Brain and Peripheral Organ Toxicity of "Ecstasy" in Adolescent Rats in Human Relevant Doses
Texto
Imagem
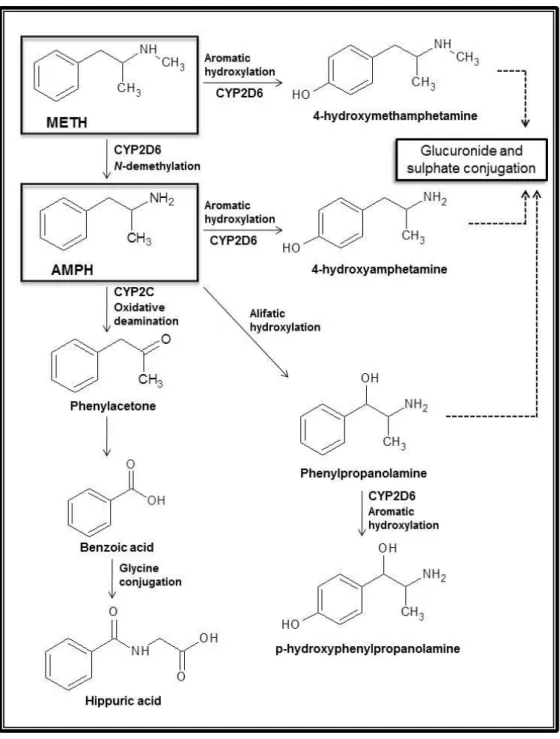
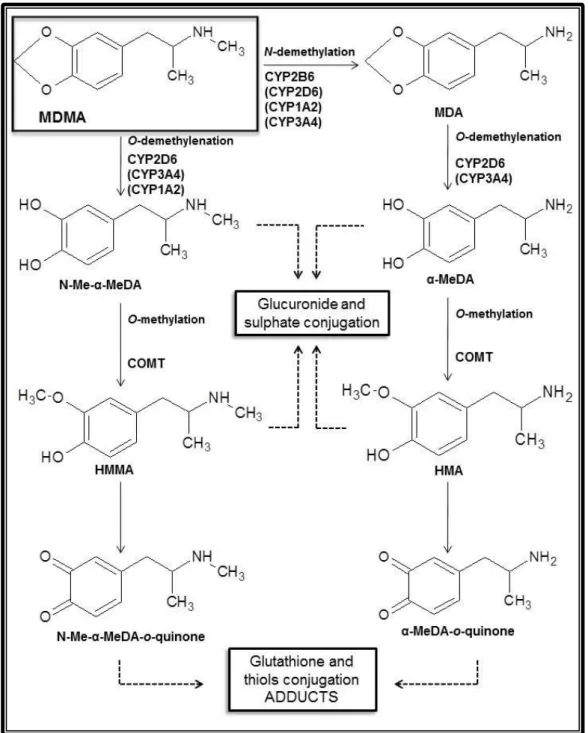
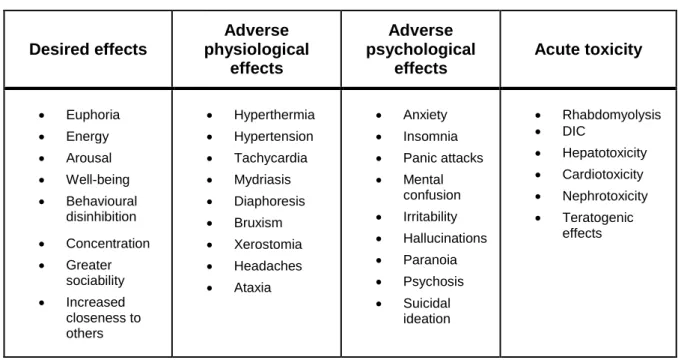
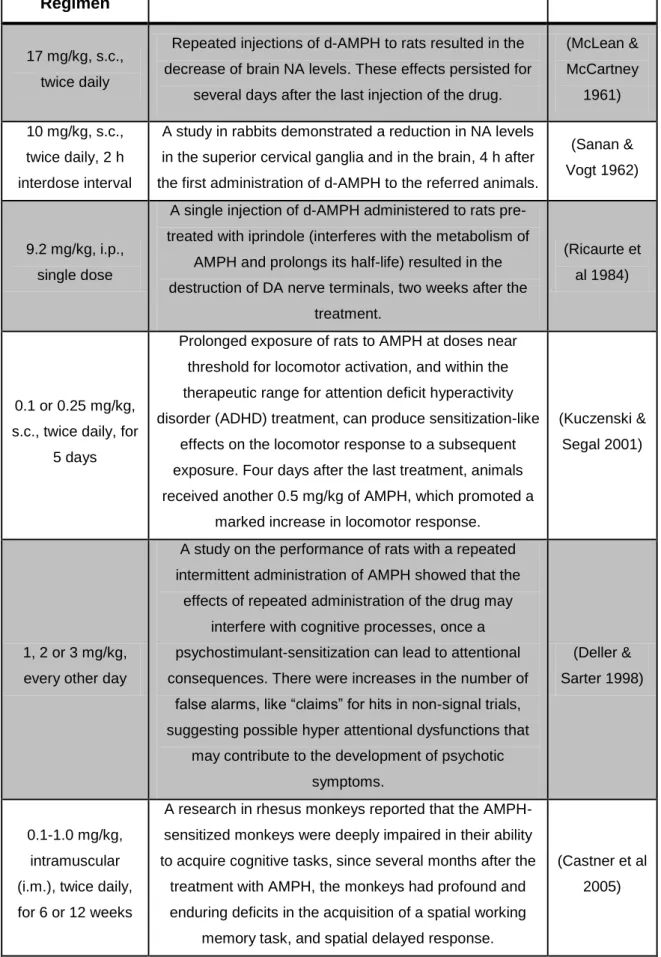
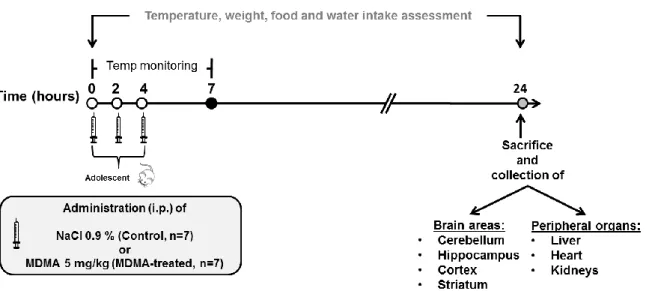
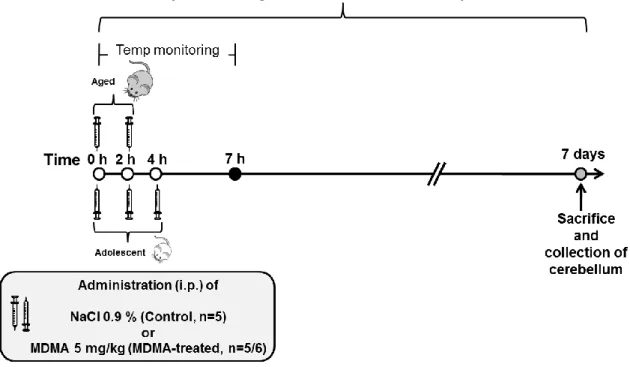
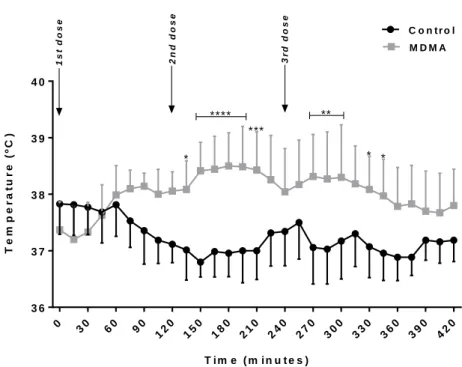
Documentos relacionados
Este artigo discute o filme Voar é com os pássaros (1971) do diretor norte-americano Robert Altman fazendo uma reflexão sobre as confluências entre as inovações da geração de
