O R I G I N A L R E S E A R C H P A P E R
Colitis generates remote antinociception in rats: the role of the
L
-arginine/NO/cGMP/PKG/K
ATPpathway and involvement
of cannabinoid and opioid systems
Andre´ Luiz dos Reis Barbosa• Rhamon Barroso de Sousa•Joa˜o Nathanael Lima Torres • Thiago Mattar Cunha• Fernando de Queiroz Cunha•Pedro Marcos Gomes Soares•
Ronaldo de Albuquerque Ribeiro• Mariana Lima Vale•Marcellus Henrique Loiola Ponte Souza
Received: 12 August 2014 / Revised: 17 September 2014 / Accepted: 19 September 2014 / Published online: 7 October 2014
ÓSpringer Basel 2014
Abstract
Objective and design The aim of this study was to investigate the possible involvement of the NO/cGMP/ PKG/KATP
?
pathway, cannabinoids and opioids in remote antinociception associated with 2,4,6-trinitrobenzene sulph onic acid (TNBS)-induced colitis.
Methods TNBS-induced colitis was induced by intraco-lonic administration of 20 mg of TNBS in 50 % ethanol. After induction, carrageenan (500lg/paw) or prostaglan-din (PG) E2(100 ng/paw) was injected in the rat’s plantar
surface and hypersensitivity was evaluated by the elec-tronic von Frey test. Rats were pre-treated with L-Noarg
one hour before carrageenan injection. L-Arginine was
given 10 min before L-Noarg injections. ODQ, KT 5823,
glibenclamide (Glib), naloxone and AM 251 or AM 630 were administered 30 min prior to carrageenan or PGE2
treatments.
Results Colitis induction by TNBS reduced PGE2 or
carrageenan-induced hypersensitivity. Antinociception produced by TNBS-induced colitis was reversed signifi-cantly (P\0.05) by L-Noarg, ODQ, KT 5823,
glibenclamide, naloxone, AM251 and AM630 treatments. Conclusions TNBS-induced colitis causes antinocicep-tion in the rat paw. This disorder appears to be mediated by activation of the NO/cGMP/PKG/KATP pathway,
endo-cannabinoids and endogenous opioids. This information may contribute to a better understanding of peripheral neurological dysfunctions occurring in Crohn’s disease. Keywords TNBS-induced colitis Hypersensitivity
NO OpioidsCannabinoids
Introduction
The term inflammatory bowel disease (IBD) is used to describe a group of chronic, recurrent intestinal disorders of complex pathogenesis, which are represented mainly by Crohn’s disease (CD) and ulcerative colitis (UC). The ex-traintestinal manifestations of IBD are diverse. The exact incidence of neurological complications is unknown, with reports varying from 0.2 to 35.7 % [1–6].
The most common extra-intestinal dysfunction found in patients or in animals with CD is peripheral neuronal desensitisation. In patients with CD, colonic hypersensi-tivity has been demonstrated with a decrease in somatic sensitivity [7]. In animal studies, it has also been demon-strated that acute stimuli such as visceral distension or
Responsible Editor: Ji Zhang.
A. L. R. Barbosa (&)
LAFFEX Laboratory of Experimental Physiopharmacology, School of Physiotherapy, Biotechnology and Biodiversity Center Research (BIOTEC), Federal University of Piauı´-CMRV, Parnaı´ba, PI 64202-020, Brazil
e-mail: [email protected]
A. L. R. BarbosaR. B. de SousaJ. N. L. Torres R. A. RibeiroM. L. ValeM. H. L. P. Souza
LAFICA Laboratory of Pharmacology of Inflammation and Cancer, Department of Physiology and Pharmacology, Federal University of Ceara´, Fortaleza, CE 60430-270, Brazil
T. M. CunhaF. Q. Cunha
Laboratory of Inflammation and Pain, Department of
Pharmacology, Ribeira˜o Preto Medical School, Av. Bandeirantes 3900, Ribeira˜o Preto, SP 14049-900, Brazil
P. M. G. Soares
Department of Morphology, NEMPI Center for Microscopy and image processing, Federal University of Ceara´, Fortaleza, CE 60430-170, Brazil
luminal application of irritants can cause longer withdrawal latencies and higher thresholds for mechanical stimulation in areas distant from the convergent somatic referral sites [8–11]. However, the mechanisms involved in the hypo-sensitivity response associated with CD have not been completely elucidated.
