Novel Function of Lysine Methyltransferase G9a in the Regulation of Sox2 Protein Stability.
Texto
Imagem
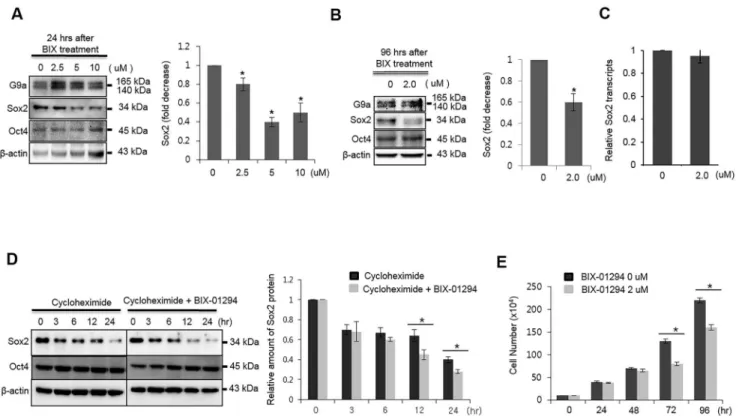
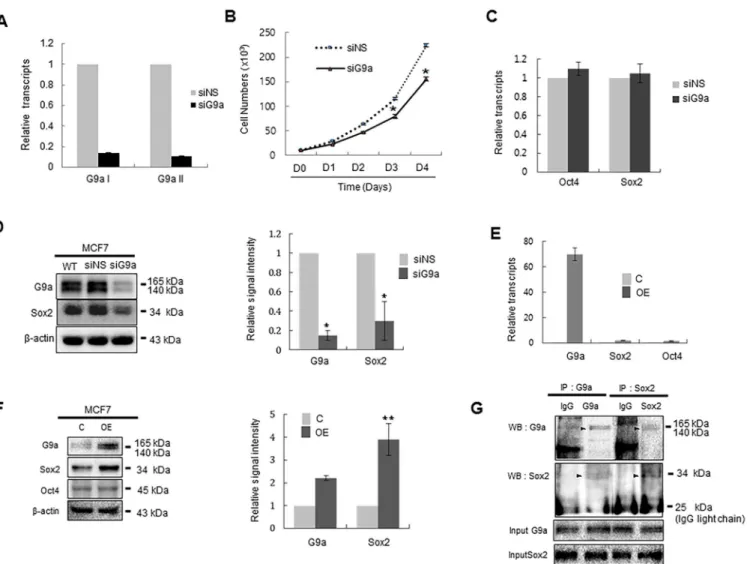
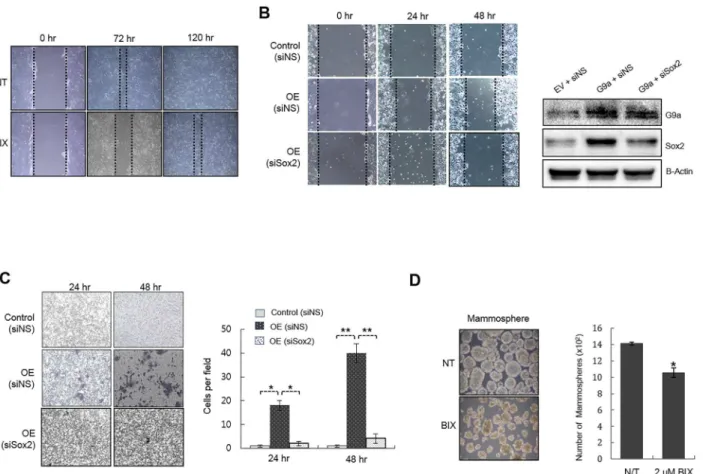
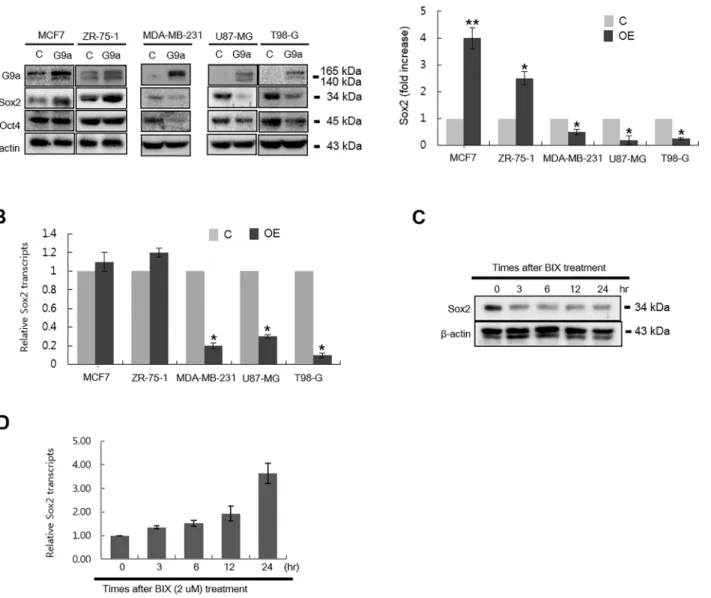
Documentos relacionados
mRNA and protein levels remain constant throughout the differentiation program in MC3T3-E1 cells, we found that in this human osteoblast cell line, the expression of TG2 protein
However, as function of Sox15 is related to Sox2 in some regulation processes in the ES cells (mentioned in the introduc- tion) and Sox15 is closely related to SoxB genes (revealed
Depletion of USP19 results in reduced rates of cell growth and accumulation of p27 Kip1 in MCF10A breast epithelial cells but not in MCF7 and MDA-MB-231 breast carcinoma cells.. We
The canonical signaling protein in autophagy regulation is the mammalian target of rapamycin (mTOR), a ubiquitous protein kinase that is also involved in the regulation of cell
To identify proteins that associate with SOX2 in i-SOX2- DAOY cells, SOX2-protein complexes from uninduced and induced i-SOX2-DAOY cells were subjected to MudPIT analysis. To
social assistance. The protection of jobs within some enterprises, cooperatives, forms of economical associations, constitute an efficient social policy, totally different from
Abstract: As in ancient architecture of Greece and Rome there was an interconnection between picturesque and monumental forms of arts, in antique period in the architecture
We also determined the critical strain rate (CSR), understood as the tangent of the inclination angle between the tangent to the crack development curve and the crack development
