduced asanti-angiogenesis compounds(Roberts, 2008;Nybergetal.,2005).Thebalancebetweenthesepro-and anti-angiogenicagentsmodulatesthenewblood vasculaturegrowth innormalcondition(Ferrara,2010;Danieleetal.,2012)andso,it canbespeculatedthatthedisruptionofthisbalanceisrequiredto maintaintheprogressoftumors.Severalanti-angiogenic agents havebeendevelopedtoinhibitdifferentstagesofangiogenesisin tumorgrowthprocesses.Theseagentsmostlyeffectivein combi-nationchemotherapy,havebecomeanattractiveapproachtotreat somecancerdiseases(EllisandHicklin,2008;Schmidt,2009).
The use of herbal drugs as combination therapy has been suggestedbydifferentresearcherstoinhibittheangiogenesisin patientswith solid tumors (Borchers et al., 1997; Saiki, 2000). Among these, attention toward the Artemisia-derived products includingartemisinin (ART) hasbeenincreased in recent years (Sagar et al.,2006).Thereisevidence thatthe crudeextract of
http://dx.doi.org/10.1016/j.bjp.2015.11.008
theArtemisiaannua(qinghao)hasbeenusedasantipyretic, astrin-gent,sedativeandanti-malarialagentsfrommorethan2000years agoinChina(Zhu,1987;Meshnicketal.,1989).Artemisinin(ART) as a natural product derived from plant A. annua (qinghao) is nowusedasworldwidecombinationtherapyagainst multidrug-resistantspecies ofplasmodium (Klayman, 1985; Millerand Su, 2011).Artemisininisasesquiterpenelactonecontainingaperoxide bridge.Thisperoxidebridgeisbelievedtoberesponsiblefor differ-entpharmacologicaleffectsofART(PosnerandO’Neill,2004).The sesquiterpenelactones(SL)aretheactiveconstituentsofavariety ofmedicinalplantsincludingArtemisiasieberiBesser,Asteraceae. ManystudieshaveshownthatSLareabletoinhibitangiogenesis, leadingtotheanti-tumoractivityofthesecompounds(Jeongetal., 2002;Okaetal.,2007;Hayashietal.,2009;Pratheeshkumarand Kuttan,2011;Yueetal.,2013;Tsuboietal.,2014).
Recently,artemisininhasbeenalsofoundtoactasapotent anti-tumoragentbothinvitroandinvivo(LiandHickman,2011).ART asacytotoxicagentisknowntobeabletoinhibitthegrowthof manycancercelllines(Efferthetal.,2001,2003;Efferth,2006). IthasbeenalsodemonstratedthatARTanditsbioactive deriva-tivesareabletoinhibittheangiogenesisactivityandmetastasis ofsomecancercelllines(LiandHickman, 2011).Further stud-ieshavereportedthatlowconcentrationoftwoARTderivatives, artesunateanddihydroartemisinin,inhibitedangiogenesisinchick chorioallantoicmembrane (CAM)and reducedthelevelsoftwo majorVEGFreceptorsonhumanumbilicalveinendothelialcells (HUVEC)(Chenetal.,2003).Itisalsoshownthatartesunatewas abletoinhibitproliferationanddifferentiationofhuman microvas-culardermal endothelialin a concentration-dependent manner (Huan-huanetal.,2004).Thepossibilityofangiogenesisinhibition byARTderivativesinsolidtumorscanbeapromisingtherapeutic approachforpreventionoftumordissemination.Artemisiasieberi isatypicaldesertplantthatgrowsinIran,Palestine,Syria,Iraq, Turkey,AfghanistanandCentralAsia(Podlech,1986)anditsART contentwasdeterminedforthefirsttime,byArabetal.in2006. TheyfoundthatthelevelofartemisininintheA.sieberi(0.14–0.2% ofdriedweightatdifferentseasons)iscomparabletothatofthe otherspeciesincludingA.annua(Arabetal.,2006).Infurtherstudy theyreportedthatboth theplantextract anda granule formu-lationderived fromtheplantextract posepotentanti-coccidial effectinbroilerchickens(Arabetal.,2009;Kaboutarietal.,2014). Thepresentstudywasdesignedtoinvestigatetheanti-angiogenic effectsofethanolicextractofA.sieberi(EEA)incomparisontoits activesubstance,artemisinininHUVECculture(invitromodel)and inchickCAMasinvivomodel.Itisalsoaimedtocomparetheeffects ofEEAandARTonthegeneexpressionofVEGFR-1,VEGFR-2and CD34recognizedasanimportantmolecularmarkerfor angiogen-esis.
Materialsandmethods
Samplepreparation
Theaerialparts ofArtemisiasieberi,Besser,Asteraceae,were collectedinSeptember2012,fromTaftCounty,YazdProvincein central partof Iran. Theplant wasidentified by Prof. Valiallah Mozaffarian. Voucherspecimens weredeposited and identified at theCentral Herbarium of Faculty of Science,Tehran Univer-sity,Tehran,Iran(No.TUH-24118).Thedriedandgroundmaterial (100g)wassubjectedtoextractionwith500mlofa96%ethanol atroomtemperatureinthreecyclesof72heach.Aftereachcycle, theextractwasfilteredthroughfilterpaperandthesolventwas removedbyvacuumdistillationunder reducedpressure witha rotaryevaporator(38◦C).Theyieldofcrudeextractsfromethanol was7.32gand keptin−20◦C untilfurtheruse.To preparethe
stocksolutions,theextractand activesubstanceweredissolved in dimethylsulfoxide (DMSO). The mixtures werethen filtered andsterilizedusing0.22mfilter,andkeptfrozenuntil analyz-ing.SerialdilutionsoftheEEAandARTwerefreshlypreparedfrom stocksolutionbeforeusebydissolvingthemintothecellculture media.