Several pharmacological approaches have been used to generate antinociception. Cannabinoids and opioids can produce peripheral antinociceptive effect through the stimulation of the L-arginine/NO/cGMP/PKG/KATP
path-way [12]. Cannabinoid agonists produce peripheral antinociception by activation of peripheral cannabinoid receptors, leading to the release of endogenous opioids and subsequent local generation of NO and cGMP [13–15]. However, the involvement of the NO/cGMP/PKG/KATP
pathway, cannabinoid and opioid systems in the remote antinociception associated with TNBS-induced colitis (TNBS colitis) in rats has not been studied.
Methods
Animals
Wistar rats weighing 180–200 g were housed in tempera-ture-controlled rooms and received water and food ad libitum until use. All experiments were conducted in accordance with NIH standards and were approved by the Committee of Ethics (protocol number 10/11) in Animal Research and Care of the Federal University of Ceara´. Drugs
2,4,6-trinitrobenzene sulphonic acid (TNBS), carrageenan, dextran, prostaglandin (PG) E2, L-Noarg, L-Arginine
(L-Arg), 1H- [1, 2, 4] -oxadiazolo [4,3-a] quinoxalin-1-one
(ODQ), glibenclamide, naloxone and KT 5823 were pur-chased from Sigma Chemicals (St. Louis, MO, USA). AM251 and AM630 were purchased from Tocris. PBS buffer or saline was used for vehicle solutions. Glibencla-mide (Glib) was dissolved in saline and 2 % Tween (Sigma, St. Louis, MO, USA). ODQ, KT5823, AM251 and AM630 were dissolved in saline and 4 % dimethyl sulph-oxide (Sigma, St. Louis, MO, USA). PGE2, carrageenan,
L-Noarg, andL-Arginine were dissolved in saline.
Induction of TNBS-induced colitis
TNBS-induced colitis was induced in male Wistar rats (180–220 g) by intracolonic administration of 20 mg of TNBS in 50 % ethanol. Other groups received ethanol
anus and 20 mg of TNBS in 50 % ethanol or ethanol in 50 % of saline was administered. Animals were then kept in a vertical position for 30 s and returned to their cages. Mechanical hypersensitivity
Hypersensitivity was induced by a subcutaneous injection of carrageenan (500 lg/paw) or PGE2 (100 ng/paw) into
the plantar surface of the right hind paw and measured by the paw pressure test [16]. The investigator was trained to apply the polypropylene filaments perpendicular to the central area of the hind paw with a gradual increase in pressure. During testing, pressure was applied to the plantar surface of hind paw to provoke a flexion reflex followed by a clear flinching response after paw withdrawal. Each von Frey filaments was applied for approximately 3–4 s to induce the end-point reflex. The most sensitive filament capable of eliciting a response was determined to be the mechanical threshold (g). A digital analgesiometer (Insight, Brazil) coupled to a rounded tip was used to realise the experiment. The nociceptive threshold was measured in the right paw and determined by the average of three consecutive trials recorded before (zero time) and 3 and 4 h after injection of carrageenan (100 ll; 500lg/ paw) or PGE2 (50ll; 100 ng/paw) (peak effect).
Hyper-algesia was calculated by the difference between these two averages (D of nociceptive threshold) and expressed in grams. To reduce stress, the rats were habituated to the apparatus 1 day prior to the experiments.
Evaluation of the antinociceptive effect of TNBS-induced colitis on carrageenan or PGE2-induced hypersensitivity
Three days after intracolonic administration of 20 mg of TNBS in 50 % ethanol, vehicle or an equivalent volume of saline, carrageenan- or PGE2 -induced mechanical
hyper-sensitivity was evaluated in the right paw. The nociceptive threshold was measured in the right paw and determined by the average of three consecutive trials recorded before (zero time) and 3 and 4 h after injection of carrageenan (100 ll; 500lg/paw) or PGE2(50ll; 100 ng/paw).
