Influence of local and landscape characteristics on the distribution and diversity of fish assemblages of streams in the Ivinhema River basin, Upper Paraná River
Texto
Imagem


Documentos relacionados
Ao constatar isso, o autor italiano entende que, por mais que essas noções possam ser mecânicas, também são instrutivas, e que a instrução (ensino) também é uma forma de
Desde então os quadrinhos passam de mídia desacreditada, já que durante muito tempo foram vistos apenas como produto cultural destinado a um publico infanto-juvenil, à forma de
“Nosso carro forte aqui é a pokã e a mandioca, quando nós temos muito produto nós leva, as vezes eles levam pra lá, e nós pegamos um pouco os produtos de lá, e nós fazemos
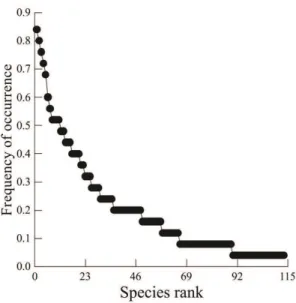
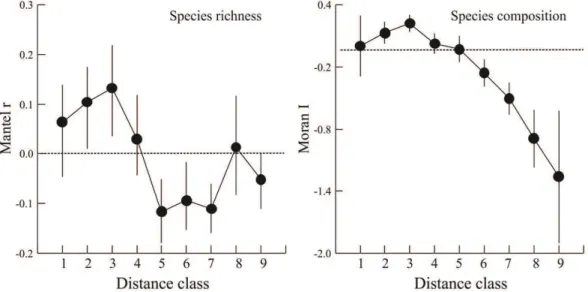
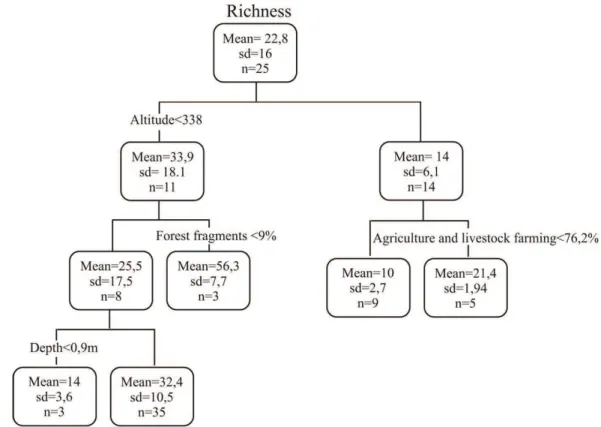
Thus, this study aimed to verify whether the mountain streams of the Tibagi river basin present spatial and temporal patterns in their structure and composition of the fish
A média mais baixa encontrada na escala foi em relação ao item Procuro formas (M= 3,47 e DP= 1,35), o que demonstra que as pessoas residentes no Sul do
Nessa perspectiva, uma das formas de abordagem da Acessibilidade informacional é, portanto, o estudo dos fatores e das melhores práticas que a favorecem. Do ponto de vista do núcleo
FIGURA 5 - FLUTUAÇÃO POPULACIONAL DE Hypothenemus eruditus, Sampsonius dampfi e Xyleborus affinis (SCOLYTINAE, CURCULIONIDAE) COLETADOS COM ARMADILHAS ETANÓLICAS (25%)
Polar and dispersive components of the total surface energy of paperboard before (uncoated) and after coating with several silica based formulations, without (I to IV) and with (Ia