DeterminationofartemisinininA.sieberi
The A. sieberi extract was dissolved in ethanol and then, hydrolyzedbyadditionof0.2%NaOH.Thehydrolyzedsolutionwas incubatedat50◦Cfor30minandthenneutralizedwith0.2mol/l aceticacid.Thepreparedsolutionwasfilteredandstoredat−20◦C untilHPLCanalysis.TheHPLCsystemconsistedofa10cm×4mm nucleosilC18column,Waters510HPLCpumpandWaters490E UV detector. The mobile phase was prepared from 20mmol/l phosphate buffer(K2HPO4+KH2PO4) and methanol (60:40, pH
7.9)withruntimeof1.5ml/min.Thevolumeofsampleinjected intothechromatographwas20latflowrateof1.5ml/min.An artemisininstandard (SigmaAldrichChemical Co.)wasusedas externalstandardtoplotthecalibrationcurveoftheartemisinin detectedfromtheextract.
Cellcultureandviabilityassay
TheHUVECwasobtainedfromNationalCellBank ofPasteur Institute.CellswereculturedinM199mediasupplementedwith 10%heatinactivatedfetalbovineserum(FBS)and100IU/ml peni-cillin plus100g/ml streptomycin(Gibco,USA). Thecells were incubatedinahumidifiedatmosphereof5%CO2at37◦C.Thecell
viabilitytestwasconductedwiththe 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide(MTT)assay.TheHUVECwere seededatadensityof1×104 cellsperwellintoa96-wellplate
tocomparecytotoxicityof EEAwithARTonnormalcells. After 24hincubation,cellswereexposedtoagradedconcentrationof 1,3.3,10,33and100g/mlofEEAandARTandtheincubationwas furthercontinuedby48h.ThemaximumconcentrationofDMSO addedtothecellsculturemediawas0.1%,andthisconcentrationof thesolventwasalwaysusedascontrol.Subsequently,MTTreagent (0.5mg/mlinsterilePBS)wasaddeddirectlytothewells.Cells werereturnedtotheincubatorfor4h.Theformationof insolu-blepurpleformazanfromyellowishMTTbyenzymaticreduction wasdissolvedinDMSOafterremovalofsupernatant.The viabil-ityofcellswasdeterminedbymeasuringtheabsorbancevalues at540nmusinganELISAreader(BioTek,USA,Gen5powerwave xs2).
Invitroanti-angiogenicassay
Table1
Nucleotidesequencesoftheprimersusedforreal-timeRT-PCR.
Gene Forwardprimer(5′–3′) Reverseprimer(5′–3′) Size
Beta2M ATGCCTGCCGTGTGAAC ATCTTCAAACCTCCATGATG 91
VEGFR-1 CATACTCAACTCCTGCCTTCTC CGCCCTGGAGTCATCAAAC 185
VEGFR-2 GTATGGAGGAGGAGGAAGTATG CCGTCTGGTTGTCATCTGG 173
CD34 ACCCCAGAGTTACCTACCCAG TGTCGTTTCTGTGATGTTTGTTG 152
Invivoanti-angiogenicassay
Thechickchorioallantoicmembrane(CAM)modelusedto com-paretheinvivoanti-angiogenicactivityofEEAandARTwasbased ontheproceduredescribedbyKirchneretal.withsome modifi-cations(Kirchneretal.,1996).Briefly,fertilizedchickeneggswere incubatedat37◦Cwith55–60%humidity.Onday3ofincubation, asquareholewasmadeontheoutershelland0.5–1mlalbumin wasremovedbyan18-gaugehypodermicneedletoallow detach-mentoftheCAMdevelopmentandthen,theholewascarefully sealed.Theeggswerereturnedtotheincubatorandthe incuba-tionwascontinuedfor5 days.On day9of incubation,a blank filterdiskanddiscscontainingdifferentconcentrationsofEEAand ART(1–100ng/100l/egg)andtheirsolvent(asnegativecontrol) wereplacedonthetopofCAMundersterilecondition.The num-bers of newly formed blood vessels in CAM of each treatment groupwerecountedonanopticalmicroscope(Olympus,Tokyo, Japan),andtheneovascularzonesofCAMunderthediskswere photographed.
Geneexpressionanalysis
HUVECcellswere exposedtoserialconcentrations of1, 3.3, 10,33 and 100g/ml of EEA andART for 24h, RNAextraction wasdonefrom106 treatedcells usinghighpureRNA isolation
kit(Roche,USA,11828665001)basedontheprotocolmanual.To removegenomiccontamination,mRNAwastreatedwithDNaseI usingakit(Fermentas,Lithuania)basedontheprotocoldescribed bythemanufacturer.ConcentrationsofRNAweredeterminedby a UV spectrophotometer (Eppendorf, Germany) and cDNA syn-thesiswasperformedusing a Revert Aid TMfirststrand cDNA synthesiskit(Fermentas,Lithuania).ForPCRquantitiesanalysis, thedesiredprimersofVEGFR-1,VEGFR-2,CD34andBeta2M(as internal control)genes were designed using Allele ID software (Table1).Inthereactiontube,10lSYBRGreenmastermix(Takara, Japan)wasaddedto2lcDNAsamples,0.5lforward,10pmol reverseprimersand7lnuclease-freewater(Qiagen,Hilden, Ger-many)toaccomplishPCRin20lofreactionmixtureusingreal timePCRinstrument(Qiagen65HO,USA).Theprogramfor reac-tionwasadenaturedat95foroneminutefollowedby40cycles ofinitiation at95 for 10s, annealing at56◦C for 15s, elongat-ingat 72◦C for 20sand a singlefinal stepat 58◦C for 90s. At theendoftheprogram,themeltingcurvewascheckedandthe datawereanalyzedbyCTcalculationusingRESTsoftware(Qiagen, USA).
Statisticalanalysis
Allvalueswerepresentedasmeans±SEMandevaluatedfor sta-tisticalsignificancewithone-wayANOVAfollowedbyBonferroni’s posthoctest.Anon-linearregressionanalysisbyGraphPadprism software6.0wasusedtoobtainGI50(theconcentrationcaused50% growthinhibitionofculturedcells).Apvalueslessthan0.05was consideredsignificant.