Evaluation of theL-Arginine/NO/PKG/KATP? pathway
in the antinociceptive effect of TNBS-induced colitis in carrageenan- or PGE2-induced hypersensitivity
TNBS?ODQ (8lg/paw; 50ll, intraplantar injection), TNBS?KT 5823 (1.5 lg/paw; 50ll, intraplantar injec-tion) and TNBS?glibenclamide (160lg/paw; 50ll, intraplantar injection). Each one of the substances, vehicles or diluents was tested alone. Rats were pre-treated with
L-Noarg one hour before carrageenan injection.L-arginine
was given 10 min before L-Noarg injections. ODQ, KT
5823 and glibenclamide were administered 30 min before carrageenan (100ll; 500lg/paw) or PGE2 (50ll; 100
ng/paw), and the nociceptive threshold was measured as described above.
Role of endogenous opioids and cannabinoids
in the antinociceptive effect induced by TNBS-induced colitis
Three days after intracolonic administration of 20 mg of TNBS in 50 % ethanol, 50 % ethanol or saline only, nal-oxone (nonspecific opioid receptor antagonist; 1lg/paw; 50ll intraplantar injection), saline (diluent for naloxone), AM251 (receptor antagonist cannabinoid type I; 80lg/ paw; 50ll intraplantar injection), AM630 (receptor antagonist cannabinoid type II; 25lg/paw; 50ll intrapl-antar injection) or dimethyl sulphoxide (diluent for AM 251 and AM 630) was injected 30 min prior to adminis-tration of carrageenan (100ll; 500lg/paw) or PGE2
(50ll; 100 ng/paw) in the right paw. In control groups of rats without colitis, naloxone, AM251 or AM630 was also injected before carrageenan or PGE2 administration. The
nociceptive threshold was then measured in the right paw and determined as described above.
Data analysis
The results are presented as the mean±SEM (n=6) for groups of six animals each. Differences between experi-mental groups were compared by analysis of variance (ANOVA) followed by Bonferroni testing. The signifi-cance level was set atP\0.05.
Results
Effect of TNBS-induced colitis on carrageenan-or PGE2-induced hypersensitivity
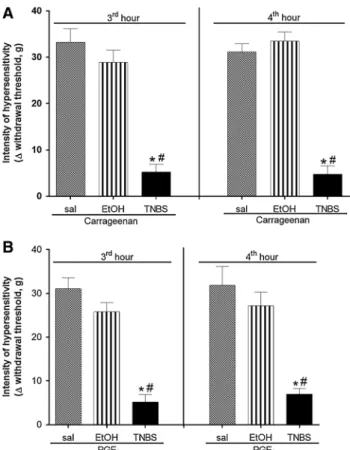
TNBS-induced colitis, but not ethanol alone, resulted in decreased hypersensitivity induced by carrageenan (Fig.1a; 3 h: 5.2±1.68 g/4 h: 4.74±1.83 g) and PGE2 (Fig.1b; 3 h: 5.03±1.59 g/4 h: 7.00 ±1.27 g) when compared to saline plus carrageenan or PGE2(carrageenan;
3 h: 33.24±2.91 g/4 h: 31.12±1.82 g/PGE2; 3 h: 31.08±2.51 g/4th hour: 31.90 ±4.26 g).
Role ofL-arginine/NO/PKG/KATP?
in the antinociceptive effect of TNBS-induced colitis in carrageenan or PGE-induced hypersensitivity
Our results show that animals with colitis induced by TNBS showed a significant anti-nociceptive effect in comparison with the carrageenan or PGE2 groups. This
effect was reversed by treatment withL-Noarg (nitric oxide
synthase nonspecific inhibitor, Figs.2a, 3a), ODQ (gua-nylyl cyclase inhibitor, Figs.2b, 3b), KT 5823 (selective inhibitor of protein kinase G, Figs.2c, 3c) and glibencla-mide (KATP channel blocker, Figs. 2d, 3d). When
administered alone, these drugs did not result in increased inflammatory carrageenan (Fig.2) or PGE2-induced hypersensitivity (Fig.3).
