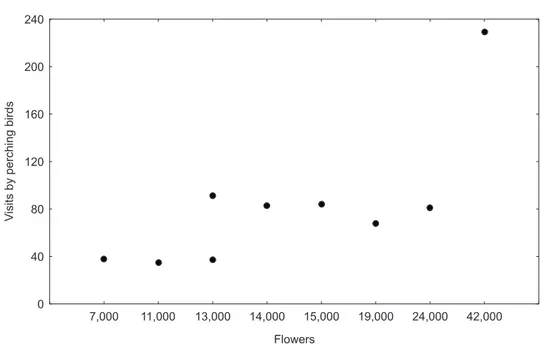
EXPLOITATION OF Erythrina dominguezii Hassl. (Fabaceae) NECTAR BY PERCHING BIRDS IN A DRY FOREST IN WESTERN BRAZIL
Texto
Imagem
Documentos relacionados
Dentre essas variáveis destaca-se o “Arcabouço Jurídico-Adminis- trativo da Gestão Pública” que pode passar a exercer um nível de influência relevante em função de definir
Diante dessas questões, senti a necessidade de continuar trilhando esse caminho de (auto) formação e desenvolvimento profissional, com intuito de conhecer e compreender,
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
In anemochorich and autochorich species, leaf loss started in the early dry season, mainly in those species which flowered in the middle of this period while leaf- less
Os modelos desenvolvidos por Kable & Jeffcry (19RO), Skilakakis (1981) c Milgroom & Fry (19RR), ('onfirmam o resultado obtido, visto que, quanto maior a cfiráda do
A Schinopsis brasiliensis Engl., conhecida como “baraúna” ou “braúna”, apresenta atividade contra diversos patógenos e visando desenvolver um novo medicamento o estudo
This suggests that the recent drought may have in fact started in the middle-late 1990s, with the intense droughts of 1993 and 1998, and then the sequence of dry years (interrupted
This suggests that the recent drought may have in fact started in the middle-late 1990s, with the intense droughts of 1993 and 1998, and then the sequence of dry years (interrupted