Results
Artemisinincontentoftheextract
TheretentiontimeforartemisinininthepreparedHPLCsystem withflowrateof1.5ml/minwas5.4min.Acalibrationcurvewas constructedusingalinearregressionalgorithmmethodby plot-tingtheareaundercurve(AUC)versusdifferentconcentrationsof standardsolutions.Thiscalibrationcurvewasobtainedby injec-tionofdifferentconcentrationsof1,5,20,50 and100g/mlof artemisinin.TheartemisinincontentoftheEEAwas0.18%ofdried weight(DW)oftheA.sieberi.
EffectofEEAandARTontheviabilityofHUVEC
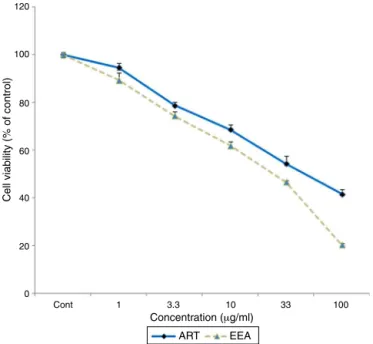
TheeffectsofdifferentconcentrationsofA.extractsonHUVEC culturedcellsisshowninFig.1.Asthisfigureshows,theEEAand ARTwerenotabletoreducethenumberofHUVECupto concentra-tionof3.3g/ml.However,theexposureoftheHUVECwithEEA andARTatconcentrationsof10–100g/mlwasassociatedwith significantreductioninthenumberofviablecells(p<0.05).
Anti-angiogeniceffectsofEEAandARTonHUVEC
After3–5daysincubationoftheHUVEC,theuntreatedcontrol wellsshowedbranchingpatternoftube-likecapillaries.However,
120
100
80
60
40
20
0
Cont 1 3.3
Concentration (μg/ml)
Cell viability (% of control)
ART EEA
10 33 100
120
100
80
60
40
20
∗ ∗∗
∗∗ ∗∗ ∗∗ ∗∗ ∗∗
∗∗
∗∗
0
Cont 0.001 0.033 0.05 Concentration (μg/ml)
% of g
ro
wth inhibition
EEA ART
1 3.3 10
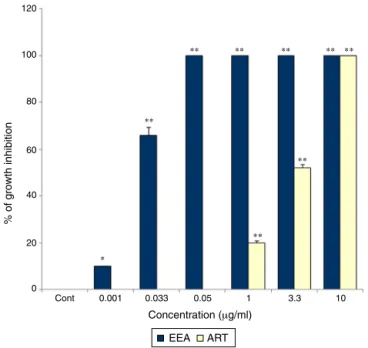
Fig.2.Theinhibitoryeffectsofdifferentconcentrationsof0.001,0.033,0.05,1,3.3, 10,33and100g/mlARTandEEAonHUVECcapillarytubeformationina three-dimensionalcollagenmatrix.Theassaywasconductedondextran-coated cytodex-3microcarriersandtheendothelialcellattachedtoparticleshasbeenmigrated throughthecollagenmatrix.Valuesareexpressedasmean±SEMfromatleastthree independentexperiments(*p<0.05and**p<0.01comparedtothecontrol).
these tube-like vessels formation by HUVEC were significantly reducedinthewellsexposedtoEEAandARTina concentration-dependentmannerwithdifferentpotencies(Fig.2).Apictureof theinhibitoryeffectsinducedby1g/mlEEAandARTon tube-likecapillariesformationinHUVECcultureisillustratedinFig.3B and C. It was foundthat the endothelialcells attached to par-ticles hadbeen proliferatedand migrated throughthecollagen matrixincontrolwellsofcellcultureplates(Fig.3A).However, capillarytubeformationwasstronglysuppressedinwellstreated with0.05 to100g/ml EEA and 10 to 100g/ml ART. Among these,theconcentrationof0.05g/mlofEEAshowedthe high-estinhibitoryeffects(p<0.001)onthree-dimensionalcultureof HUVEC(Fig.2).TreatmentofculturecellswithARTatthe con-centrationsof 0.001–0.05g/ml hadnosignificanteffectonthe proliferation of HUVEC. However,1g/ml or more of ART sig-nificantlyinhibitedcellproliferation(p<0.001).The50%growth inhibitoryeffects(GI50)ofbothcompoundsestimatedon
capillar-iesformationinHUVECisshowninTable2.Asthistableshowsthe GI50intheEEAissignificantlylessthanART,illustratingthehigher
anti-angiogenicactivityofEEAthanART.
Table2
Theconcentrationcaused50%growthinhibition(GI50)byethanolicandextractof A.sieberi(EEA)andARTonHUVECculture.TheGI50levelsareestimatedbya non-linearregressionanalysisusingtheGraphPadPrismsoftware(R2>0.97).
Sample GI50(g/ml)
ART 3.5
EEA 0.024
120
100
80
∗ ∗
∗ ∗∗
∗∗ ∗∗ ∗∗
60
40
20
0
Cont 1 3.3 10
Concentration (μg/ml)
% of g
ro
wth inhibition
ART EEA
20 33 50
Fig. 4. Effects of ART and EEA on chick chorioallantoic membrane (CAM) angiogenesis. The disks containing different concentrations of ART and EEA (1–100ng/100l/egg)wereplacedonselectedareasofCAMonday9andtheeggs werefurtherincubatedat37◦Cfor48h.Onecontrolandonedrug-containingdisk
wereplacedoneachCAMandthenthenumbersofmicrovesselsunderthedisks werecountedusingalightmicroscope.*p<0.05,**p<0.01comparedtocontrol, n=3.