Fig. 1 Effect of TNBS-induced colitis induced by TNBS on
carra-geenan or PGE2-induced hypersensitivity. Carrageenan (500lg/paw)
or PGE2(100 ng/paw) was injected and paw hypersensitivity was
measured 3 and 4 h after carrageenan and PGE2intraplantar injection.
aEvaluation of paw hypersensitivity induced by carrageenan after 3 and 4 h on the third day after colitis induction by TNBS.bEvaluation of paw hypersensitivity induced by PGE23 and 4 h on the third day
after colitis induction by TNBS.*P\0.05 compared to saline group; #P
Fig. 2 Role of theL-Arginine/
NO/PKG/KATP
?
pathway in the antinociceptive effect of TNBS-induced colitis in carrageenan-induced hypersensitivity. Carrageenan (500lg/paw) was administered in the right paw of rats with or without colitis on the third day after colitis induction. The analgesic effect of TNBS-induced colitis was measured on the right hind paw 3 or 4 h after carrageenan-induced hypersensitivity.L-Arg
(200 mg/kg; 0,5 ml;a) was administered 10 min before
L-Noarg injection (a). ODQ
(8lg/paw; 50ll;b), KT 5823 (1,5lg/paw; 50ll, intraplantar
c) and glibenclamide (160lg/ paw; 50ll, intraplantar injection;d) were administered 30 m before carrageenan administration in the right paw. Asteriskstatistically significant differences between the TNBS group and saline group.Hash statistically significant differences between the colitis by TNBS group and ethanol (EtOH) group.Psistatistically significant differences between the groups treated withL-Noarg,
ODQ, KT 5823, glibenclamide and the TNBS group.Upsilon statistically significant differences between theL-Arg group and the
TNBS?L-Arg?L-Noarg group.P\0.05 for
Fig. 3 Role ofL-Arginine/NO/
PKG/KATP
?
pathway in the antinociceptive effect of TNBS-induced colitis in PGE2-induced
hypersensitivity. PGE2(100 ng/
paw; 50ll) was administered in the right paw of rats with or without colitis on the third day after colitis induction. The analgesic effect of TNBS-induced colitis was measured on the right hind paw 3 or 4 h after PGE2-induced hypersensitivity. L-Arg (200 mg/kg; 0,5 ml;b)
was administered 10 min before
L-Noarg injection (a). ODQ
(8lg/paw; 50ll;b), KT 5823 (1,5lg/paw; 50ll;c) and glibenclamide (160lg/paw; 50ll;d) were administered 30 m before PGE2
administration in the right paw of rats.Asteriskstatistically significant differences between the colitis by TNBS group and saline group.Hashstatistically significant differences between the colitis by TNBS group and ethanol (EtOH) group.Psi statistically significant differences between the groups treated withL-Noarg, ODQ, KT
5823, glibenclamide and the TNBS group.Upsilon statistically significant differences betweenL-Arg
group and TNBS?L-Arg? L-Noarg group.P\0.05 to test
Role of endogenous opioids and cannabinoids in the antinociceptive effect of TNBS-induced colitis induced by TNBS
As observed in Fig.4, naloxone significantly reversed the TNBS-induced antinociceptive effect following adminis-tration of carrageenan (Fig.4, a) or PGE2 (Fig.4, b).
Similarly, AM251 or AM630 also significantly reversed (Fig.5, a and b) the antinociceptive effect observed in animals with TNBS-induced colitis after carrageenan (a) or PGE2( b) administration.
Discussion
Crohn’s disease is a chronic inflammatory disorder pri-marily affecting the gastrointestinal tract. This intestinal inflammation is accompanied by extraintestinal manifes-tations. The most common neurological dysfunction found in IBD is peripheral neuronal desensitisation. Peripheral desensitisation (PD) is one of the most frequently reported
symptoms are commonly identified in patients with CD [1, 17] and also have been verified in animal models of intestinal inflammation [7]. However, the mechanisms involved in this neurological disorder are not totally understood.