Nota: Envolve dados dos seguintes cursos cadastrados como licenciaturas e com os seguintes nomes: Ciência da Computação, Ciências da Computação, Computação, Computação
To evaluate if the ALDH3A2, ALDH3B1, and ALDH4A1 isoforms could to be used as MDS diagnostic markers, we determine the capacity of the gene expression levels of these ALDHs
However, we found that across the monetary and the physical condition subjects showed the same level of trust and reciprocity. Moreover, subjects’ behavior across
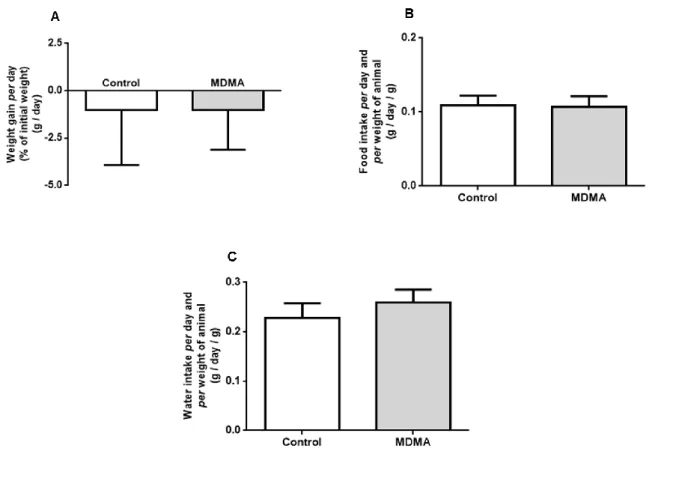
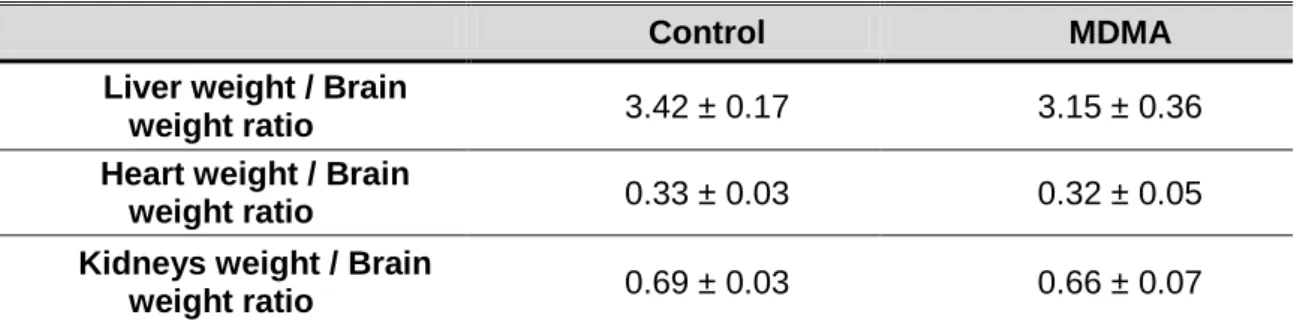
This extract showed no acute or chronic toxicity in rats, as confirmed by no change in the body weight of the treated animals compared to controls and no significant changes in.
Root length (A), root volume (B) and root fresh mass (C) of soybean plants, depending on the application form and concentration of manipueira, in the management of
To determine the relative expression levels of AQP4 throughout the brain, protein was isolated from the cortex, diencephalon, hippocampus, cerebellum, and brainstem, separated
Univariate analysis of size-adjusted shell measurements revealed significant di fferences between the two taxa for shell depth, muscle scar height, and length of ligamental area..