Anti-angiogeniceffectsofEEAandARTonCAM
TheresultsoftheCAMassayshowedthatEEAandARTwere significantly(p<0.01)abletoimpedetheinvivoangiogenesisina concentration-dependentmanner.Fig.4showstheanti-angiogenic effectsofEEAandARTonCAMofthechickenembryos.The anti-angiogenicactivityofEEAandARTstartedatlowconcentrationsof 1ng/100l/eggwith10and5%ofinhibition,respectively(Fig.4). Thecompleteanti-angiogenicactivityofEEAwasshownat con-centrationof 33,while ARTatthesameconcentrationhad75% ofinhibition(Fig.5BandCrespectively).ARTatconcentrationof 50ng/100l/eggcompletelyinhibitedangiogenesis(Fig.4). How-ever,exposureofCAMwithsolventsoftestedcompoundsusedas
Fig.5. RepresentativepicturesillustratetheinhibitoryeffectsofARTandEEAonchickembryochorioallantoicmembrane(CAM).Thepicturesshowtheinhibitionof angiogenesisinthepresenceof33ng/100l/eggconcentrationofEEAandARTinwhich:(A)Control,(B)EEAwith100%inhibitionand(C)ARTwith75%inhibition.
negativecontrol,didnotshowanyanti-angiogenicactivityonthe CAM(Fig.5A).
EffectsofEEAandARTongeneexpression
Dataanalysisofthereal-timeRT-PCRresultsshowedadecrease in transcript levels of VEGFR-1, VEGFR-2 and CD34 in HUVEC cultureexposedtotheEEAinaconcentration-dependentmanner. AsshowninFig.6A,theconcentrationsof3.3,10and 33g/ml EEAdiminishedtranscriptionofVEGFR-1to85.9%(p<0.01),71.6% (p<0.001)and48.9%(p<0.001).TheARTatthesameconcentration wasalsoable todecrease themRNAexpression ofVEGFR-2 to 84.1%(p<0.01),71%(p<0.001)and41.8%(p<0.001),respectively. Moreover,the mentioned concentrations of EEA reduced tran-scriptionofCD34geneto89.1%(p<0.01),81%(p<0.01)and72.1% (p<0.001)respectively(Fig.6A).ARTalsoinhibitedtheexpression of VEGFR-1, VEGFR-2 and CD34 in a concentration-dependent manner. Fig. 6B demonstrates that ART at the concentrations of3.3,10 and33g/mldiminishedtranscription ofVEGFR-1to respectivevalues of 88.8% (p<0.01), 71.6% (p<0.01) and 65.8% (p<0.001). The transcription of VEGFR-2 was also reduced by ARTtothe levelsof 84.8% (p<0.01), 77.3% (p<0.01) and 65.3% (p<0.001), respectively, whereas the expression of CD34 was ablatedto89.9%(p<0.01),84.1%(p<0.01)and77.1% (p<0.001), respectively.
Discussion
Itwaspreviouslysuggestedthattheanti-malarialartemisinin derivativesmayalsopossessantitumoractivity(Efferthetal.,2001, 2003;Efferth,2006).Somestudieshaveshowntheanti-angiogenic potentialofseveralartemisininderivativesinvitrousingthe cul-turedHUVECmodel(Chenetal.,2003,2004a,b).In thepresent investigation,we sought toinvestigate the invitro and in vivo antiangiogenicactivityof ethanolicextract ofA. sieberiin com-parisontoartemisinininthree-dimensionalcultureofHUVECand CAMofchicks,respectively.BothEEAandARTwereabletoreduce theangiogenesisactivitiesinHUVECcultureina concentration-dependentmanner.However,theEEAshowedgreaterinhibitory effectsonangiogenesisasitwasdeterminedinthelevelofGI50.
Supportingtheinvitrofindings,itwasfoundthat EEAandART wereabletoinhibitangiogenesisinCAMofthechickswithhigher potencyforEEA.ThecytotoxicityassayshowedthatEEAandART arerelativelysafeonHUVECattheconcentrationsinhibitingthe angiogenesis.
Artemisininanditsderivativemoleculescontainan endoper-oxidebondreactingwithaferrousironatomleadingtocytotoxic carbon-centered radicals (Olliaro et al., 2001). These carbon-centered radicals are potent alkylating agents involving in the
1.2
∗
∗∗
∗∗ ∗∗
∗∗
∗∗∗∗∗∗
∗∗∗
∗∗∗
∗∗∗
∗∗∗ ∗∗∗ ∗∗
∗
∗
∗∗
∗∗∗∗ ∗∗
∗∗
∗∗ ∗∗∗
∗∗∗ ∗∗ ∗
∗∗∗ ∗∗∗ ∗∗
∗
A
B
F
old change
Change f
old
1.0
0.8
0.6
0.4
0.2
0.0
1.2
1.0
0.8
0.6
0.4
0.2
0.0
Cont 1 3.3
EEA concentration (μg/ml)
ART concentration (μg/ml)
VEGFR-1 VEGFR-2 CD34
10 33 100
Cont 1 3.3 10 33 100
anti-malarial activity of these compounds. There is evidence that the anti-tumor activity of artemisinin is attributed tothe endoperoxidebondpresentonthestructureofmolecules(Galal et al., 2002). Besides a broad spectrum of activity against malaria,studieshaveidentifiedpotentialanti-cancermechanisms ofartemisinin derivatives(ART) suchasnormalizationof upre-gulated Wnt/-catenin pathway in colorectal cancer (Li et al., 2007). Other anti-cancer activity pathways that ART may be involvedincludeinhibition ofenhancedangiogenesisassociated withtumors(Wartenbergetal.,2003;Dell’Evaetal.,2004;Chen etal.,2004a,b;Anfossoetal.,2006;LiandZhou,2005;Longoetal., 2006;Wuetal.,2006;Zhouetal.,2007).Theantitumoractivities ofARThavebeenreportedbysomeinvestigatorsbybothinvitro andinvivomodelsofstudies(Caoetal.,2009;Chenand Cleck, 2009).Efferthetal.havereportedthatartesunateasawater sol-ublederivativeofartemisininwaspotentiallyeffectiveagainsta varietyofcancercelllinesincludingleukemiaand colontumor cells.TheaveragelevelofGI50estimatedagainstthesecelllines
were1.11±0.56Mand2.13±0.74M,respectively.However, thenon-smalllungcancercelllinesareshownahighlevelofGI50
(25.62±14.95M).AnintermediateGI50valueswasobtainedfor
melanomas,breast,ovarian,prostate,CNS,and renalcancercell lines(2001).