Our results demonstrated that colitis induced by TNBS produced a peripheral antinociceptive effect in PGE2 or
carrageenan-induced mechanical hypersensitivity, and this effect was at least in part mediated by the opioid and cannabinoid systems, and further by activation of the NO/cGMP/PKG pathway followed by opening of KATP
?
channels.
The induction of inflammatory hypersensitivity begins through the sensitisation of primary nociceptive neurons, an effect that is caused by nociceptor-sensitising mediators such as IL-1b, TNF-a, IL-6, and the chemokines IL-8, and chemokine-induced neutrophil chemoattractant-1 (CINC-1) [18]. Carrageenan-induced hypersensitivity is dependent on the concomitant release and action of IL-1b, TNF-aand IL-8, which induces the subsequent release of PGE2and PAF by
resident cells [19]. Alternately, PGE2 acts on nociceptive
Fig. 4 Role of endogenous
opioids in the antinociceptive effect of TNBS-induced colitis. Naloxone (nonspecific opioid receptor antagonist; 1lg/paw; 50ll intraplantar injection) was administered 30 m before carrageenan (500lg/paw; 100ll;a) or PGE2(100 ng/
paw; 50ll,b) administration in the right paw of rats with or without colitis on the third day after colitis induction. The analgesic effect of TNBS-induced colitis was measured on the right hindpaw 3 or 4 h after carrageenan or PGE2-induced
hypersensitivity.Asterisk statistically significant differences between the colitis by TNBS group and saline group.Hashstatistically significant differences between the colitis by TNBS group and ethanol (EtOH) group.Psi indicates statistically significant differences between colitis by TNBS?naloxone group and the colitis by TNBS group. P\0.05 for significance
receptors, increasing the production of cAMP and thus generating an increased calcium influx [19,20]. According to our results, we can infer that the increase in desensitisation in the group of animals with TNBS-induced colitis seems to be partially dependent on the action of cytokines on noci-ceptive neurons. However, it is not the only mechanism involved in this desensitisation, once the effect PGE2 in
peripheral sensitisation is independent of the action of pro-inflammatory cytokines on peripheric nociceptors, and our results demonstrated that this PGE2nociceptive effect was
inhibited during the TNBS-induced colitis.
In active CD, the colonic mucosa produces and releases NO at a high concentration. Interestingly, NO-positive neurons are increased in submucosal and myenteric plex-uses of the small intestine in CD [21–23]. A study using tonometric sampling of gas in the rectum detected a marked increase in NO rectal concentration in patients with CD [24]. Additionally, there is evidence that an overpro-duction of NO and its metabolites nitrite and nitrate occurs in intestinal inflammation of patients with CD. These same
substances were also found in high concentrations in the plasma of these patients [25–29].
NO is involved in several mechanisms of antihyper-sensitivity, but the involvement of nitric oxide in PD during TNBS-induced colitis has not been clearly demon-strated. This endogenous gas participates in the analgesic effects of dipyrone, opioids and pertussis toxin [15,30,31]. In inflammatory hypersensitivity, NO activates the enzyme guanylate cyclase, which is directly responsible for increases in intracellular levels of cGMP [32]. Then, cGMP induces the opposite effect of cAMP and promotes anti-hypersensitivity [33]. The literature data demonstrated that activation of protein kinase G (PKG) by cGMP is necessary for the opening of ATP-sensitive potassium channel (KATP)
channels in analgesia [31]. However, there is no evidence for the participation of this analgesic biochemical pathway in the neuronal desensitisation associated with colitis.
