Cover crop rotations in no-till system: short-term CO
Texto
Imagem




Documentos relacionados
The aim of the study is to explore social and cultural factors that influence the practice of children’s participation rights in Vietnamese families in Vietnam
Esse documento foi redigido, em dezembro de 1844, por Guilherme Martins do Nascimento, um marceneiro crioulo, provavelmente sobrinho de Antonio Caetano Coelho, quando Joaquim
The results obtained from the second numerical trial, which considered a total of ten periods, four plots and three animals (b1, b2, b3) also show the validity of the model
Após banho e secagem, aplicar em camada Þ na, uniforme e com massagem ligeira em toda a pele abaixo do queixo e, em particular, nas pregas; fazer uma segunda aplicação 24 horas
Four areas submitted to distinct managements (conventional tillage system; no-till system; integration crop/livestock and continuous livestock) were sampled in relation to
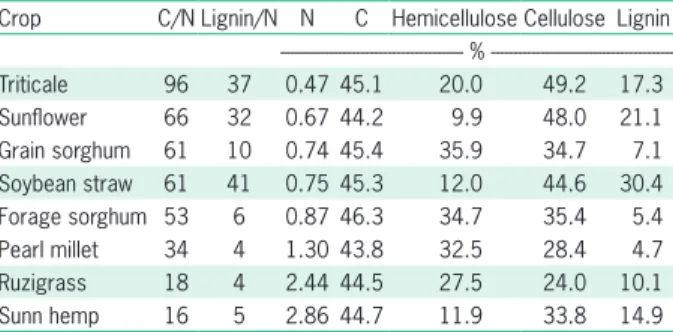
occurs in the crop itself, reducing the crops response to N fertilization. The use of agricultural land, with a intercropping cover system in winter, results in greater
(1998), at a place nearby and in the same period of the previous study, with crop rotation systems for wheat (system I: wheat/soybean; system II: wheat/soybean, and
In order to achieve sustainability, different crop combinations and different strategies for management of the spontaneous vegetation will be tested, and the crop systems will