A potent anti-angiogenic activity against tumor stroma cells in the rat embryos are shown by artemisinin and its derivatives (Firestone and Sundar, 2009). Oh et al. reported that different thioacetal ART derivatives including 10-a-phenylthiodihydroartemisinins and particularly 10-b-benzenesulfonyl-9-epi-dihydroartemisinin had inhibitory activities against HUVEC proliferation on matrigel. They also showed that both compounds had strong inhibitory effect on angiogenesis in CAM at the concentration of 5g/egg by 90% (2004). Furthermore, Chen et al. showed that artesunate and dihydroartemisinin(DHA)significantlyinhibitedangiogenesisin aconcentration-dependentmannerwithdifferentconcentration of 12.5–50M and 2.5–50M, respectively (2003).Supporting these,Huan-huanetal.foundthatartesunatehighlyinhibitedcell proliferationanddifferentiationofhumanmicrovasculardermal endothelialcells in a concentration-dependent mannerranging from 12.5 to 100M (Huan-huan et al., 2004). The antitumor activitiesof ART have beenalso reportedin in vivo modelsby few studies. It is found that DHA significantly inhibited chick CAMangiogenesisatlowconcentrationsof5–30nmol/100L/egg (Chen et al., 2004a). Jung et al. reported that anti-angiogenic activity of a non acetal-type derivative of artemisinin showed onCAMmodelismoreorcomparabletothoseoffumagillinand thalidomide with complete inhibitory effects at concentration 80nmol/egg(Jungetal.,2006).
Inthesafetyassay,wefoundthatEEAandARTdidnotshow anytoxicityontheHUVECatlowconcentrationswhichthese com-poundswereextensivelyabletoinhibittheangiogenesis.Though EEAandARTreducedtheviabilityofHUVECatconcentrationof 10–100g/ml.Acompleteanti-angiogenesiswasshownbyEEA at concentrationof 0.05g/ml onthree-dimensional culture of HUVEC.Dataobtainedfrominvitroassayshowedthata partial inhibition was started by EEA at concentration of 0.001g/ml, andat0.05–100g/mlitshowedafullanti-angiogenesiseffects withoutanysubstantialtoxic effectonthecells.In thepresent study,wealsofoundthatextractwasabletoshowanti-angiogenic effectsonchickCAMmodelinaconcentration-dependent man-ner.Astheresultsshowedtheinhibitoryeffects ofEEA started at very low concentrationof 1ng/100l/egg and completed at 33ng/100l/egg. These results showed higher anti-angiogenic activityof EEAthanART.It seemsthebioactivityofA.sieberiis notexclusivelyduetoartemisininandothercomponentsofthe Artemisiasp.OurdataareinagreementwiththeresultsofEfferth
etal.showingthatvariousA.annuaextractshave aremarkable heterogeneityofinhibitoryactivity,which reflectingthe biolog-icalvariabilitybetweendifferentplantindividuals(Efferthetal., 2011).
VEGF anditstyrosine kinasereceptors,VEGFR-1 and mainly VEGFR-2(KDR),areknownasmainmediatorsofangiogenesisin both physiological and pathological conditions. VEGF induces a cascadeof signalingpathwaysthroughbindingtothereceptors resultinginproliferation,migration,survivalandvascular perme-ability(KarkkainenandPetrova,2000;Kerbel,2008).VEGFandits receptorsareknownasthemostcommonlytargetedmolecules foranti-angiogenicdrugsindifferenttypesoftumorgrowth pro-cesses(Borgstrometal.,1996;Dvorak,2002;Yla-Herttualaetal., 2007).ARThave beenshown toinhibitproliferation, migration andtubeformationofHUVECthroughinhibitingtheVEGFbinding tosurfacereceptorsonHUVECandreducingexpressionofVEGF receptorsFlt-1andKDR/flk-1onHUVEC(Chenetal.,2004a,b;Cao etal.,2009).ArtemisininreducestheexpressionoftheVEGF recep-torKDR/flk-1invarioustypesoftumorsandendothelialcellsand slow thegrowthof humanovarian cancerHO-8910 xenografts innudemice(Chenet al.,2004b;LiandZhou,2005;Wuetal., 2006; Zhouet al.,2007;Cao etal., 2009).Induction of apopto-sisbyartesunateinHUVECisassociatedwithdownregulationof anti-apoptoticproteinBcl-2andupregulationofpro-apoptotic pro-tein BAX(Wu etal.,2004).Inaddition,mRNAexpressionof30 outof90ofangiogenesis-relatedgenesissignificantlycorrelated withthecellularresponsetoART.Thiscansupportthe hypothe-sisthatARTexerttheiranti-tumoractivitiesthroughinhibitionof tumorangiogenesis(Anfossoetal.,2006).Therefore,inthepresent studythereal-timeRT-PCRtestwasusedtoevaluatetheEEAand ARTeffectsonVEGFR-1,VEGFR-2andCD34geneexpression lev-els.DataobtainedfromthisstudydemonstratedthatEEAandART significantlyreducedVEGFR-1,VEGFR-2andCD34transcriptgene expression.