Using inhibitors of theL-arginine/NO/cGMP/PKG/KATP
pathway, we demonstrated that the antinociceptive effect of TNBS-induced colitis involves this biochemical
Fig. 5 Role of endogenous cannabinoids in antinociceptive effect of
TNBS-induced colitis. AM251 (receptor antagonist cannabinoid type I; 80lg/paw; 50ll intraplantar injection) or AM630 (receptor antagonist cannabinoid type II; 25lg/paw; 50ll intraplantar injec-tion) was administered 30 m before carrageenan (500lg/paw; 100ll; a) or PGE2 (100 ng/paw, 50ll; b) administration to the
right paw of rats with or without colitis on the third day after colitis induction. The analgesic effect of TNBS-induced colitis induced by
TNBS of the right hindpaw was measured 3 or 4 h after carrageenan or PGE2-induced hypersensitivity. Asterisk statistically significant
differences between the colitis by TNBS group and saline group. Hashstatistically significant differences between the colitis by TNBS group and ethanol (EtOH) group.Psistatistically significant differ-ences between the TNBS?AM251 group or TNBS?AM630 group and the colitis by TNBS group.P\0.05 for significance (ANOVA/
pathway. The antinociceptive effect of the experimental model of colitis was blocked by the local administration of the NO synthesis inhibitor L-Noarg. Moreover, it was
demonstrated that the cGMP pathway is associated with colitis induced by the antinociceptive effect of TNBS, as ODQ, a soluble guanylyl-cyclase inhibitor, blocked colitis-induced antinociception, as observed with analgesic drugs such as dipyrone and diclofenac [34, 35]. Therefore, our data suggest that the antinociceptive effect of Crohn’s model is mediated by induction of NO production with subsequent activation of guanylyl cyclase. Pre-treatment with sulphonylureas glibenclamide (ATP-sensitive K? channel blockers) inhibited the antinociceptive effect, suggesting a role of ATP-sensitive K?channels in Crohn’s model-induced antinociception. Similar to analgesic drugs [31], the activation of the ATP-sensitive K?channels by CD is dependent on activation of guanylyl cyclase and PKG, as a specific protein kinase G inhibitor (KT5823) could inhibit the colitis-induced antinociceptive effect. Our data are in accordance with studies demonstrating that PKG promotes the opening of ATP-sensitive K?channels in other systems [36–38]. According to our results, we can infer that, most likely, Crohn’s disease can stimulate the production of nitric oxide in inflamed colon tissue and NO activated the enzyme guanylate cyclase, producing PD by activation of the PKG/KATP pathway.
Other systems involved in the peripheric neuronal desensitisation are the opioids and cannabinoids. The activation of both systems causes a decrease in cAMP production by inhibiting the enzyme adenylate cyclase, producing a blocker of calcium channels that are voltage sensitive to the opening of potassium channels [39–41]. Moreover, opioids and cannabinoids can produce an anti-hyperalgesic effect by activating the NO/cGMP/PKG/KATP
pathway [13–15]. Our results clearly demonstrated that the pre-treatment with nonspecific receptor opioid blocker (naloxone) or specific receptor cannabinoid type I (AM251) and type II (AM630) antagonists reversed the antinociceptive effect induced by TNBS-induced colitis, suggesting that the systems play a role in this response.
Endocannabinoids and endogenous opioids are involved in protective mechanisms, reduce mucosal damage and improve bowel motility [42]. The same study showed that during colonic inflammation, high levels of anandamide and endogenous opioids have been found in biopsy samples of patients with colitis as well as in the colon of TNBS-colitis in rats [42]. However, no data show the participation of cannabinoid and opioid systems in PD during TNBS-induced colitis.
According to our findings, we can infer that the endo-cannabinoids released from the inflamed colon can activate
generation of NO and activation of the NO/cGMP/PKG/ KATP
?
pathway [13–15] and promoting peripheral neuronal desensitisation.
Conclusions
In conclusion, the present study showed an intrinsic peripheral neuronal desensitisation during colonic inflammation induced by TNBS. This neuronal disorder seemed to be mediated by activation of the NO/cGMP/ PKG/KATP
?
pathway, endocannabinoids and endogenous opioids. This information may contribute to a better comprehension of the peripheral neurological disorders in patients with CD.
Acknowledgments The authors gratefully acknowledge the
tech-nical assistance of Maria Silvandira Freire Franc¸a, and we thank UFPI/CNPq for fellowship support.