Thesynergisticeffectsofthemixturesofbioactiveconstituents andtheirbyproductscontainedintheplantextractsmayaccount fortheapparentenhancedpotencyoftheplantextractscompared to the individual constituents (Wagner and Ulrich-Merzenich, 2009; Ulrich-Merzenich et al., 2009). The extract of Artemisia speciescontainmixturesoforganicchemicalsthatcomefrom dif-ferent part of plant includingleaves, stems,flowers and roots. ThemajorcompoundsisolatedfromArtemisiaspeciesinclude ter-penoids, flavonoids, coumarins,acetylenes, caffeoylquinic acids, andsterols(Tanetal.,1998).ThereasonwhyART,asoneofthe mainconstituentoftheArtemisiaspeciesdidnotshowgreatfold changeontheexpressionofVEGFR-1,VEGFR-2andCD34rather thantotheEEAisnotclearbythistime.Butgenerally,wecanstate thatthetotalextractoftheA.sieberionthegeneexpressionwas greaterthanthatoftheindividuallycorrespondingactive ingre-dient.Thesefindingconfirmedtheanti-angiogenesisactivitiesof thetestcompoundsdemonstratedinvitroandinvivomodelsof ourstudy.Itsuggeststhatthepossiblemechanismforinhibitionof tube-likevesselsformationbyHUVECmaybethrough downreg-ulationofVEGFR-1,VEGFR-2andCD34genesinresponsetothe extracts.
Authors’contribution
ZA(PhDstudent)contributedbyrunningthelaboratorywork, biologicalstudies,analysisofthedataanddraftingthepaper.SM contributedtoeditingofthemanuscriptandconductingthe labo-ratorywork.SAcontributedtorunningthelaboratoryworksand preparingmaterialforanalysis.HAAdesignedthestudy,supervised thelaboratorywork,wasresponsibleforfinancialsupport,analysis ofthedata,andcontributedtocriticalreadingofthemanuscript. Alltheauthorshavereadthefinalmanuscriptandapprovedits submission.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
References
Anfosso,L.,Efferth,T.,Albini,A.,Pfeffer,U.,2006.Microarrayexpressionprofilesof angiogenesis-relatedgenespredicttumorcellresponsetoartemisinins. Phar-macogenomics6,269–278.
Arab,H.A.,Rahbari,S.,Rassouli,A.,Moslemi,M.H.,Khosravirad,F.,2006. Determi-nationofartemisinininArtemisiasieberiandanticoccidialeffectsoftheplant extractinbroilerchickens.Trop.Anim.HealthProd.38,497–503.
Arab,H.A.,Mardjanmehr,S.H.,Shahbazfar,A.,Rassouli,A.,Abdollahi,M.,Nekouie, O.,2009.Toxicopathologiceffectsofartemisinininbroilerchickensfollowinga singleoraldose:anLD50study.Int.J.PoultrySci.8,808–812.
Borchers,A.T.,Hackman,R.M.,Keen,C.L.,Stern,J.S.,Gershwin,M.E.,1997. Comple-mentarymedicine:areviewofimmunomodulatoryeffectsofChineseherbal medicines.Am.J.Clin.Nutr.66,1303–1312.
Borgstrom,P.,Hillan,K.J.,Sriramarao,P.,Ferrara,N.,1996.Completeinhibitionof angiogenesisandgrowthofmicrotumorsbyanti-vascularendothelialgrowth factorneutralizingantibody:novelconceptsofangiostatictherapyfrom intrav-italvideomicroscopy.CancerRes.56,4032–4039.
Cao,Y.,Arbiser,J.,D’Amato,R.J.,D’Amore,P.A.,Ingber,D.E.,Kerbel,R.,Klagsbrun,M., Lim,S.,Moses,M.A.,Zetter,B.,Dvorak,H.,Langer,R.,2011.Forty-yearjourney ofangiogenesistranslationalresearch.Sci.Transl.Med.3,114rv3.
Cao,Y.,Zhong,W.,Sun,Y.,2009.Improvementofantiangiogeniccancertherapyby understandingthemechanismsofangiogenicfactorinterplayanddrug resis-tance.Semin.CancerBiol.19,338–343.
Cao,Y.,Cao,R.,Hedlund,E.M.,2008.Regulationoftumorangiogenesisand metas-tasisbyFGFandPDGFsignalingpathways.J.Mol.Med.(Berl.)86,785–789. Chen,H.H.,Zhou,H.J.,Fang,X.,2003.Inhibitionofhumancancercelllinegrowthand
humanumbilicalveinendothelialcellangiogenesisbyartemisininderivatives invitro.Pharmacol.Res.48,231–236.
Chen,H.H.,Zhou,H.J.,Wang,W.Q.,Wu,G.D.,2004a.Antimalarialdihydroartemisinin alsoinhibitsangiogenesis.CancerChemother.Pharmacol.53,423–432. Chen,H.H.,Zhou,H.J.,Wu,G.D.,Lou,X.E.,2004b.Inhibitoryeffectsofartesunateon
angiogenesisandonexpressionsofvascularendothelialgrowthfactorandVEGF receptorKDR/flk-1.Pharmacology71,1–9.
Chen,H.X.,Cleck,J.N.,2009.AdverseeffectsofanticanceragentsthattargettheVEGF pathway.Nat.Rev.Clin.Oncol.6,465–477.
Daniele,G.,Corral,J.,Molife,L.R.,deBono,J.S.,2012.FGFreceptorinhibitors:rolein cancertherapy.Curr.Oncol.Rep.14,111–119.
Dell’Eva,R.,Pfeffer,U.,Vene,R.,Anfosso,L.,Forlani,A.,Albini,A.,Efferth,T.,2004. InhibitionofangiogenesisinvivoandgrowthofKaposi’ssarcomaxenograft tumorsbytheanti-malarialartesunate.Biochem.Pharmacol.68,2359–2366. Dvorak,H.F.,2002.Vascularpermeabilityfactor/vascularendothelialgrowthfactor:
acriticalcytokineintumorangiogenesisandapotentialtargetfordiagnosisand therapy.J.Clin.Oncol.20,4368–4380.