References
1. Gondim FAA, Brannagan TH, Sander HW, Chin RL. Peripheral neuropathy in patients with inflammatory bowel disease. Brain. 2005;128:867–79.
2. Greenstein AJ, Janowitz HD, Sachar DB. The extra-intestinal complications of Crohn’s disease and ulcerative colitis: a study of 700 patients. Medicine. 1976;55:401–12.
3. Gendelman S, Present D, Janowitz HD. Neurological complica-tions of inflammatory bowel disease. Gastroenterology. 1982;82:1065–75.
4. Rankin GB, Watts HD, Melnyk CS, Kelley ML Jr. National cooperative Crohn’s disease study: extraintestinal manifestations and perianal complications. Gastroenterology. 1979;77:914–20. 5. Lossos A, River Y, Eliakim A, Steiner I. Neurologic aspects of
inflammatory bowel disease. Neurology. 1995;45:416–21. 6. Elsehety A, Bertorini TE. Neurologic and neuropsychiatric
complications of Crohn’s disease. S Med J. 1997;90:606–10. 7. Richard JT, Wang G. Colonic inflammation decreases thermal
sensitivity of the forepaw and hindpaw in the rat. Neurosci Lett. 2004;359:81–4.
8. Chandler MJ, Qin C, Zhang J, Foreman RD. Differential effects of urinary bladder distension on high cervical projection neurons in primates. Brain Res. 2002;949:97–104.
9. Ness TJ, Gebhart GF. Interactions between visceral and cutane-ous nociception in the rat. Noxicutane-ous cutanecutane-ous stimuli inhibit visceral nociceptive neurons and reflexes. J Neurophysiol. 1991;66:20–8.
10. Qin C, Chandler MJ, Foreman RD. Effects of urinary bladder distension on activity of T3–T4 spinal neurons receiving cardiac and somatic noxious inputs in rats. Brain Res. 2003;971:210–20. 11. Traub RJ, Wang G. Colonic inflammation decreases thermal sensitivity of the forepaw and hindpaw in the rat. Neurosci Lett. 2004;359:81–4.
12. Wise LE, Cannavacciulo R, Cravatt BF. Evaluation of fatty acid amides in the carrageenan-induced paw edema model. Neuro-pharmacology. 2008;54:181–8.
14. Reis GML, Ramos MA, Pacheco DF. Endogenous cannabinoid receptor agonist anandamide induces peripheral antinociception by activation of ATP-sensitive K? channels. Life Sci. 2011;88:653–7.
15. Cunha TM, Roman-Campos D, Lotufo CM, et al. Morphine peripheral analgesia depends on activation of the PI3Kgamma/ AKT/nNOS/NO/KATP signaling pathway. Proc Natl Acad Sci. 2010;107:4442–7.
16. Cunha TM, Verri WA, Vivancos GG. An electronic pressure-meter nociception paw test for mice. Braz J Med Biol Res. 2004;37:401–7.
17. Stahlberg D, Barany F, Einarsson K. Neurophysiologic studies of patients with Crohn’s disease on long-term treatment with met-ronidazole. Scand J Gastroenterol. 1991;26:219–24.
18. Cunha TM, Verri WAJr, Parada CA, et al. Hypernociceptive role of cytokines and chemokines: targets for analgesic drug devel-opment? Pharmacol Ther. 2006;112:116–38.
19. DiRosa M, Giroud JP, Willoughby DA. Studies on acute inflammatory response mediators induced in rats in different sites by carrageenan and turpentine. J Pathol. 1971;104:15–29. 20. Soares AC, Leite R, Tatsuo MAKF, et al. Activation of
ATP-sensitive K?channels : mechanism of peripheral antinociceptive action of the nitric oxide donor, sodium nitroprusside. Eur J Pharmacol. 2000;400:67–71.
21. Rachmilewitz D, Stamler JS, Bachwich D, et al. Enhanced colonic nitric oxide generation and nitric oxide synthase activity in ulcerative colitis and Crohn’s disease. Gut. 1995;36:718–23. 22. Roediger WEW, Lawson MJ, Nance SH, et al. Detectable colonic
nitrite levels in inflammatory bowel disease—mucosal or bacte-rial malfunction? Digestion. 1986;35:199–204.