Efferth,T.,Dunstan,H.,Sauerbrey,A.,Miyachi,H.,Chitambar,C.R.,2001.The anti-malarialartesunateisalsoactiveagainstcancer.Int.J.Oncol.18,767–773. Efferth,T.,Sauerbrey,A.,Olbrich,A.,Gebhart,E.,Rauch,P.,Weber,H.O.,Hengstler,
J.G.,Halatsch,M.E.,Volm,M.,Tew,K.D.,Ross,D.D.,Funk,J.O.,2003.Molecular modesofactionofartesunateintumorcelllines.Mol.Pharmacol.64,382–394. Efferth,T.,2006.Molecularpharmacologyandpharmacogenomicsofartemisinin
anditsderivativesincancercells.Curr.DrugTargets7,407–421.
Efferth,T.,Herrmann,F.,Tahrani,A.,Wink,M.,2011.Cytotoxicactivityofsecondary metabolitesderivedfromArtemisiaannuaL.towardscancercellsincomparison toitsdesignatedactiveconstituentartemisinin.Phytomedicine18,959–969. Ellis,L.M.,Hicklin,D.J.,2008.VEGF-targetedtherapy:mechanismsofanti-tumour
activity.Nat.Rev.Cancer8,579–591.
Ferrara,N.,Alitalo,K.,1999.Clinicalapplicationsofangiogenicgrowthfactorsand theirinhibitors.Nat.Med.5,1359–1364.
Ferrara,N., 2010.Pathways mediatingVEGF-independenttumorangiogenesis. CytokineGrowthFactorRev.21,21–26.
Firestone,G.L.,Sundar,S.N.,2009.Anticanceractivitiesofartemisininandits bioac-tivederivatives.Expert.Rev.Mol.Med.11,e32.
Folkman,J.,1995.Angiogenesisincancervascular,rheumatoidandotherdisease. Nat.Med.1,27–31.
Galal,A.M.,Ross,S.A.,ElSohly,M.A.,ElSohly,H.N.,El-Feraly,F.S., Ahmed,M.S., McPhail,A.T.,2002.Deoxyartemisininderivativesfromphotooxygenationof anhydrodeoxydihydroartemisininandtheircytotoxicevaluation.J.Nat.Prod. 65,184–188.
Griffith,C.K.,Miller,C.,Sainson,R.C.,Calvert,J.W.,Jeon,N.L.,Hughes,C.C.,George, S.C.,2005.Diffusionlimitsofaninvitrothickprevascularizedtissue.TissueEng. 11,257–266.
Hayashi, A., Arai, M., Fujita, M., Kobayashi, M., 2009. Pyripyropenes, fungal sesquiterpenesconjugatedwithalpha-pyroneandpyridinemoietiesexhibits anti-angiogenicactivityagainsthumanumbilicalveinendothelialcells.Biol. Pharm.Bull.32,1261–1265.
Huan-huan,C.,Li-Li,Y.,Shang-Bin,L.,2004.Artesunatereduceschicken chorioallan-toicmembraneneovascularisationandexhibitsantiangiogenicandapoptotic activityonhumanmicrovasculardermalendothelialcell.CancerLett.211, 163–173.
Jeong,S.J.,Itokawa,T.,Shibuya,M.,Kuwano,M.,Ono,M.,Higuchi,R.,Miyamoto, T.,2002.CostunolideasesquiterpenelactonefromSaussurealappa,inhibitsthe VEGFRKDR/Flk-1signalingpathway.CancerLett.187,129–133.
Jung, M., Tak, J., Chung, W.Y., Park, K.K., 2006. Antiangiogenic activity of deoxoartemisininderivativesonchorioallantoicmembrane.Bioorg.Med.Chem. Lett.16,1227–1230.
Kaboutari,J.,Arab,H.A.,Ebrahimi,K.,Rahbari,S.,2014.Prophylacticand thera-peuticeffectsofanovelgranulatedformulationofArtemisiaextractonbroiler coccidiosis.Trop.Anim.HealthProd.46,43–48.
Karkkainen,M.J.,Petrova,T.V.,2000.Vascularendothelialgrowthfactor recep-torsintheregulationofangiogenesisandlymphangiogenesis.Oncogene19, 5598–5605.
Kerbel,R.S.,2008.Tumorangiogenesis.N.Engl.J.Med.358,2039–2049.
Kirchner,L.M.,Schmidt,S.P.,Gruber,B.S.,1996.Quantitationofangiogenesisinthe chickchorioallantoicmembranemodelusingfractalanalysis.Microvasc.Res. 51,2–14.
Klayman,D.L.,1985.Qinghaosu(artemisinin):anantimalarialdrugfromChina. Science228,1049–1055.
Koch,A.E., 1998.Angiogenesis:implicationsfor rheumatoidarthritis. Arthritis Rheum.41,951–962(Review).
Li,Q.,Hickman, M.,2011.Toxicokineticandtoxicodynamic(TK/TD)evaluation to determine and predict the neurotoxicity of artemisinins.Toxicol 279, 1–9.
Li,L.N.,Zhang,H.D.,Yuan,S.J.,Tian,Z.Y.,Wang,L.,Sun,Z.X.,2007.Artesunate attenuatesthegrowthofhumancolorectalcarcinomaandinhibitshyperactive Wnt/beta-cateninpathway.Int.J.Cancer121,1360–1365.
Li, J.,Zhou,H.J.,2005. Dihydroartemisinininhibitsthe expressionofvascular endothelialgrowthfactorinK562cells.YaoXueXueBao40,1041–1045. Longo,M.,Zanoncelli,S.,Manera,D.,Brughera,M.,Colombo,P.,Lansen,J.,Mazué,
G., Gomes,M., Taylor, W.R.,Olliaro, P., 2006. Effects of the antimalarial drugdihydroartemisinin(DHA)onratembryosinvitro.Reprod.Toxicol.21, 83–93.
Meshnick,S.R.,Tsang,T.W.,Lin,F.B.,Pan,H.Z.,Chang,C.N.,Kuypers,F.,Chiu,D.,Lubin, B.,1989.Activatedoxygenmediatestheantimalarialactivityofqinghaosu.Prog. Clin.Biol.Res.313,95–104.