23. Oudkerk-Pool M, Bouma G, Visser JJ. Serum nitrate levels in ulcerative colitis and Crohn’s disease. Scand J Gastroenterol. 1995;30:784–8.
24. Geboes K, Mebis J, Rutgeers P, et al. Demonstration of nitric oxide positive neurons in Crohn’s disease. Gastroenterology. 1993;104:705–10.
25. Guslandi M. Nitric oxide and inflammatory bowel diseases. Eur J Clin Invest. 1998;28:904–7.
26. Levine JJ, Pettei MJ, Valderrama E. Nitric oxide sand inflam-matory bowel disease: evidence for local intestinal production in children with active colonic disease. J Ped Gastroenterol Nutr. 1998;26:34–8.
27. Boughton-Smith NK, Evans SM, Hawkey CJ. Nitric oxide syn-thase activity in ulcerative colitis and Crohn’s disease. Lancet. 1993;342:338–40.
28. Singer II, Kawka DW, Scott S. Expression of inducible nitric oxide synthase and nitrotyrosine in colonic epithelium in inflam-matory bowel disease. Gastroenterology. 1996;111:871–85. 29. Miller MJS, Xhang XJ, Sadowska-Krowicka H. Nitric oxide
release in response to gut injury. Scand J Gastroenterol. 1993;28:149–54.
30. Brito GAC, Sachs D, Cunha FQ, et al. Peripheral antinociceptive effect of pertussis toxin: activation of the arginine/NO/cGMP/ PKG/TP-sensitive K?
channel pathway. Eur J Neurosci. 2006;24:1175–81.
31. Sachs D, Cunha FQ, Ferreira SH. Peripheral analgesic blockade of hypersensitivity: activation of arginine/NO/cGMP/protein kinase G/ATP-sensitive K?
channel pathway. PNAS. 2004;101:3680–5. 32. Moncada S, Palmer RM, Higgs EA. Nitric oxide: physiology,
pathophysiology, and pharmacology. Pharmacol Rev. 1991;43:109–42.
33. Goldberg ND, Haddox MK, Nicol SE. Biologic regulation through opposing influences of cyclic GMP and cyclic AMP: the Yin Yang hypothesis. Adv Cyclic Nucleotide Res. 1975;5:307–30. 34. Granados-Soto V, Flores-Murrieta FJ, Castaneda-Hernandez G,
et al. Evidence for the involvement of nitric oxide in the antin-ociceptive effect of ketorolac. Eur J Pharmacol. 1995;277:281–4. 35. Lorenzetti BB, Ferreira SH. Activation of the arginine-nitric oxide pathway in primary sensory neurons contributes to dipy-rone-induced spinal and peripheral analgesia. Inflamm Res. 1996;45:308–11.
36. Han J, Kim N, Kim E, et al. Modulation of ATP-sensitive K?
channels by cGMP-dependent protein kinase in rabbit ventricular myocytes. J Biol Chem. 2001;276:22140–7.
37. Han J, Kim N, Joo H, et al. ATP-sensitive K?
channel activation by nitric oxide and protein kinase G in rabbit ventricular myo-cytes. Am J Physiol Heart Circ Physiol. 2002;283:1545–54. 38. Segawa K, Minami K, Shiga Y, et al. Inhibitory effects of
nic-orandil on rat mesangial cell proliferation via the protein kinase G pathway. Nephron. 2001;87:263–8.
39. Cichewicz DL. Synergistic interactions between cannabinoid and opioid analgesics. Life Sci. 2004;74:1317–24.
40. Howlett AC, Barth F, Bonner TI. Classification of cannabinoid receptors. Pharmacol Rev. 2002;54:161–202.
41. Vigano D, Rubino T, Parolaro D. Molecular and cellular basis of cannabinoid and opioid interactions. Pharmacol Biochem Behav. 2005;81:360–8.