Miller,L.H.,Su,X.,2011.Artemisinin:discoveryfromtheChineseherbalgarden.Cell 146,855–858.
Nehls,V.,Drenckhahn,D.,1995.Anovelmicrocarrier-basedinvitroassayforrapid andreliablequantificationofthree-dimensionalcellmigrationand angiogene-sis.Microvasc.Res.50,311–322.
Nyberg,P.,Xie,L.,Kalluri,R.,2005.Endogenousinhibitorsofangiogenesis.Cancer Res.65,3967–3979.
Oh,S.,Jeong,I.H.,Ahn,C.M.,Shin,W.S.,Lee,S.,2004.Synthesisand antiangio-genicactivityofthioacetalartemisininderivatives.Bioorg.Med.Chem.12, 3783–3790.
Oka,D.,Nishimura,K.,Shiba,M.,Nakai,Y.,Arai,Y.,Nakayama,M.,Takayama,H., Inoue,H.,Okuyama,A.,Nonomura,N.,2007.Sesquiterpenelactone partheno-lidesuppressestumorgrowthinaxenograftmodelofrenalcellcarcinomaby inhibitingtheactivationofNF-B.Int.J.Cancer120,2576–2581.
Olliaro,P.L.,Haynes,R.K.,Meunier,B.,Yuthavong,Y.,2001.Possiblemodesofaction oftheartemisinin-typecompounds.TrendsParasitol.17,122–126.
O’Reilly,M.S.,Boehm,T.,Shing,Y.,Fukai,N.,Vasios,G.,Lane,W.S.,Flynn,E., Birk-head,J.R.,Olsen,B.R.,Folkman,J.,1997.Endostatin:anendogenousinhibitorof angiogenesisandtumorgrowth.Cell88,277–285.
Podlech,D.,1986.Artemisia.In:Rechinger,K.H.(Ed.),FloraIranica,Florades Iranis-chenHochlandesundderumrahmendenGebirge,Graz,Akademische Druck-undVerlagsanstaltAustria,vol.158.,pp.159–223.
Posner,G.W.,O’Neill,P.M.,2004.Knowledgeoftheproposedchemical mecha-nismofactionandcytochromep450metabolismofantimalarialtrioxaneslike artemisininallowsrationaldesignofnewantimalarialperoxides.Acc.Chem. Res.37,397–404.
Pratheeshkumar,P.,Kuttan,G.,2011.Vernolide-Ainhibitstumourspecific angio-genesisbyregulatingproinflammatorycytokinesVEGF,MMPsandTIMP.Eur.J. Pharmacol.10,10–18.
Roberts,D.D.,2008.Thrombospondins:fromstructuretotherapeutics.Cell.Mol.Life Sci.65,669–671.
Schmidt,C.,2009.Whydotumorsbecomeresistanttoantiangiogenesisdrugs?J. Natl.CancerInst.101,1530–1532.
Saiki,I.,2000.AKampomedicine“Juzen-taiho-to”–preventionofmalignant pro-gressionandmetastasisoftumorcellsandthemechanismofaction.Biol.Pharm. Bull.23,677–688.
Tan,R.X.,Zheng,W.F.,Tang,H.Q.,1998.Biologicallyactivesubstancesfromthegenus Artemisia.PlantaMed.64,295–302.
Tsuboi,K.,Matsuo,Y.,Shamoto,T.,Shibata,T.,Koide,S.,Morimoto,M.,Guha,S.,Sung, B.,Aggarwal,B.B.,Takahashi,H.,Takeyama,H.,2014.Zerumboneinhibitstumor angiogenesisviaNF-Bingastriccancer.Oncol.Rep.31,57–64.
Ulrich-Merzenich,G.,Panek,D.,Zeitler,H.,Wagner,H.,Vetter,H.,2009.New per-spectivesforsynergyresearchwith‘omic’technologies.Phytomedicine16, 495–508.
Wagner,H.,Ulrich-Merzenich,G.,2009.Synergyresearch:approachinganew gen-erationofphytopharmaceuticals.Phytomedicine16,97–110.
Wartenberg,M.,Wolf,S.,Budde,P.,Grünheck,F.,Acker,H.,Hescheler,J., Warten-berg,G.,Sauer,H.,2003.Theantimalariaagentartemisininexertsantiangiogenic effectsinmouseembryonicstemcell-derivedembryoidbodies.Lab.Invest.83, 1647–1655.
Wu,G.D.,Zhou,H.J.,Wu,X.H.,2004.Apoptosisofhumanumbilicalveinendothelial cellsinducedbyartesunate.Vascul.Pharmacol.41,205–212.
Wu, X.H., Zhou, H.J., Lee, J., 2006. Dihydroartemisinin inhibits angiogenesis inducedbymultiplemyelomaRPMI8226cellsunderhypoxicconditionsvia downregulationofvascularendothelialgrowth factorexpressionand sup-pressionofvascularendothelialgrowthfactorsecretion.AnticancerDrugs17, 839–848.
Yla-Herttuala,S.,Rissanen,T.T.,Vajanto,I.,Hartikainen,J.,2007.Vascularendothelial growthfactors:biologyandcurrentstatusofclinicalapplicationsin cardiovas-cularmedicine.J.Am.Coll.Cardiol.49,1015–2106.
Yue,G.G.,Chan,B.C.,Kwok,H.F.,Wong,Y.L.,Leung,H.W.,Ji,C.J.,Fung,K.P.,Leung,P.C., Tan,N.H.,Lau,C.B.,2013.Anti-angiogenesisandimmunomodulatoryactivitiesof ananti-tumorsesquiterpenebigelovinisolatedfromInulahelianthus-aquatica. Eur.J.Med.Chem.59,243–252.
Zhou,H.J.,Wang,W.Q.,Wu,G.D.,Lee,J.,Li,A.,2007.Artesunateinhibitsangiogenesis anddownregulatesvascularendothelialgrowthfactorexpressioninchronic myeloidleukemiaK562cells.Vascul.Pharmacol.47,131–138.